
How Is the Nociceptive Withdrawal Reflex Influenced by Increasing Doses of Propofol in Pigs?

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
Statistic
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Egan, T.D. Are Opioids Indispensable for General Anaesthesia? Br. J. Anaesth. 2019, 122, e127–e135. [Google Scholar] [CrossRef] [PubMed]
- Mirra, A.; Maidanskaia, E.G.; Carmo, L.P.; Levionnois, O.; Spadavecchia, C. How Is Depth of Anaesthesia Assessed in Experimental Pigs? A Scoping Review. PLoS ONE 2023, 18, e0283511. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Dingley, J.; Elstad, M.; Scull-Brown, E.; Steen, P.A.; Thoresen, M. Minimum Alveolar Concentration (MAC) for Sevoflurane and Xenon at Normothermia and Hypothermia in Newborn Pigs. Acta Anaesthesiol. Scand. 2013, 57, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Valverde, A.; Morey, T.E.; Hernández, J.; Davies, W. Validation of Several Types of Noxious Stimuli for Use in Determining the Minimum Alveolar Concentration for Inhalation Anesthetics in Dogs and Rabbits. Am. J. Vet. Res. 2003, 64, 957–962. [Google Scholar] [CrossRef] [PubMed]
- Antognini, J.F.; Barter, L.; Carstens, E. Movement as an Index of Anesthetic Depth in Humans and Experimental Animals. Comp. Med. 2005, 55, 413–418. [Google Scholar] [PubMed]
- Whelan, G.; Flecknell, P.A. The Assessment of Depth of Anaesthesia in Animals and Man. Lab. Anim. 1992, 26, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kugelberg, E.; Eklund, K.; Grimby, L. An Electromyographic Study of the Nociceptive Reflexes of the Lower Limb. Mechanism of the Plantar Responses. Brain 1960, 83, 394–410. [Google Scholar] [CrossRef] [PubMed]
- Hagbarth, K.E. Spinal Withdrawal Reflexes in the Human Lower Limbs. J. Neurol. Neurosurg. Psychiatry 1960, 23, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Baars, J.H.; Rintisch, U.; Rehberg, B.; Lahrmann, K.H.; von Dincklage, F. Prediction of Motor Responses to Surgical Stimuli during Bilateral Orchiectomy of Pigs Using Nociceptive Flexion Reflexes and the Bispectral Index Derived from the Electroencephalogram. Vet. J. 2013, 195, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Von Dincklage, F.; Hackbarth, M.; Mager, R.; Rehberg, B.; Baars, J.H. Monitoring of the Responsiveness to Noxious Stimuli during Anaesthesia with Propofol and Remifentanil by Using RIII Reflex Threshold and Bispectral Index. Br. J. Anaesth. 2010, 104, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Von Dincklage, F.; Velten, H.; Rehberg, B.; Baars, J.H. Monitoring of the Responsiveness to Noxious Stimuli during Sevoflurane Mono-Anaesthesia by Using RIII Reflex Threshold and Bispectral Index. Br. J. Anaesth. 2010, 104, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Von Dincklage, F.; Send, K.; Hackbarth, M.; Rehberg, B.; Baars, J.H. Comparison of the Nociceptive Flexion Reflex Threshold and the Bispectral Index as Monitors of Movement Responses to Noxious Stimuli under Propofol Mono-Anaesthesia. Br. J. Anaesth. 2009, 102, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Jinks, S.L.; Martin, J.T.; Carstens, E.; Jung, S.W.; Antognini, J.F. Peri-MAC Depression of a Nociceptive Withdrawal Reflex Is Accompanied by Reduced Dorsal Horn Activity with Halothane but Not Isoflurane. Anesthesiology 2003, 98, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jin, T.; Qin, B.; Turndorf, H. Suppression of Spinal Cord Motoneuron Excitability Correlates with Surgical Immobility during Isoflurane Anesthesia. Anesthesiology 1998, 88, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Antognini, J.F.; Carstens, E.; Buzin, V. Isoflurane Depresses Motoneuron Excitability by a Direct Spinal Action: An F-Wave Study. Anesth. Analg. 1999, 88, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Antognini, J.F.; Atherley, R.; Carstens, E. Isoflurane Action in Spinal Cord Indirectly Depresses Cortical Activity Associated with Electrical Stimulation of the Reticular Formation. Anesth. Analg. 2003, 96, 999–1003. [Google Scholar] [CrossRef]
- Antognini, J.F.; Wang, X.W.; Carstens, E. Isoflurane Action in the Spinal Cord Blunts Electroencephalographic and Thalamic-Reticular Formation Responses to Noxious Stimulation in Goats. Anesthesiology. 2000, 92, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Antognini, J.; Wang, X.W. Isoflurane Indirectly Depresses Middle Latency Auditory Evoked Poten- Tials by Action in the Spinal Cord in the Goat. Can. J. Anesth. 1999, 46, 692–695. [Google Scholar] [CrossRef]
- Biurrun Manresa, J.A.; Neziri, A.Y.; Curatolo, M.; Arendt-Nielsen, L.; Andersen, O.K. Test-Retest Reliability of the Nociceptive Withdrawal Reflex and Electrical Pain Thresholds after Single and Repeated Stimulation in Patients with Chronic Low Back Pain. Eur. J. Appl. Physiol. 2011, 111, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Herm, C.; Silbereisen, V.; Graf, B.M.; Lassen, C.L. Long Term Reliability of Nociceptive Withdrawal Reflex Thresholds. J. Neurosci. Methods 2019, 320, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Arifin, W.N.; Zahiruddin, W.M. Sample Size Calculation in Animal Studies Using Resource Equation Approach. Malays. J. Med. Sci. 2017, 24, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Diez Bernal, S.; Studer, N.; Thormann, W.; Spadavecchia, C.; Levionnois, O. Pharmacokinetic–Pharmacodynamic Modelling of the Antinociceptive Effect of a Romifidine Infusion in Standing Horses. Vet. Anaesth. Analg. 2020, 47, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Siegenthaler, J.; Pleyers, T.; Raillard, M.; Spadavecchia, C.; Levionnois, O.L. Effect of Medetomidine, Dexmedetomidine, and Their Reversal with Atipamezole on the Nociceptive Withdrawal Reflex in Beagles. Animals 2020, 10, 1240. [Google Scholar] [CrossRef] [PubMed]
- Von Dincklage, F.; Hackbarth, M.; Schneider, M.; Baars, J.H.; Rehberg, B. Introduction of a Continual RIII Reflex Threshold Tracking Algorithm. Brain Res. 2009, 1260, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Rhudy, J.L.; France, C.R. Defining the Nociceptive Flexion Reflex (NFR) Threshold in Human Participants: A Comparison of Different Scoring Criteria. Pain 2007, 128, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Mirra, A.; Casoni, D.; Barge, P.; Hight, D.; Levionnois, O.; Spadavecchia, C. Usability of the SedLine® Electroencephalographic Monitor of Depth of Anaesthesia in Pigs: A Pilot Study. J. Clin. Monit. Comput. 2022, 36, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Egan, T.D.; Kern, S.E.; Johnson, K.B.; Pace, N.L. The Pharmacokinetics and Pharmacodynamics of Propofol in a Modified Cyclodextrin Formulation (Captisol®) versus Propofol in a Lipid Formulation (Diprivan®): An Electroencephalographic and Hemodynamic Study in a Porcine Model. Anesth. Analg. 2003, 97, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Peacock, J.E.; Lewis, R.P.; Reilly, C.S.; Nimmo, W.S. Effect of Different Rates of Infusion of Propofol for Induction of Anaesthesia in Elderly Patients. Br. J. Anaesth. 1990, 65, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Stokes, D.N.; Hutton, P. Rate-Dependent Induction Phenomena with Propofol: Implications for the Relative Potency of Intravenous Anesthetics. Anesth. Analg. 1991, 72, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Le Bars, D.; Dickenson, A.H.; Besson, J.M. Diffuse Noxious Inhibitory Controls (DNIC). I. Effects on Dorsal Horn Convergent Neurones in the Rat. Pain 1979, 6, 283–304. [Google Scholar] [CrossRef] [PubMed]
- Dickenson, A.H.; Bars, D.L.E.; Besson, J.M. Diffuse Noxious Inhibitory Controls (DNIC). Effects on Trigeminal Nucleus Caudalis Neurones in the Rat. Brain Res. 1980, 200, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Antognini, J.F.; Carstens, E. In Vivo Characterization of Clinical Anaesthesia and Its Components. Br. J. Anaesth. 2002, 89, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Lichtner, G.; Auksztulewicz, R.; Velten, H.; Mavrodis, D.; Scheel, M.; Blankenburg, F.; von Dincklage, F. Nociceptive Activation in Spinal Cord and Brain Persists during Deep General Anaesthesia. Br. J. Anaesth. 2018, 121, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Zacny, J.P.; Coalson, D.W.; Young, C.J.; Klafta, J.M.; Lichtor, J.L.; Rupani, G.; Thapar, P.; Apfelbaum, J.L. Propofol at Conscious Sedation Doses Produces Mild Analgesia to Cold Pressor-Induced Pain in Healthy Volunteers. J. Clin. Anesth. 1996, 8, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Baars, J.H.; Mager, R.; Dankert, K.; Hackbarth, M.; Von Dincklage, F.; Rehberg, B. Effects of Sevoflurane and Propofol on the Nociceptive Withdrawal Reflex and on the H Reflex. Anesthesiology 2009, 111, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Bandschapp, O.; Filitz, J.; Ihmsen, H.; Berset, A.; Urwyler, A.; Koppert, W.; Ruppen, W. Analgesic and Antihyperalgesic Properties of Propofol in a Human Pain Model. Anesthesiology 2010, 113, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Takechi, K.; Cartens, M.; Klein, A.; Cartens, E. The Antinociceptive and Antihyperalgesic Effects of Topical Propofol on Dorsal Horn Neurons in the Rat. Anesth. Analg. 2008, 116, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Mylius, V.; Kunz, M.; Schepelmann, K.; Lautenbacher, S. Sex Differences in Nociceptive Withdrawal Reflex and Pain Perception. Somatosens. Mot. Res. 2005, 22, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Neziri, A.Y.; Andersen, O.K.; Petersen-Felix, S.; Radanov, B.; Dickenson, A.H.; Scaramozzino, P.; Arendt-Nielsen, L.; Curatolo, M. The Nociceptive Withdrawal Reflex: Normative Values of Thresholds and Reflex Receptive Fields. Eur. J. Pain 2010, 14, 134–141. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
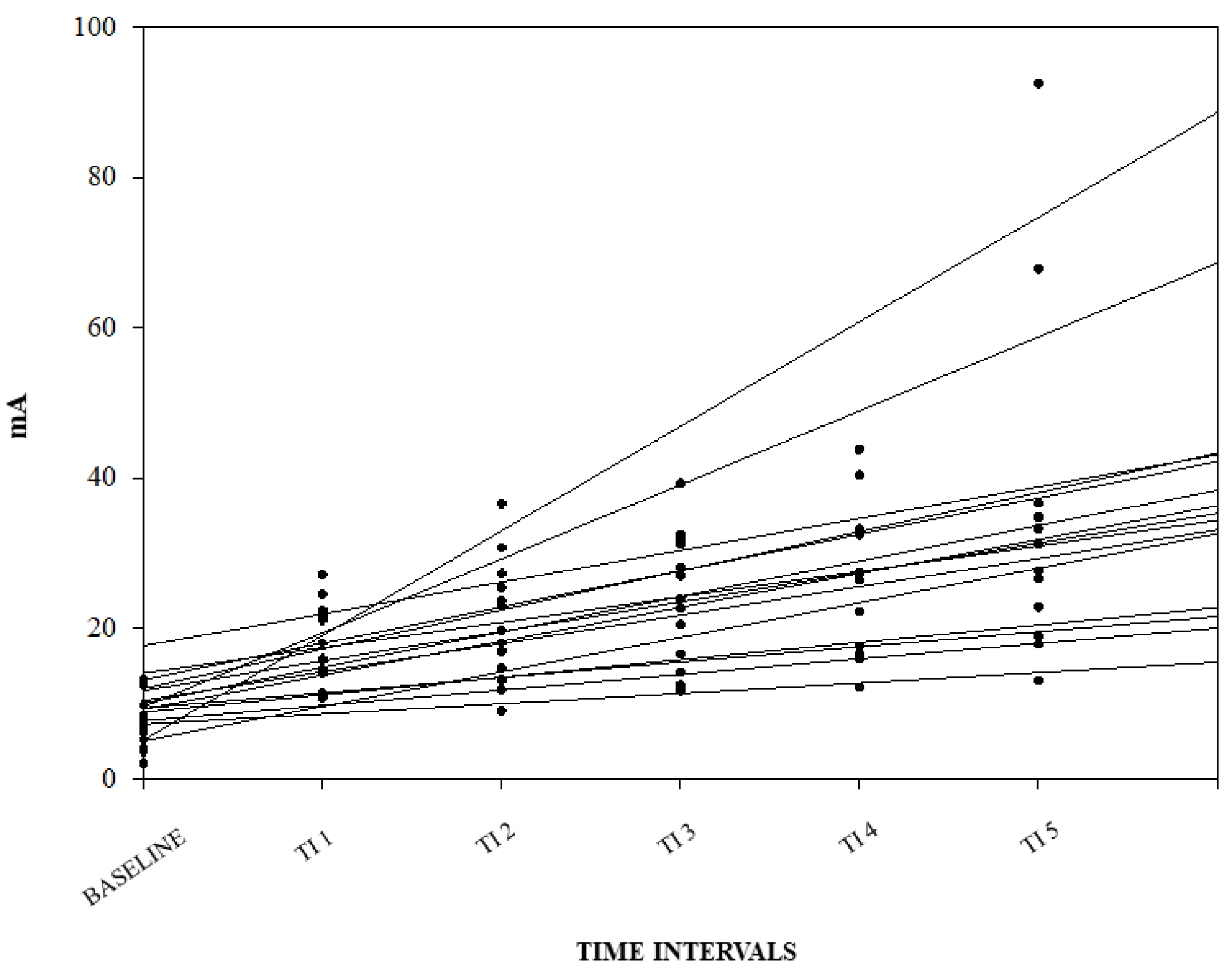
| NWRt (mA) | |||
|---|---|---|---|
| Median | IQ 25% | IQ 75% | |
| Baseline | 7.2 | 4.9 | 10.5 |
| TI1 | 16.9 | 13.3 | 22.1 |
| TI2 | 18.8 | 14.2 | 25.8 |
| TI3 | 23.2 | 15.9 | 31.4 |
| TI4 | 26.7 | 17.3 | 32.9 |
| TI5 | 29.4 | 21.8 | 35.3 |
| Intubation | 30.9 | 20.1 | 34.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirra, A.; Gamez Maidanskaia, E.; Levionnois, O.L.; Spadavecchia, C. How Is the Nociceptive Withdrawal Reflex Influenced by Increasing Doses of Propofol in Pigs? Animals 2024, 14, 1081. https://doi.org/10.3390/ani14071081
Mirra A, Gamez Maidanskaia E, Levionnois OL, Spadavecchia C. How Is the Nociceptive Withdrawal Reflex Influenced by Increasing Doses of Propofol in Pigs? Animals. 2024; 14(7):1081. https://doi.org/10.3390/ani14071081
Chicago/Turabian StyleMirra, Alessandro, Ekaterina Gamez Maidanskaia, Olivier Louis Levionnois, and Claudia Spadavecchia. 2024. "How Is the Nociceptive Withdrawal Reflex Influenced by Increasing Doses of Propofol in Pigs?" Animals 14, no. 7: 1081. https://doi.org/10.3390/ani14071081