
Analysis of Genetic Diversity in Romanian Carpatina Goats Using SNP Genotyping Data
 , , , , , , and
, , , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Goat Sample Collection and Experimental Design
2.2. Genotyping and Quality Control
2.3. Genetic Diversity
2.4. Linkage Disequilibrium (LD) and Effective Population Size (Ne)
3. Results
3.1. Genetic Diversity
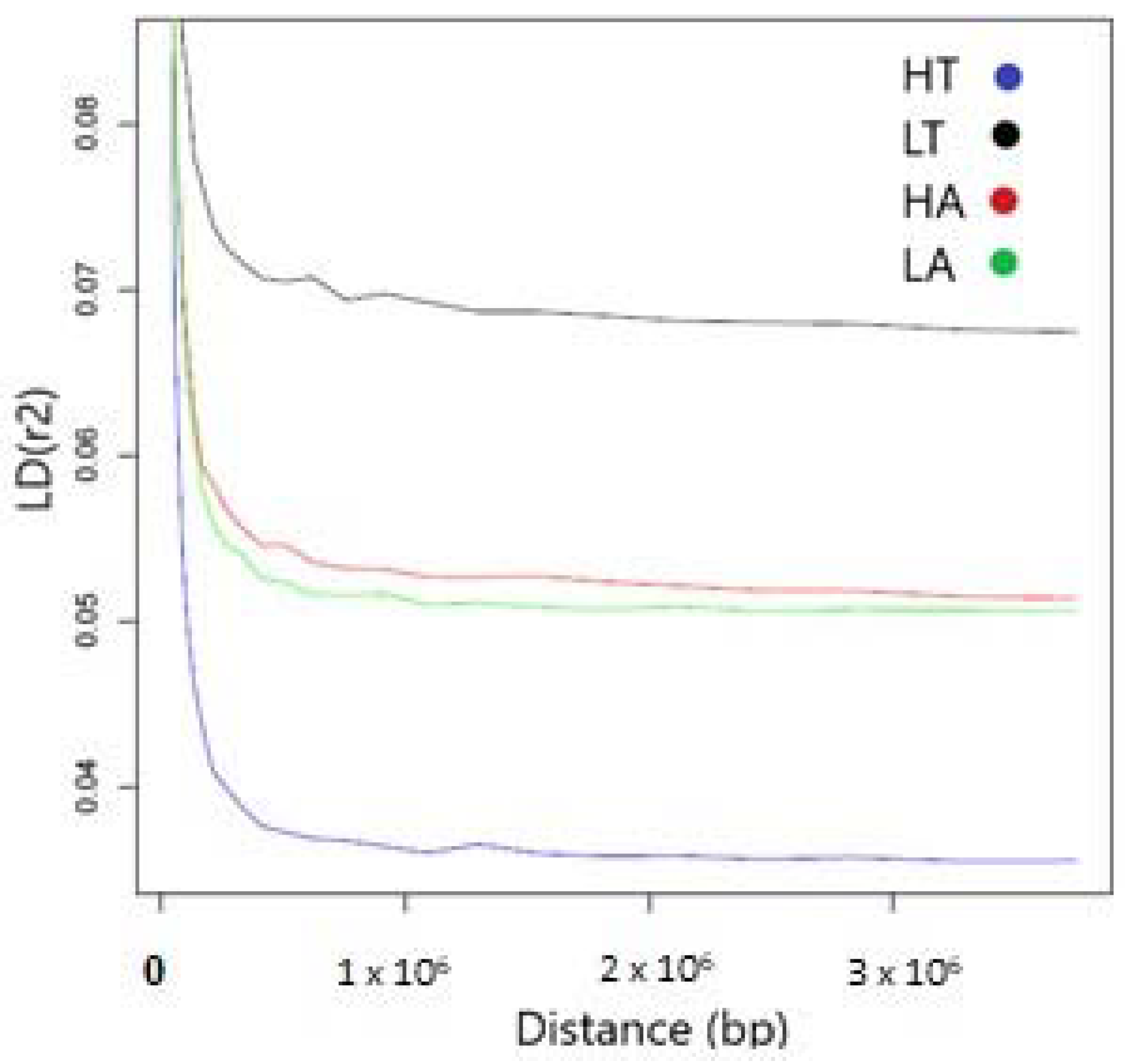
3.2. Linkage Disequilibrium (LD) and Effective Population Size (Ne)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alberto, F.J.; Boyer, F.; Orozco-terWengel, P.; Streeter, I.; Servin, B.; de Villemereuil, P.; Benjelloun, B.; Librado, P.; Biscarini, F.; Colli, L.; et al. Convergent Genomic Signatures of Domestication in Sheep and Goats. Nat. Commun. 2018, 9, 813. [Google Scholar] [CrossRef]
- Mason, I.L. A World Dictionary of Livestock Breeds, Types and Varieties, 4th ed.; CAB International: London, UK, 1996; p. 273. [Google Scholar]
- Vlaic, A.; Odagiu, A.; Vlaic, B.; Dărăban, S.; Coşier, V. Biometric Approach of Carpatina Goat Reared in South Romania Climate. ProEnvironment 2016, 9, 192–196. [Google Scholar]
- Ilie, D.E.; Kusza, S.; Sauer, M.; Gavojdian, D. Genetic Characterization of Indigenous Goat Breeds in Romania and Hungary with a Special Focus on Genetic Resistance to Mastitis and Gastrointestinal Parasitism Based on 40 Snps. PLoS ONE 2018, 13, e0197051. [Google Scholar] [CrossRef]
- Padeanu, I. Tehnologia Cresterii Ovinelor Și Caprinelor; Editura Mirton: Timisoara, Romania, 2001; p. 375. (In Romanian) [Google Scholar]
- Pascal, C.; Padeanu, I.; Calin, I.; Daraban, S.; Nacu, G. Researches related to meat yield aptitudes of Carpatina breed reared in Romania. Sci. Pap. Anim. Sci. Biotech. 2011, 55, 328–331. [Google Scholar]
- Kusza, S.; Ilie, D.E.; Sauer, M.; Nagy, K.; Patras, I.; Gavojdian, D. Genetic Polymorphism of CSN2 Gene in Banat White and Carpatina Goats. Acta Biochimica Polonica 2016, 63, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Zaulet, M.; Kevorkian, S.; Enache, M.; Ghita, E.; Lazar, C.; Rebedea, M. Genetic Polymorphism of some Proteins in the Milk of Carpathian Goat. Sci. Pap. Anim. Husb. Biotechnol. Timişoara 2008, 41, 201–207. [Google Scholar]
- Colli, L.; Milanesi, M.; Talenti, A.; Bertolini, F.; Chen, M.; Crisà, A.; Daly, K.G.; Del Corvo, M.; Guldbrandtsen, B.; Lenstra, J.A.; et al. Genome-Wide SNP Profiling of Worldwide Goat Populations Reveals Strong Partitioning of Diversity and Highlights Post-Domestication Migration Routes. Genet. Sel. Evol. 2018, 50, 58. [Google Scholar] [CrossRef] [PubMed]
- Ajmone-Marsan, P.; Colli, L.; Han, J.L.; Achilli, A.; Lancioni, H.; Joost, S.; Crepaldi, P.; Pilla, F.; Stella, A.; Taberlet, P.; et al. The Characterization of Goat Genetic Diversity: Towards a Genomic Approach. Small Rumin. Res. 2014, 121, 58–72. [Google Scholar] [CrossRef]
- Dixit, S.P.; Verma, N.K.; Aggarwal, R.A.K.; Vyas, M.K.; Rana, J.; Sharma, A.; Tyagi, P.; Arya, P.; Ulmek, B.R. Genetic Diversity and Relationship among Southern Indian Goat Breeds Based on Microsatellite Markers. Small Rumin. Res. 2010, 91, 153–159. [Google Scholar] [CrossRef]
- Korkmaz Ağaoğlu, Ö.; Ertuğrul, O. Assessment of Genetic Diversity, Genetic Relationship and Bottleneck Using Microsatellites in Some Native Turkish Goat Breeds. Small Rumin. Res. 2012, 105, 53–60. [Google Scholar] [CrossRef]
- Nicoloso, L.; Bomba, L.; Colli, L.; Negrini, R.; Milanesi, M.; Mazza, R.; Sechi, T.; Frattini, S.; Talenti, A.; Coizet, B.; et al. Genetic Diversity of Italian Goat Breeds Assessed with a Medium-Density SNP Chip. Genet. Sel. Evol. 2015, 47, 62. [Google Scholar] [CrossRef]
- Visser, C.; Lashmar, S.F.; Van Marle-Köster, E.; Poli, M.A.; Allain, D. Genetic Diversity and Population Structure in South African, French and Argentinian Angora Goats from Genome-Wide SNP Data. PLoS ONE 2016, 11, e0154353. [Google Scholar] [CrossRef]
- Liu, S.; He, S.; Chen, L.; Li, W.; Di, J.; Liu, M. Estimates of Linkage Disequilibrium and Effective Population Sizes in Chinese Merino (Xinjiang Type) Sheep by Genome-Wide Snps. Genes Genom. 2017, 39, 733–745. [Google Scholar] [CrossRef]
- Manunza, A.; Noce, A.; Serradilla, J.M.; Goyache, F.; Martínez, A.; Capote, J.; Delgado, J.V.; Jordana, J.; Muñoz, E.; Molina, A.; et al. A Genome-Wide Perspective about the Diversity and Demographic History of Seven Spanish Goat Breeds. Genet. Sel. Evol. 2016, 48, 52. [Google Scholar] [CrossRef] [PubMed]
- Kosgey, I.S. Breeding objectives and breeding strategies for small ruminants in the tropics. Ph.D. Thesis, Wageningen University, Gelderland, The Netherlands, 2004. [Google Scholar]
- Carolino, N.; Vicente, A.; Carolino, I. Genetic Improvement of Local Goats. In Sustainable Goat Production in Adverse Environments; Springer: Berlin/Heidelberg, Germany, 2017; Volume I, pp. 111–134. [Google Scholar] [CrossRef]
- Jotun, H.; Mikkel, S.; Carsten, W. Gene Genealogies, Variation and Evolution: A Primer in Coalescent Theory; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses under Linux and Windows. Mol. Ecol. Res. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Barbato, M.; Orozco-terWengel, P.; Tapio, M.; Bruford, M.W. SNEP: A Tool to Estimate Trends in Recent Effective Population Size Trajectories Using Genome-Wide SNP Data. Front. Genet. 2015, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Sved, J.A. Linkage Disequilibrium and Homozygosity of Chromosome Segments in Finite Populations. Theor. Popul. Biol. 1971, 2, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Hayes, B.J.; Visscher, P.M.; McPartlan, H.C.; Goddard, M.E. Novel Multilocus Measure of Linkage Disequilibrium to Estimate Past Effective Population Size. Genome Res. 2003, 13, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, B.; Zhang, T.; Yan, X.; Yu, Y.; Li, J.; Mei, B.; Wang, Z.; Zhang, Y.; Wang, R.; et al. Assessing Genetic Diversity and Estimating the Inbreeding Effect on Economic Traits of Inner Mongolia White Cashmere Goats through Pedigree Analysis. Front. Vet. Sci. 2021, 8, 665872. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing under Dependency. Ann. Statist. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Scherf, B.D., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2015. Available online: https://www.fao.org/3/i4787e/i4787e.pdf (accessed on 15 August 2023).
- Paim, T.; do Faria, D.A.; Hay, E.H.; McManus, C.; Lanari, M.R.; Esquivel, L.C.; Cascante, M.I.; Alfaro, E.J.; Mendez, A.; Faco, O.; et al. New World Goat Populations Are a Genetically Diverse Reservoir for Future Use. Sci. Rep. 2019, 9, 1476. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Iommelli, P.; Infascelli, L.; Raffrenato, E. Traditional Sources of Ingredients for the Food Industry: Animal Sources. In Sustainable Food Science—A Comprehensive Approach; Elsevier: Amsterdam, The Netherlands, 2023; pp. 7–20. [Google Scholar] [CrossRef]
- Massender, E.; Oliveira, H.R.; Brito, L.F.; Maignel, L.; Jafarikia, M.; Baes, C.F.; Sullivan, B.; Schenkel, F.S. Genome-Wide Association Study for Milk Production and Conformation Traits in Canadian Alpine and Saanen Dairy Goats. J. Dairy Sci. 2023, 106, 1168–1189. [Google Scholar] [CrossRef]
- Martin, P.; Palhière, I.; Maroteau, C.; Bardou, P.; Canale-Tabet, K.; Sarry, J.; Woloszyn, F.; Bertrand-Michel, J.; Racke, I.; Besir, H.; et al. A Genome Scan for Milk Production Traits in Dairy Goats Reveals Two New Mutations in DGAT1 Reducing Milk Fat Content. Sci. Rep. 2017, 7, 1872. [Google Scholar] [CrossRef] [PubMed]
- Koluman Darcan, N.; Silanikove, N. The Advantages of Goats for Future Adaptation to Climate Change: A Conceptual Overview. Small Rumin. Res. 2018, 163, 34–38. [Google Scholar] [CrossRef]
- Berihulay, H.; Abied, A.; He, X.; Jiang, L.; Ma, Y. Adaptation Mechanisms of Small Ruminants to Environmental Heat Stress. Animals 2019, 9, 75. [Google Scholar] [CrossRef]
- Zhang, L. Genetic Diversity of Five Goat Breeds in China Based on Microsatellite Markers. Afr. J. Biotechnol. 2012, 11, 11671–11677. [Google Scholar] [CrossRef]
- Wei, C.; Lu, J.; Xu, L.; Liu, G.; Wang, Z.; Zhao, F.; Zhang, L.; Han, X.; Du, L.; Liu, C. Genetic Structure of Chinese Indigenous Goats and the Special Geographical Structure in the Southwest China as a Geographic Barrier Driving the Fragmentation of a Large Population. PLoS ONE 2014, 9, e94435. [Google Scholar] [CrossRef]
- Muneerah, M.A.E.; Tamrin, N.A.M.; Salisi, M.S.; Zulkifly, S.; Ghazali, S.S.M.; Temuli, J.J.; Rosali, M.H.; Nazari, S.; Nik, W.M.K.W.; Mamat-Hamidi, K. Microsatellite-Based Genetic Characterization of the Indigenous Katjang Goat in Peninsular Malaysia. Animals 2021, 11, 1328. [Google Scholar] [CrossRef]
- Ellegren, H. Microsatellite Mutations in the Germline: Implications for evolutionary inference. Trends Genet. 2000, 16, 551–558. [Google Scholar] [CrossRef]
- Zimmerman, S.J.; Aldridge, C.L.; Oyler-McCance, S.J. An Empirical Comparison of Population Genetic Analyses Using Microsatellite and SNP Data for a Species of Conservation Concern. BMC Genom. 2020, 21, 382. [Google Scholar] [CrossRef]
- Morin, P.A.; Luikart, G.; Wayne, R.K.; the SNP Workshop Group. SNPs in Ecology, Evolution and Conservation. Trends Ecol. Evol. 2004, 19, 208–216. [Google Scholar] [CrossRef]
- Xing, C.; Schumacher, F.R.; Xing, G.; Lu, Q.; Wang, T.; Elston, R.C. Comparison of Microsatellites, Single-Nucleotide Polymorphisms (Snps) and Composite Markers Derived from Snps in Linkage Analysis. BMC Genet. 2005, 6, S29. [Google Scholar] [CrossRef]
- Kaiser, S.A.; Taylor, S.A.; Chen, N.; Sillett, T.S.; Bondra, E.R.; Webster, M.S. A Comparative Assessment of Snp and Microsatellite Markers for Assigning Parentage in a Socially Monogamous Bird. Mol. Ecol. Resour. 2016, 17, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Allendorf, F.W.; Luikart, G.H.; Aitken, S.N. Conservation and the Genetics of Populations; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Berihulay, H.; Islam, R.; Jiang, L.; Ma, Y. Genome-Wide Linkage Disequilibrium and the Extent of Effective Population Sizes in Six Chinese Goat Populations Using a 50k Single Nucleotide Polymorphism Panel. Animals 2019, 9, 350. [Google Scholar] [CrossRef]
- Biscarini, F.; Cozzi, P.; Orozco-ter Wengel, P. Lessons Learnt on the Analysis of Large Sequence Data in Animal Genomics. Anim. Genet. 2018, 49, 147–158. [Google Scholar] [CrossRef]
- Michailidou, S.; Tsangaris, G.T.; Tzora, A.; Skoufos, I.; Banos, G.; Argiriou, A.; Arsenos, G. Analysis of Genome-Wide DNA Arrays Reveals the Genomic Population Structure and Diversity in Autochthonous Greek Goat Breeds. PLoS ONE 2019, 14, e0226179. [Google Scholar] [CrossRef]
- Brito, L.F.; Jafarikia, M.; Grossi, D.A.; Kijas, J.W.; Porto-Neto, L.R.; Ventura, R.V.; Salgorzaei, M.; Schenkel, F.S. Characterization of Linkage Disequilibrium, Consistency of Gametic Phase and Admixture in Australian and Canadian Goats. BMC Genet. 2015, 16, 67. [Google Scholar] [CrossRef]
- Mucha, S.; Mrode, R.; MacLaren-Lee, I.; Coffey, M.; Conington, J. Estimation of Genomic Breeding Values for Milk Yield in UK Dairy Goats. J. Dairy Sci. 2015, 98, 8201–8208. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lu, J.; Fei, X.; Lu, Z.; Quan, K.; Liu, Y.; Chu, M.; Di, R.; Wei, C.; Wang, H. Selection Signatures Analysis Reveals Genes Associated with High-Altitude Adaptation in Tibetan Goats from Nagqu, Tibet. Animals 2020, 10, 1599. [Google Scholar] [CrossRef]
- Hall, S.J. Genetic Differentiation among Livestock Breeds—Values for Fst. Animals 2022, 12, 1115. [Google Scholar] [CrossRef]
- Cappuccio, I.; Pariset, L.; Ajmone-Marsan, P.; Dunner, S.; Cortes, O.; Erhardt, G.; Lühken, G.; Gutscher, K.; Joost, S.; Nijman, I.J.; et al. Allele Frequencies and Diversity Parameters of 27 Single Nucleotide Polymorphisms within and across Goat Breeds. Mol. Ecol. Notes 2006, 6, 992–997. [Google Scholar] [CrossRef]
- Kawęcka, A.; Podbielska, A.; Miksza-Cybulska, A.; Pasternak, M.; Sikora, J.; Szmatoła, T. Genetic Structure of Reconstituted Native Carpathian Goat Breed Based on Information from Microsatellite Markers. Ann. Anim. Sci. 2022, 22, 1235–1244. [Google Scholar] [CrossRef]
- Juler, C. După Coada Oilor: Long-Distance Transhumance and Its Survival in Romania. Pastor. Res. Policy Pract. 2014, 4, 4. [Google Scholar] [CrossRef]
- Islam, R.; Liu, Z.; Li, Y.; Jiang, L.; Ma, Y. Conservation Assessment of the State Goat Farms by Using SNP Genotyping Data. Genes 2020, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Franklin, I.R. Evolutionary change in small populations. In Conservation Biology: An Evolutionary-Ecological Perspective; Soulé, M.E., Wilcox, B.M., Eds.; Sinauer Associates Inc.: Sunderland, MA, USA, 1980; pp. 135–149. [Google Scholar]
- Laikre, L.; Hohenlohe, P.A.; Allendorf, F.W.; Bertola, L.D.; Breed, M.F.; Bruford, M.W.; Funk, W.C.; Gajardo, G.; González-Rodríguez, A.; Grueber, C.E.; et al. Authors’ Reply to Letter to the Editor: Continued Improvement to Genetic Diversity Indicator for CBD. Conserv. Genet. 2021, 22, 533–536. [Google Scholar] [CrossRef]
- Hoban, S.; da Silva, J.M.; Mastretta-Yanes, A.; Grueber, C.E.; Heuertz, M.; Hunter, M.E.; Mergeay, J.; Paz-Vinas, I.; Fukaya, K.; Ishihama, F.; et al. Monitoring Status and Trends in Genetic Diversity for the Convention on Biological Diversity: An Ongoing Assessment of Genetic Indicators in Nine Countries. Conserv. Lett. 2023, 16, e12953. [Google Scholar] [CrossRef]
- Meuwissen, T. Genetic Management of Small Populations: A Review. Acta Agric. Scand. Sect. A Anim. Sci. 2009, 59, 71–79. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Sample Code | Sample Size (n) | Sampling Area (County) | AAT **/ALT *** |
|---|---|---|---|
| HA | 22 | SV, SB, CV, BV, AG * | 355 m–1241 m |
| LA | 22 | NT, BR, TL, CT, OT, TR | 0 m–188 m |
| HT | 33 | BR, TL, CT, TR | 12 °C–13.1 °C |
| LT | 17 | CV, BV | 7.1 °C–7.6 °C |
| Group Acronym | n | PI-HAT | MAF | Ho | HE | FIS |
|---|---|---|---|---|---|---|
| HA | 22 | 0.007 | 0.331 | 0.416 | 0.416 | 0.021 |
| LA | 22 | 0.004 | 0.333 | 0.419 | 0.418 | 0.019 |
| HT | 33 | 0.008 | 0.335 | 0.418 | 0.420 | 0.020 |
| LT | 17 | 0.007 | 0.328 | 0.416 | 0.413 | 0.022 |
| Overall | 67 | 0.007 | 0.332 | 0.418 | 0.418 | 0.021 |
| Group Symbol | HA | LA | HT | LT |
|---|---|---|---|---|
| HA | 0.000 | 0.000 | 0.982 | |
| LA | 0.007 | 0.991 | 0.000 | |
| HT | 0.006 | 0.000 | 0.000 | |
| LT | 0.000 | 0.006 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlaic, B.A.; Vlaic, A.; Russo, I.-R.; Colli, L.; Bruford, M.W.; Odagiu, A.; Orozco-terWengel, P.; CLIMGEN Consortium. Analysis of Genetic Diversity in Romanian Carpatina Goats Using SNP Genotyping Data. Animals 2024, 14, 560. https://doi.org/10.3390/ani14040560
Vlaic BA, Vlaic A, Russo I-R, Colli L, Bruford MW, Odagiu A, Orozco-terWengel P, CLIMGEN Consortium. Analysis of Genetic Diversity in Romanian Carpatina Goats Using SNP Genotyping Data. Animals. 2024; 14(4):560. https://doi.org/10.3390/ani14040560
Chicago/Turabian StyleVlaic, Bogdan Alin, Augustin Vlaic, Isa-Rita Russo, Licia Colli, Michael William Bruford, Antonia Odagiu, Pablo Orozco-terWengel, and CLIMGEN Consortium. 2024. "Analysis of Genetic Diversity in Romanian Carpatina Goats Using SNP Genotyping Data" Animals 14, no. 4: 560. https://doi.org/10.3390/ani14040560