
Emotional “Contagion” in Piglets after Sensory Avoidance of Rewarding and Punishing Treatment


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Management
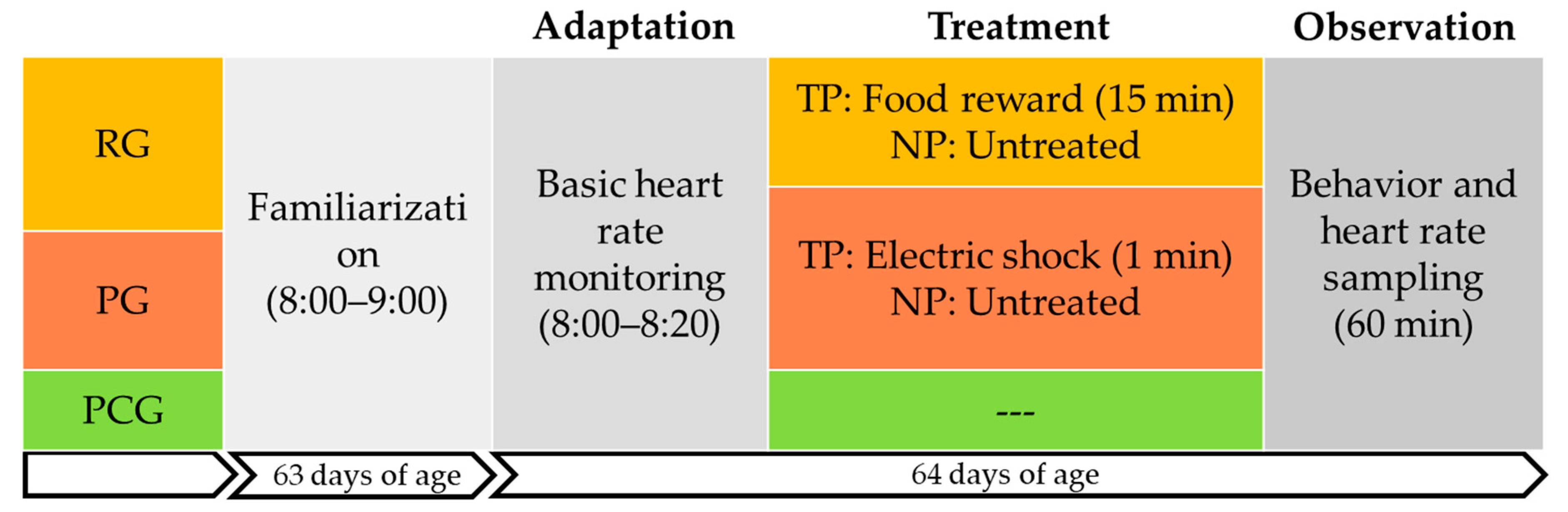
2.2. Experimental Animal Grouping
2.3. Treatments
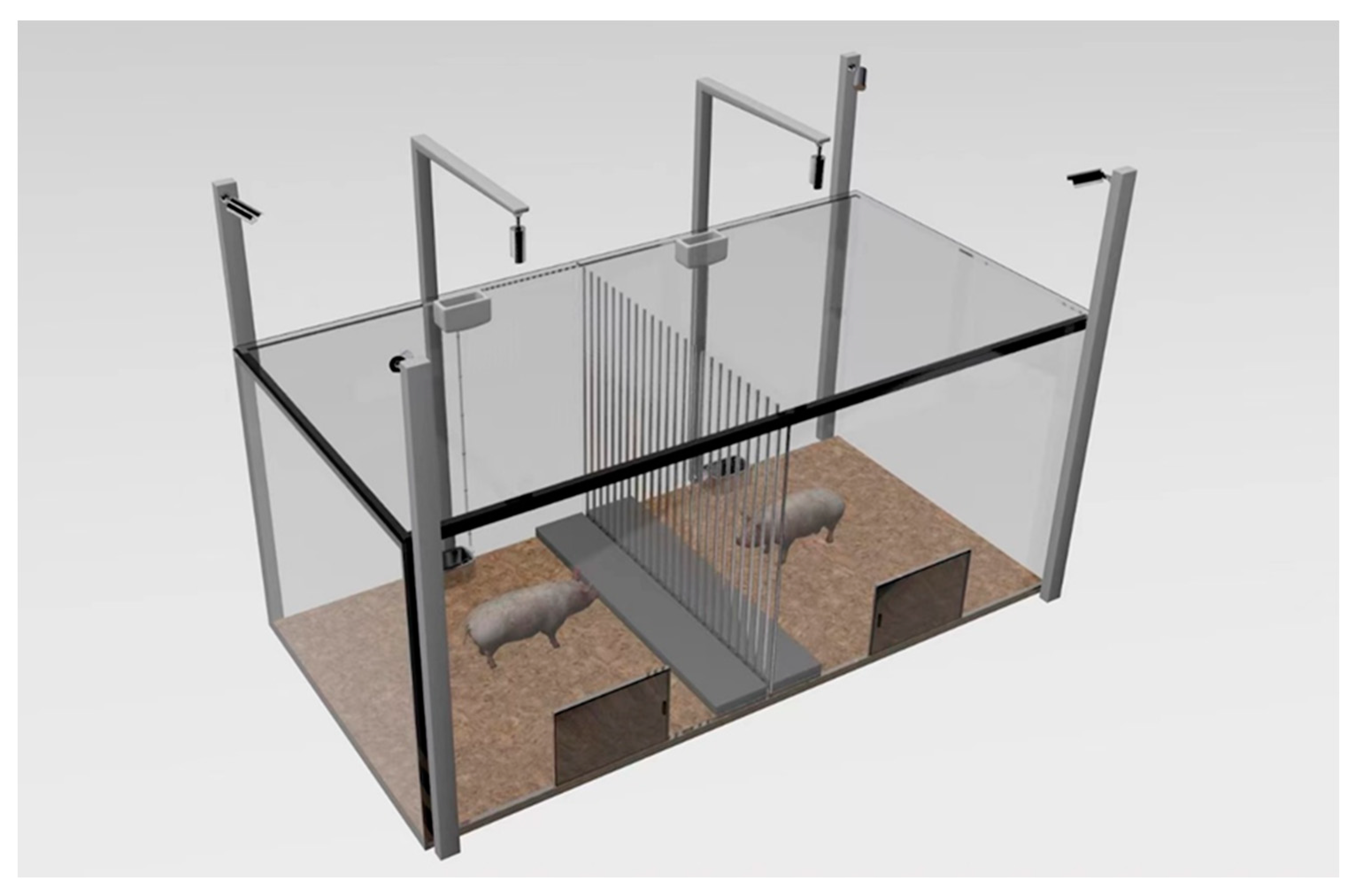
2.4. Behavioral Observation
2.5. Heart Rate Test
2.6. Data Analysis
3. Results
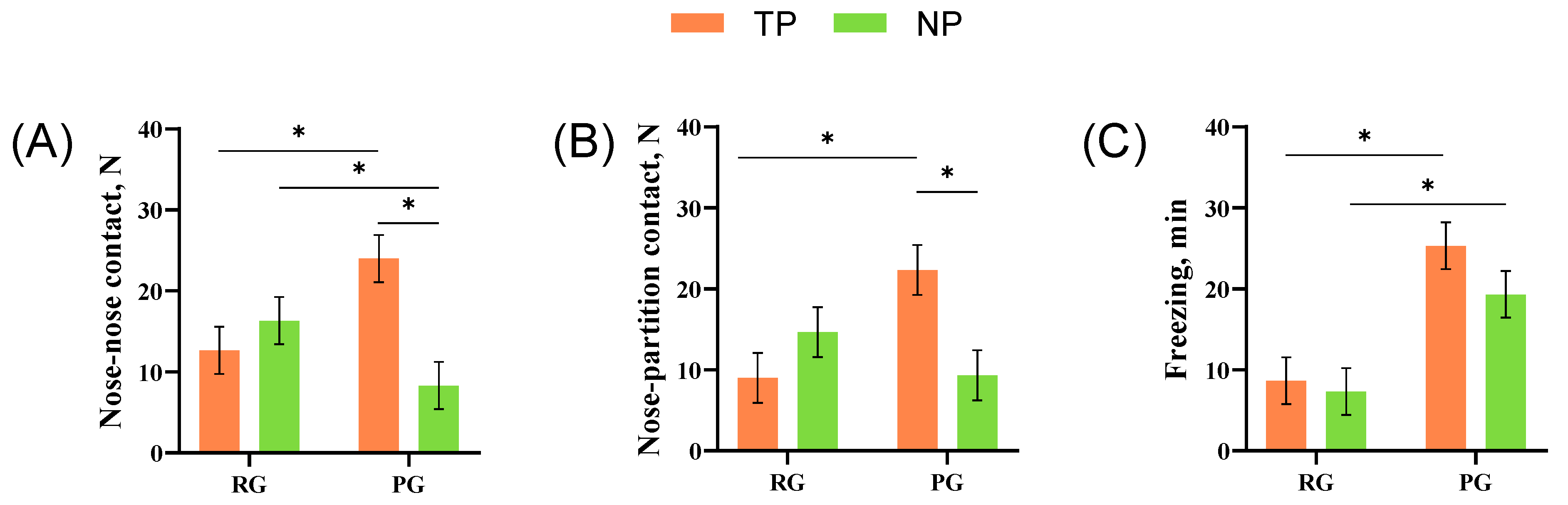
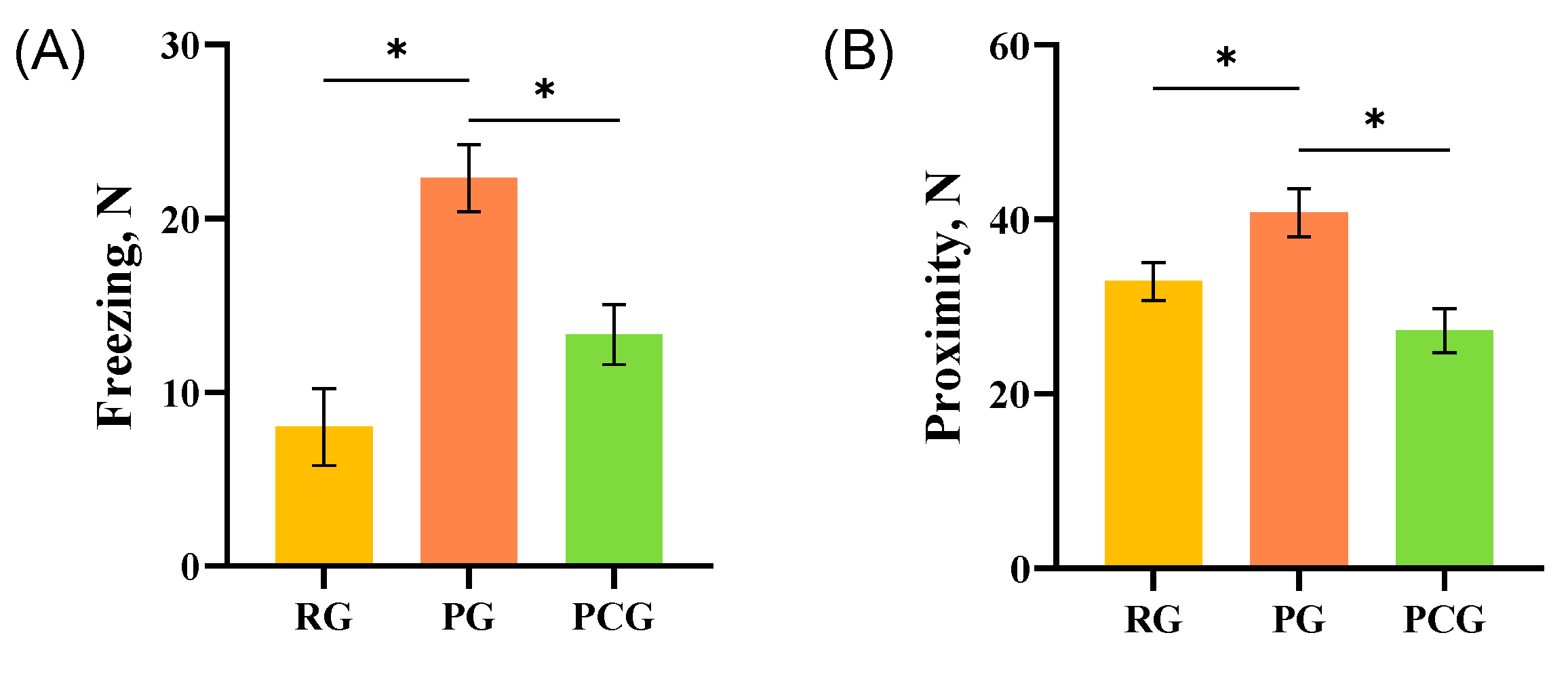
3.1. Behavioral Performance
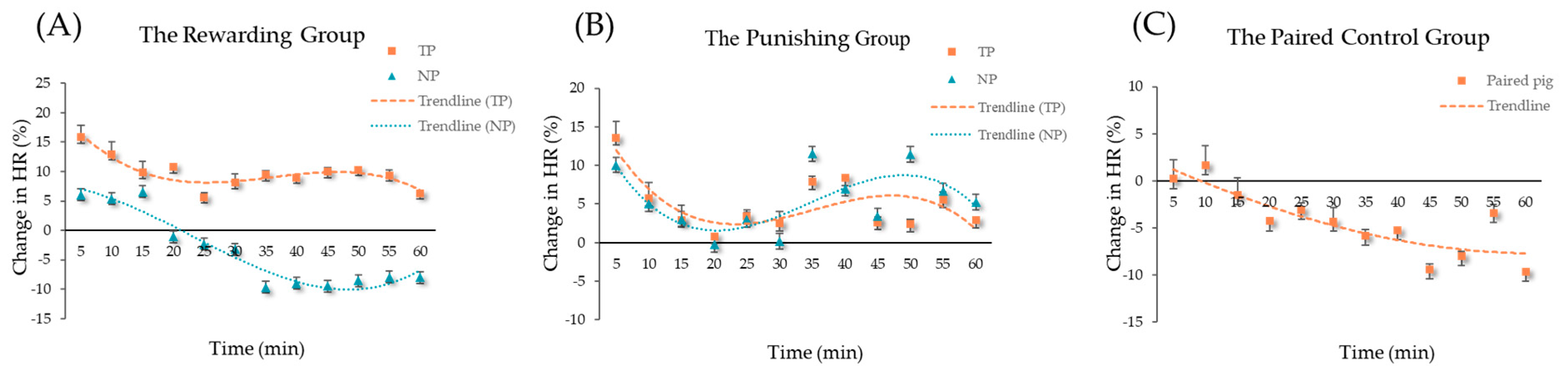
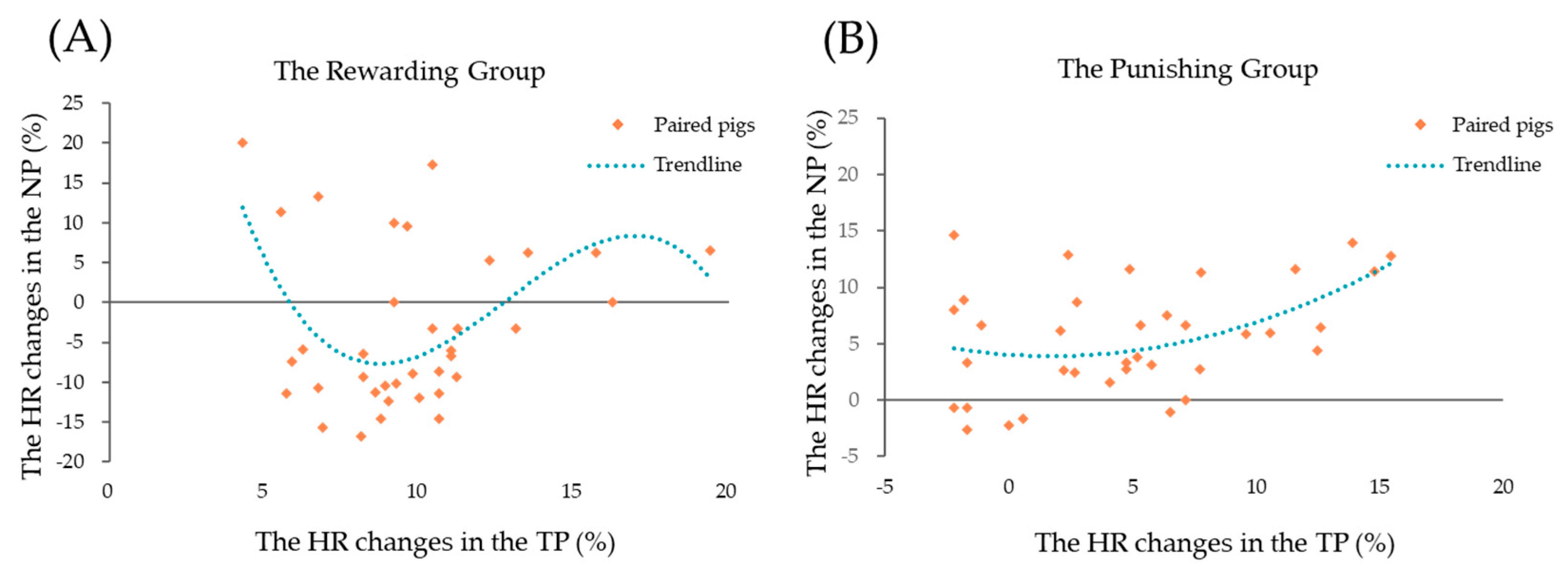
3.2. Regression of Heart Rate
4. Discussion
4.1. Reactions of Treated Pigs in Punishment and Reward Groups
4.2. Emotional Contagion and Social Interactions between Treated and Naive Pigs
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bonastre, C.; Mitjana, O.; Tejedor, M.T.; Calavia, M.; Yuste, A.G.; Ubeda, J.L.; Falceto, M.V. Acute Physiological Responses to Castration-Related Pain in Piglets: The Effect of Two Local Anesthetics with or without Meloxicam. Animal 2016, 10, 1474–1481. [Google Scholar] [CrossRef]
- Camerlink, I.; Farish, M.; D’Eath, R.; Arnott, G.; Turner, S. Long Term Benefits on Social Behaviour after Early Life Socialization of Piglets. Animals 2018, 8, 192. [Google Scholar] [CrossRef]
- Hakansson, F.; Lahrmann, H.P.; Forkman, B. A Longitudinal Study of Pre- and Post-Weaning Tail Damage in Non-Docked Pigs. Animal 2020, 14, 2159–2166. [Google Scholar] [CrossRef] [PubMed]
- Krugmann, K.L.; Mieloch, F.J.; Krieter, J.; Czycholl, I. Investigation of Influence of Growing Pigs’ Positive Affective State on Behavioral and Physiological Parameters Using Structural Equation Modeling. J. Anim. Sci. 2020, 98, skaa028. [Google Scholar] [CrossRef]
- Baciadonna, L.; Briefer, E.F.; Favaro, L.; McElligott, A.G. Goats Distinguish between Positive and Negative Emotion-Linked Vocalisations. Front. Zool. 2019, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, O.; O’Driscoll, K.; Baxter, E.M.; Boyle, L.A. Artificial Rearing Affects the Emotional State and Reactivity of Pigs Post-Weaning. Anim. Welf. 2019, 28, 433–442. [Google Scholar] [CrossRef]
- Marcet-Rius, M.; Kalonji, G.; Cozzi, A.; Bienboire-Frosini, C.; Monneret, P.; Kowalczyk, I.; Teruel, E.; Codecasa, E.; Pageat, P. Effects of Straw Provision, as Environmental Enrichment, on Behavioural Indicators of Welfare and Emotions in Pigs Reared in an Experimental System. Livest. Sci. 2019, 221, 89–94. [Google Scholar] [CrossRef]
- Imfeld-Mueller, S.; Van Wezemael, L.; Stauffacher, M.; Gygax, L.; Hillmann, E. Do Pigs Distinguish between Situations of Different Emotional Valences during Anticipation? Appl. Anim. Behav. Sci. 2011, 131, 86–93. [Google Scholar] [CrossRef]
- Krebs, N.; McGlone, J.J. Effects of Exposing Pigs to Moving and Odors in a Simulated Slaughter Chute. Appl. Anim. Behav. Sci. 2009, 116, 179–185. [Google Scholar] [CrossRef]
- Reimert, I.; Bolhuis, J.E.; Kemp, B.; Rodenburg, T.B. Emotions on the Loose: Emotional Contagion and the Role of Oxytocin in Pigs. Anim. Cogn. 2015, 18, 517–532. [Google Scholar] [CrossRef]
- Reimert, I.; Bolhuis, J.E.; Kemp, B.; Rodenburg, T.B. Indicators of Positive and Negative Emotions and Emotional Contagion in Pigs. Physiol. Behav. 2013, 109, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Edgar, J.L.; Nicol, C.J.; Clark, C.C.A.; Paul, E.S. Measuring Empathic Responses in Animals. Appl. Anim. Behav. Sci. 2012, 138, 182–193. [Google Scholar] [CrossRef]
- Keum, S.; Kim, A.; Shin, J.J.; Kim, J.-H.; Park, J.; Shin, H.-S. A Missense Variant at the Nrxn3 Locus Enhances Empathy Fear in the Mouse. Neuron 2018, 98, 588–601.e5. [Google Scholar] [CrossRef] [PubMed]
- de Waal, F.B.M. Putting the Altruism Back into Altruism: The Evolution of Empathy. Annu. Rev. Psychol. 2008, 59, 279–300. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Takahashi, Y.; Matsumoto, Y.; Okamoto, M.; Ishihara, T. Conformity-like Behaviour in Mice Observing the Freezing of Other Mice: A Model of Empathy. BMC Neurosci. 2020, 21, 19. [Google Scholar] [CrossRef] [PubMed]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Okamoto, M.; Matsumoto, Y.; Aoki, S.; Ishihara, T. Empathic Behavior According to the State of Others in Mice. Brain Behav. 2018, 8, e00986. [Google Scholar] [CrossRef] [PubMed]
- Edgar, J.L.; Paul, E.S.; Nicol, C.J. Protective Mother Hens: Cognitive Influences on the Avian Maternal Response. Anim. Behav. 2013, 86, 223–229. [Google Scholar] [CrossRef]
- Marino, L. Thinking Chickens: A Review of Cognition, Emotion, and Behavior in the Domestic Chicken. Anim. Cogn. 2017, 20, 127–147. [Google Scholar] [CrossRef]
- Groessbacher, V.; Lawrence, A.B.; Winckler, C.; Spinka, M. Negative Play Contagion in Calves. Sci. Rep. 2020, 10, 21699. [Google Scholar] [CrossRef]
- Buchanan, T.W.; Preston, S.D. Commentary: Social Stress Contagion in Rats: Behavioral, Autonomic and Neuroendocrine Correlates. Front. Behav. Neurosci. 2017, 11, 175. [Google Scholar] [CrossRef]
- Goumon, S.; Špinka, M. Emotional Contagion of Distress in Young Pigs Is Potentiated by Previous Exposure to the Same Stressor. Anim. Cogn. 2016, 19, 501–511. [Google Scholar] [CrossRef]
- Marcet-Rius, M.; Pageat, P.; Bienboire-Frosini, C.; Teruel, E.; Monneret, P.; Leclercq, J.; Cozzi, A. The Provision of Toys to Pigs Can Improve the Human-Animal Relationship. Porc. Health Manag. 2020, 6, 29. [Google Scholar] [CrossRef]
- Reimert, I.; Fong, S.; Rodenburg, T.B.; Bolhuis, J.E. Emotional States and Emotional Contagion in Pigs after Exposure to a Positive and Negative Treatment. Appl. Anim. Behav. Sci. 2017, 193, 37–42. [Google Scholar] [CrossRef]
- Russell, J.A. Core Affect and the Psychological Construction of Emotion. Psychol. Rev. 2003, 110, 145–172. [Google Scholar] [CrossRef]
- Camerlink, I.; Reimert, I.; Bolhuis, J.E. Intranasal Oxytocin Administration in Relationship to Social Behaviour in Domestic Pigs. Physiol. Behav. 2016, 163, 51–55. [Google Scholar] [CrossRef]
- Reimert, I.; Bolhuis, J.E.; Kemp, B.; Rodenburg, T.B. Social Support in Pigs with Different Coping Styles. Physiol. Behav. 2014, 129, 221–229. [Google Scholar] [CrossRef]
- Tallet, C.; Rakotomahandry, M.; Herlemont, S.; Prunier, A. Evidence of Pain, Stress, and Fear of Humans During Tail Docking and the Next Four Weeks in Piglets (Sus Scrofa Domesticus). Front. Vet. Sci. 2019, 6, 462. [Google Scholar] [CrossRef]
- Zupan, M.; Zanella, A.J. Peripheral Regulation of Stress and Fear Responses in Pigs from Tail-Biting Pens. R. Bras. Zootec. 2017, 46, 33–38. [Google Scholar] [CrossRef]
- Düpjan, S.; Tuchscherer, A.; Langbein, J.; Schön, P.-C.; Manteuffel, G.; Puppe, B. Behavioural and Cardiac Responses towards Conspecific Distress Calls in Domestic Pigs (Sus Scrofa). Physiol. Behav. 2011, 103, 445–452. [Google Scholar] [CrossRef]
- Correa, E.K.; Bianchi, I.; Perondi, A.; de los Santos, J.R.G.; Correa, M.N.; Castilhos, D.D.; Gil-Turnes, C.; Lucia, T. Chemical and Microbiological Characteristics of Rice Husk Bedding Having Distinct Depths and Used for Growing-Finishing Swine. Bioresour. Technol. 2009, 100, 5318–5322. [Google Scholar] [CrossRef]
- Measures for Ethical Review of Science and Technology. Available online: https://www.most.gov.cn/xxgk/xinxifenlei/fdzdgknr/fgzc/gfxwj/gfxwj2023/202310/t20231008_188309.html (accessed on 11 March 2024).
- Zhang, J.; Critchley, L.A.H.; Lee, D.C.W.; Khaw, K.S.; Lee, S.W.Y. The Effect of Head up Tilting on Bioreactance Cardiac Output and Stroke Volume Readings Using Suprasternal Transcutaneous Doppler as a Control in Healthy Young Adults. J. Clin. Monit. Comput. 2016, 30, 519–526. [Google Scholar] [CrossRef]
- Fureix, C.; Meagher, R.K. What Can Inactivity (in Its Various Forms) Reveal about Affective States in Non-Human Animals? A Review. Appl. Anim. Behav. Sci. 2015, 171, 8–24. [Google Scholar] [CrossRef]
- Baciadonna, L.; Duepjan, S.; Briefer, E.F.; de la Torre, M.P.; Nawroth, C. Looking on the Bright Side of Livestock Emotions-the Potential of Their Transmission to Promote Positive Welfare. Front. Vet. Sci. 2018, 5, 218. [Google Scholar] [CrossRef]
- Keysers, C.; Knapska, E.; Moita, M.A.; Gazzola, V. Emotional Contagion and Prosocial Behavior in Rodents. Trends Cogn. Sci. 2022, 26, 688–706. [Google Scholar] [CrossRef]
- Adriaense, J.E.C.; Martin, J.S.; Schiestl, M.; Lamm, C.; Bugnyar, T. Negative Emotional Contagion and Cognitive Bias in Common Ravens (Corvus Corax). Proc. Natl. Acad. Sci. USA 2019, 116, 11547–11552. [Google Scholar] [CrossRef]
- Jeon, D.; Kim, S.; Chetana, M.; Jo, D.; Ruley, H.E.; Lin, S.-Y.; Rabah, D.; Kinet, J.-P.; Shin, H.-S. Observational Fear Learning Involves Affective Pain System and Cav1.2 Ca2+ Channels in ACC. Nat. Neurosci. 2010, 13, 482–488. [Google Scholar] [CrossRef]
- Sanders, J.; Mayford, M.; Jeste, D. Empathic Fear Responses in Mice Are Triggered by Recognition of a Shared Experience. PLoS ONE 2013, 8, e74609. [Google Scholar] [CrossRef]
- de Waal, F.B.M.; Preston, S.D. Mammalian Empathy: Behavioural Manifestations and Neural Basis. Nat. Rev. Neurosci. 2017, 18, 498–509. [Google Scholar] [CrossRef]
- Knapska, E.; Mikosz, M.; Werka, T.; Maren, S. Social Modulation of Learning in Rats. Learn. Mem. 2010, 17, 824–831. [Google Scholar] [CrossRef]
- Forkman, B.; Boissy, A.; Meunier-Salaün, M.-C.; Canali, E.; Jones, R.B. A Critical Review of Fear Tests Used on Cattle, Pigs, Sheep, Poultry and Horses. Physiol. Behav. 2007, 92, 340–374. [Google Scholar] [CrossRef]
- Murphy, E.; Nordquist, R.E.; van der Staay, F.J. A Review of Behavioural Methods to Study Emotion and Mood in Pigs, Sus Scrofa. Appl. Anim. Behav. Sci. 2014, 159, 9–28. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Definition |
|---|---|
| Nose–nose contact (N) | Touching the nose of another pig with the rooting disc (initiated by different pig roles) |
| Nose–partition contact (N) | Touching the central partition with the rooting disc |
| Proximity (min) | Head within 25 cm of the center partition |
| Exploring (min) | Sniffing, nosing, or rooting the rice hull and the walls of the pen in the rice husk area |
| Escape attempts (N) | Pig jumps in the air or against the wall or door of a compartment |
| Freezing (N) | Standing motionless with whole body and head fixed |
| Equation | Role | R2 | F | df1 | df2 | p-Value |
|---|---|---|---|---|---|---|
| The rewarding group | ||||||
| cubic | TPs | 0.59 | 15.31 | 3 | 32 | <0.01 |
| cubic | NPs | 0.36 | 6.06 | 3 | 32 | <0.01 |
| The punishing group | ||||||
| cubic | TPs | 0.26 | 3.65 | 3 | 32 | 0.02 |
| cubic | NPs | 0.31 | 4.77 | 3 | 32 | <0.01 |
| The paired control group | ||||||
| linear | paired pig | 0.24 | 22.07 | 1 | 70 | <0.01 |
| Equation | R2 | F | df1 | df2 | p-Value |
|---|---|---|---|---|---|
| The rewarding group | |||||
| cubic | 0.257 | 3.685 | 3 | 32 | 0.02 |
| The punishing group | |||||
| quadratic | 0.207 | 4.319 | 2 | 33 | 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yang, X.; Sun, F.; Zhang, Y.; Yao, Y.; Bai, Z.; Yu, J.; Liu, X.; Zhao, Q.; Li, X.; et al. Emotional “Contagion” in Piglets after Sensory Avoidance of Rewarding and Punishing Treatment. Animals 2024, 14, 1110. https://doi.org/10.3390/ani14071110
Zhang Y, Yang X, Sun F, Zhang Y, Yao Y, Bai Z, Yu J, Liu X, Zhao Q, Li X, et al. Emotional “Contagion” in Piglets after Sensory Avoidance of Rewarding and Punishing Treatment. Animals. 2024; 14(7):1110. https://doi.org/10.3390/ani14071110
Chicago/Turabian StyleZhang, Ye, Xuesong Yang, Fang Sun, Yaqian Zhang, Yuhan Yao, Ziyu Bai, Jiaqi Yu, Xiangyu Liu, Qian Zhao, Xiang Li, and et al. 2024. "Emotional “Contagion” in Piglets after Sensory Avoidance of Rewarding and Punishing Treatment" Animals 14, no. 7: 1110. https://doi.org/10.3390/ani14071110