
Effects of Cheliped Amputation on the Personality of Crayfish

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Note
2.2. Experimental Animals and Rearing Conditions
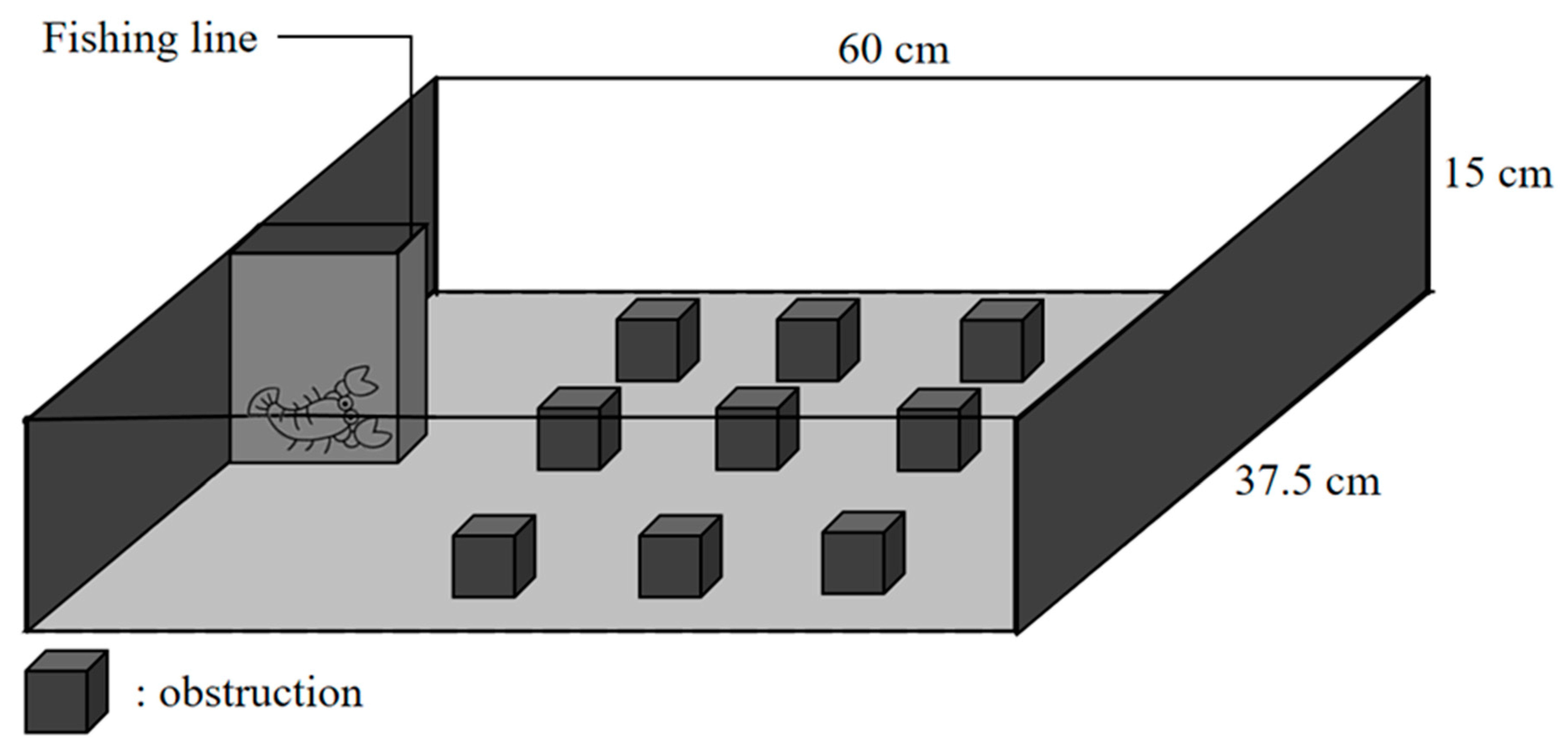
2.3. Exploration Test
2.4. Aggression Test
2.5. Statistical Analyses
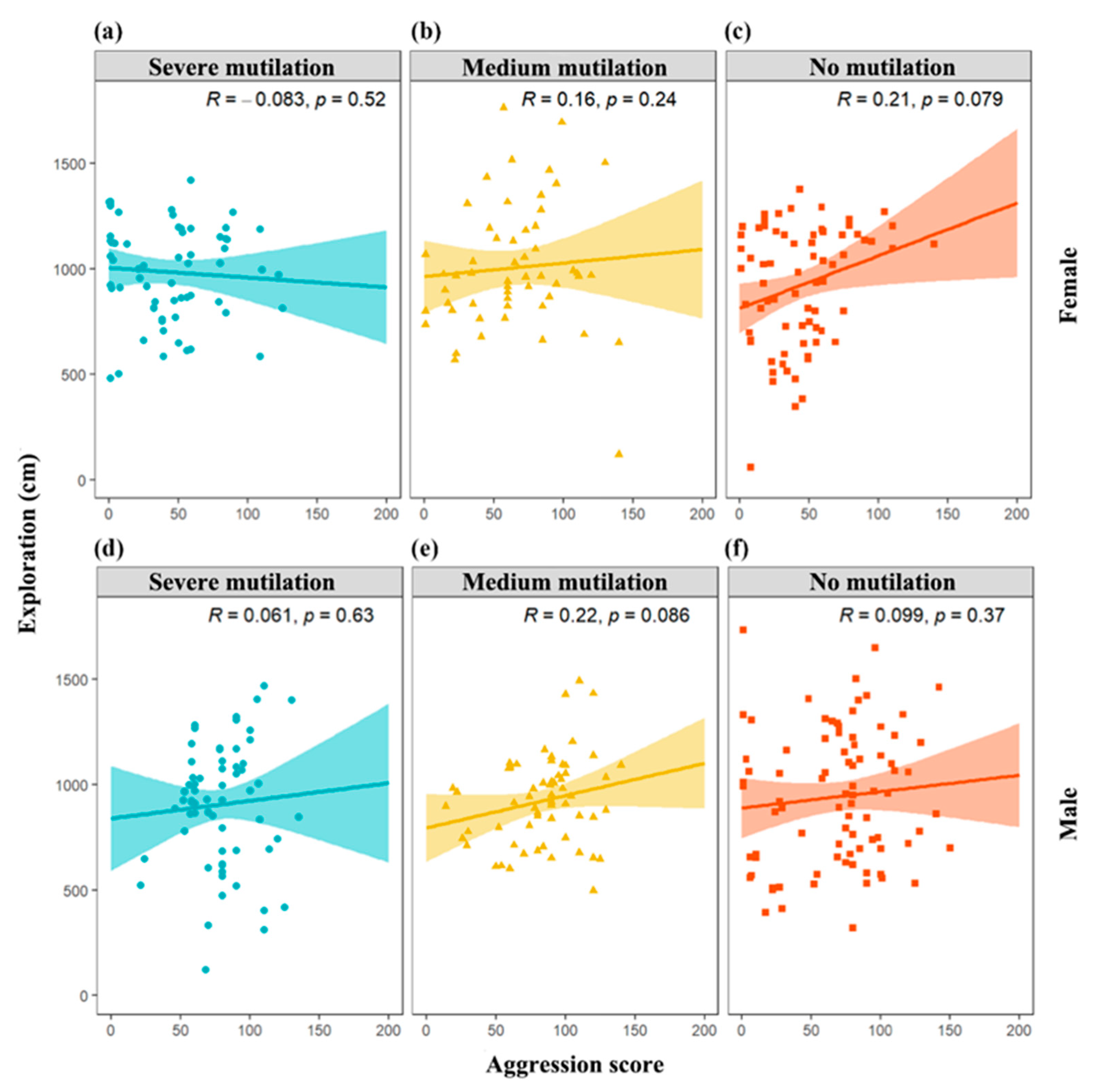
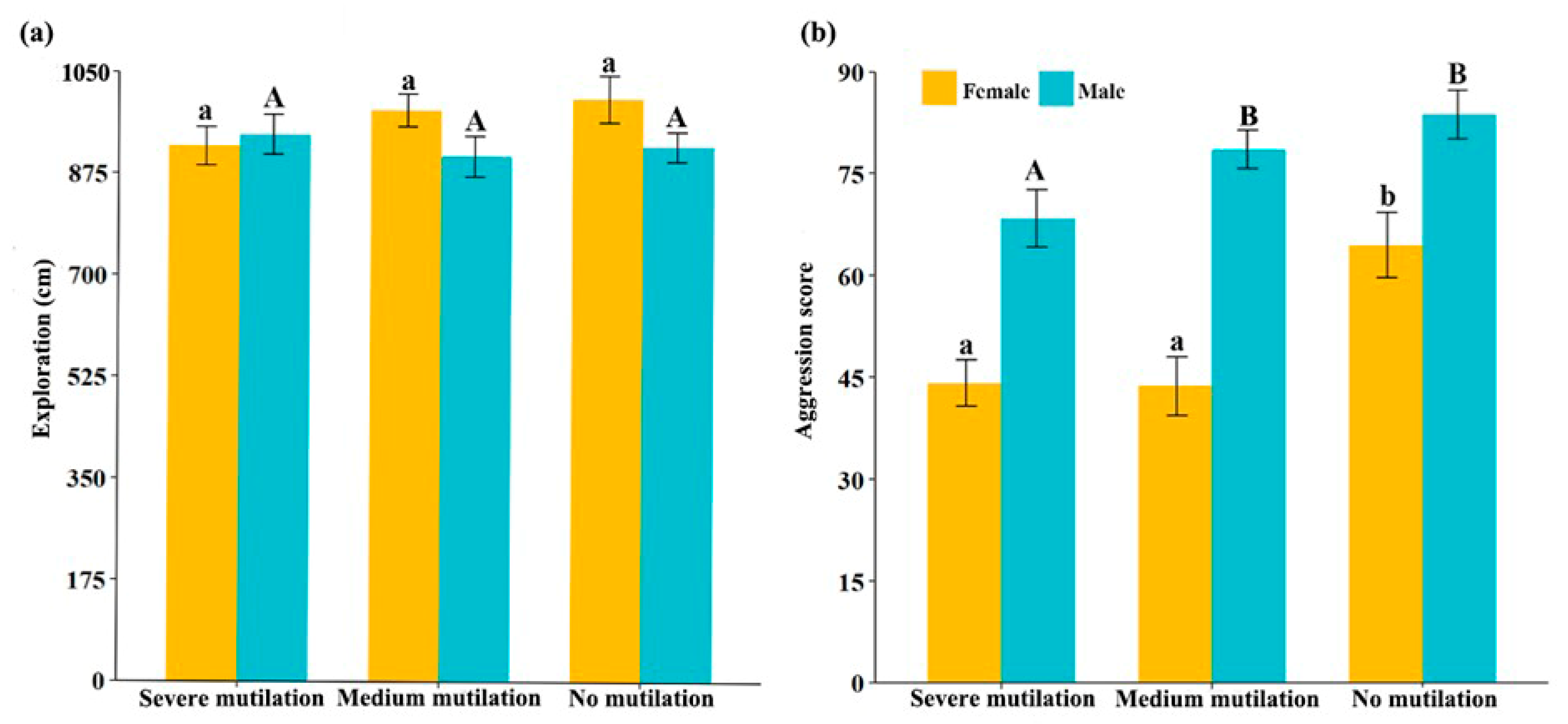
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Briffa, M.; Weiss, A. Animal personality. Curr. Biol. 2010, 20, R912–R914. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Kats, L.B.; Maurer, E.F. Behavioural correlations across situations and the evolution of antipredator behaviour in a sunfish–salamander system. Anim. Behav. 2003, 65, 29–44. [Google Scholar] [CrossRef]
- Cote, J.; Clobert, J. Social personalities influence natal dispersal in a lizard. Proc. Biol. Sci. 2007, 274, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Réale, D.; Reader, S.M.; Sol, D.; McDougall, P.T.; Dingemanse, N.J. Integrating animal temperament within ecology and evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.S. Adaptive individual differences within single populations. Philos. Trans. R. Soc. Lond. B 1998, 353, 199–205. [Google Scholar] [CrossRef]
- Pfennig, D.W.; Wund, M.A.; Snell-Rood, E.C.; Cruickshank, T.; Schlichting, C.D.; Moczek, A.P. Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol. 2010, 25, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Sinn, D.L.; Apiolaza, L.A.; Moltschaniwskyj, N.A. Heritability and fitness-related consequences of squid personality traits. J. Evol. Biol. 2006, 19, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Drent, P.J.; Van Noordwijk, A.J.; Van Oers, K.; de Jong, G.; Kempenaers, B. Contribution of genetics to the study of animal personalities: A review of case studies. Behaviour 2005, 142, 1185–1206. [Google Scholar] [CrossRef]
- Bierbach, D.; Laskowski, K.L.; Wolf, M. Behavioural individuality in clonal fish arises despite near-identical rearing conditions. Nat. Commun. 2017, 8, 15361. [Google Scholar] [CrossRef]
- Polverino, G.; Bierbach, D.; Killen, S.S.; Uusi-Heikkilä, S.; Arlinghaus, R. Body length rather than routine metabolic rate and body condition correlates with activity and risk-taking in juvenile zebrafish Danio rerio. J. Fish Biol. 2016, 89, 2251–2267. [Google Scholar] [CrossRef]
- Xu, W.; Yao, Q.; Zhang, W.; Zhang, F.; Li, H.; Xu, R.; Li, C.; Zhang, B. Environmental complexity during early life shapes average behavior in adulthood. Behav. Ecol. 2021, 32, 105–113. [Google Scholar] [CrossRef]
- Suckow, N.M.; Pollock, H.S.; Hauber, M.E.; Kastner, M.; Savidge, J.A.; Baker, K.; Rogers, H.S. Nest defense, personality, and fitness of a locally endangered island passerine. Ethology 2022, 128, 499–507. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Réale, D. Natural selection and animal personality. Behaviour 2005, 142, 1159–1184. [Google Scholar] [CrossRef]
- Nagel, L.; Schluter, D. Body size, natural selection, and speciation in sticklebacks. Evolution 1998, 52, 209–218. [Google Scholar] [CrossRef]
- Vincent, T.L.; Brown, J.S. Evolutionary Game Theory, Natural Selection, and Darwinian Dynamics; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Giraudeau, M.; Nolan, P.M.; Black, C.E.; Earl, S.R.; Hasegawa, M.; McGraw, K.J. Song characteristics track bill morphology along a gradient of urbanization in house finches (Haemorhous mexicanus). Front. Zool. 2014, 11, 83. [Google Scholar] [CrossRef] [PubMed]
- Sejian, V.; Bhatta, R.; Gaughan, J.B.; Dunshea, F.R.; Lacetera, N. Review: Adaptation of animals to heat stress. Animal 2018, 12, s431–s444. [Google Scholar] [CrossRef] [PubMed]
- Weeks, B.C.; Naeem, S.; Winger, B.M.; Cracraft, J. The relationship between morphology and behavior in mixed-species flocks of island birds. Ecol. Evol. 2020, 10, 10593–10606. [Google Scholar] [CrossRef]
- French, G.C.A.; Stürup, M.; Rizzuto, S.; van Wyk, J.H.; Edwards, D.; Dolan, R.W.; Wintner, S.P.; Towner, A.V.; Hughes, W.O.H. The tooth, the whole tooth and nothing but the tooth: Tooth shape and ontogenetic shift dynamics in the white shark Carcharodon carcharias. J. Fish Biol. 2017, 91, 1032–1047. [Google Scholar] [CrossRef]
- Wirth-Dzieciolowska, E.; Czuminska, K.; Reklewska, B.; Katkiewicz, M. Life time reproductive performance and functional changes in reproductive organs of mice selected divergently for body weight over 90 generations. Anim. Sci. Pap. Rep. 1996, 14, 187–198. [Google Scholar]
- Biro, P.A.; Stamps, J.A. Are animal personality traits linked to life-history productivity? Trends Ecol. Evol. 2008, 23, 361–368. [Google Scholar] [CrossRef]
- Kubec, J.; Kouba, A.; Buřič, M. Communication, behaviour, and decision making in crayfish: A review. Zool. Anz. 2019, 278, 28–37. [Google Scholar] [CrossRef]
- Polverino, G.; Ruberto, T.; Staaks, G.; Mehner, T. Tank size alters mean behaviours and individual rank orders in personality traits of fish depending on their life stage. Anim. Behav. 2016, 115, 127–135. [Google Scholar] [CrossRef]
- Hedrick, A.V.; Kortet, R. Sex differences in the repeatability of boldness over metamorphosis. Behav. Ecol. Sociobiol. 2012, 66, 407–412. [Google Scholar] [CrossRef]
- Langenhof, M.R.; Apperloo, R.; Komdeur, J. Small variations in early-life environment can affect coping behaviour in response to foraging challenge in the three-spined stickleback. PLoS ONE 2016, 11, e0147000. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bailey, J.M.; Oliveri, A.N.; Karbhari, N.; Brooks, R.A.; De La Rocha, A.J.; Janardhan, S.; Levin, E.D. Persistent behavioral effects following early life exposure to retinoic acid or valproic acid in zebrafish. Neurotoxicology 2016, 52, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Mariappan, P.; Balasundaram, C.; Schmitz, B. Decapod crustacean chelipeds: An overview. J. Biosci. 2000, 25, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Clarac, F. Walking patterns of a limb after autotomy in Decapod Crustacea [proceedings]. J. Physiol. 1978, 277, 55P. [Google Scholar]
- Kuris, A.M.; Mager, M. Effect of limb regeneration on size increase at molt of the shore crabs Hemigrapsus oregonensis and Pachygrapsus crassipes. J. Exp. Zool. 1975, 193, 353–360. [Google Scholar] [CrossRef]
- Masunari, N.; Sekiné, K.; Kang, B.J.; Takada, Y.; Hatakeyama, M.; Saigusa, M. Ontogeny of Cheliped Laterality and Mechanisms of Reversal of Handedness in the Durophagous Gazami Crab, Portunus trituberculatus. Biol. Bull. 2020, 238, 25–40. [Google Scholar] [CrossRef]
- Øverli, O.; Winberg, S.; Pottinger, T.G. Behavioral and neuroendocrine correlates of selection for stress responsiveness in rainbow trout—A review. Integr. Comp. Biol. 2005, 45, 463–474. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.M. Insights for behavioral ecology from behavioral syndromes. Adv. Study Behav. 2008, 38, 227–281. [Google Scholar]
- Grant, J.W.A. Whether or not to defend? The influence of resource distribution. Mar. Behav. Physiol. 1993, 23, 137–153. [Google Scholar] [CrossRef]
- Ranta, E.; Lindström, K. Body size and shelter possession in mature signal crayfish, Pacifastacus leniusculus. Ann. Zool. Fenn. 1993, 30, 125–132. [Google Scholar]
- Alderman, D.J. Geographical spread of bacterial and fungal diseases of crustaceans. Rev. Sci. Tech. 1996, 15, 603–632. [Google Scholar] [CrossRef] [PubMed]
- Bovbjerg, R.V. Some factors affecting aggressive behavior in crayfish. Physiol. Zool. 1956, 29, 127–136. [Google Scholar] [CrossRef]
- Xu, Z.H.; Zhao, C.Y.; Zho, X. The relation between carapace color and growth of Procambarus clarkii. Acta Agric. Zhejiangensis 2010, 22, 839–842. [Google Scholar]
- Edenbrow, M.; Croft, D.P. Environmental and genetic effects shape the development of personality traits in the mangrove killifish Kryptolebias marmoratus. Oikos 2013, 122, 667–681. [Google Scholar] [CrossRef]
- Carter, A.J.; Feeney, W.E.; Marshall, H.H.; Cowlishaw, G.; Heinsohn, R. Animal personality: What are behavioural ecologists measuring? Biol. Rev. Camb. Philos. Soc. 2013, 88, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Bell, A.M.; Johnson, J.C.; Ziemba, R.E. Behavioral syndromes: An integrative overview. Q. Rev. Biol. 2004, 79, 241–277. [Google Scholar] [CrossRef]
- Stoffel, M.A.; Nakagawa, S.; Schielzeth, H. rptR: Repeatability estimation and variance decomposition by generalized linear mixed-effects models. Methods Ecol. Evol. 2017, 8, 1639–1644. [Google Scholar] [CrossRef]
- Revelle, W. Psych: Procedures for Psychological, Psychometric, and Personality Research; Package, Version 1.9, Comprehensive R Archive Network (CRAN); Northwestern University: Evanston, IL, USA, 2019; p. 12. [Google Scholar]
- Wolf, M.; Weissing, F.J. Animal personalities: Consequences for ecology and evolution. Trends Ecol. Evol. 2012, 27, 452–461. [Google Scholar] [CrossRef]
- Johnstone, R.A.; Manica, A. Evolution of personality differences in leadership. Proc. Natl. Acad. Sci. USA 2011, 108, 8373–8378. [Google Scholar] [CrossRef]
- Duckworth, R.A. Aggressive behaviour affects selection on morphology by influencing settlement patterns in a passerine bird. Proc. Biol. Sci. 2006, 273, 1789–1795. [Google Scholar] [CrossRef] [PubMed]
- Koski, S.E. How to measure animal personality and why does it matter? Integrating the psychological and biological approaches to animal personality. In From Genes to Animal Behavior: Social Structures, Personalities, Communication by Color; Springer: Tokyo, Japan, 2011; pp. 115–136. [Google Scholar]
- Dall, S.R.X.; Houston, A.I.; Mcnamara, J.M. The behavioural ecology of personality: Consistent individual differences from an adaptive perspective. Ecol. Lett. 2004, 7, 734–739. [Google Scholar] [CrossRef]
- Wingfield, J.C. Control of territorial aggression in a changing environment. Psychoneuroendocrinology 1994, 19, 709–721. [Google Scholar] [CrossRef]
- Garamszegi, L.Z.; Hegyi, G.; Heylen, D.; Ninni, P.; De Lope, F.; Eens, M.; Møller, A.P. The design of complex sexual traits in male barn swallows: Associations between signal attributes. J. Evol. Biol. 2006, 19, 2052–2066. [Google Scholar] [CrossRef] [PubMed]
- Gil, D.; Gahr, M. The honesty of bird song: Multiple constraints for multiple traits. Trends Ecol. Evol. 2002, 17, 133–141. [Google Scholar] [CrossRef]
- Bell, A.M.; Hankison, S.J.; Laskowski, K.L. The repeatability of behaviour: A meta-analysis. Anim. Behav. 2009, 77, 771–783. [Google Scholar] [CrossRef]
- Smith, B.R.; Blumstein, D.T. Fitness consequences of personality: A meta-analysis. Behav. Ecol. 2008, 19, 448–455. [Google Scholar] [CrossRef]
- Harris, M.R.; Siefferman, L. Interspecific competition influences fitness benefits of assortative mating for territorial aggression in eastern bluebirds (Sialia sialis). PLoS ONE 2014, 9, e88668. [Google Scholar] [CrossRef]
- Mackay, J.R.D.; Turner, S.P.; Hyslop, J.; Deag, J.M.; Haskell, M.J. Short-term temperament tests in beef cattle relate to long-term measures of behavior recorded in the home pen1. J. Anim. Sci. 2013, 91, 4917–4924. [Google Scholar] [CrossRef]
- Kortet, R.; Vainikka, A.; Janhunen, M.; Piironen, J.; Hyvärinen, P. Behavioral variation shows heritability in juvenile brown trout Salmo trutta. Behav. Ecol. Sociobiol. 2014, 68, 927–934. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef]
- Kelleher, S.R.; Silla, A.J.; Byrne, P.G. Animal personality and behavioral syndromes in amphibians: A review of the evidence, experimental approaches, and implications for conservation. Behav. Ecol. Sociobiol. 2018, 72, 79. [Google Scholar] [CrossRef]
- Seward, J.P. Aggressive behavior in the rat. I. General characteristics; age and sex differences. J. Comp. Psychol. 1945, 38, 175–197. [Google Scholar] [CrossRef]
- Garamszegi, L.Z.; Markó, G.; Herczeg, G. A meta-analysis of correlated behaviors with implications for behavioral syndromes: Relationships between particular behavioral traits. Behav. Ecol. 2013, 24, 1068–1080. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Wright, J.; Kazem, A.J.; Thomas, D.K.; Hickling, R.; Dawnay, N. Behavioural syndromes differ predictably between 12 populations of three-spined stickleback. J. Anim. Ecol. 2007, 76, 1128–1138. [Google Scholar] [CrossRef] [PubMed]
- Minderman, J.; Reid, J.M.; Evans, P.G.H.; Whittingham, M.J. Personality traits in wild starlings: Exploration behavior and environmental sensitivity. Behav. Ecol. 2009, 20, 830–837. [Google Scholar] [CrossRef]
- Mason, G.J. Stereotypies: A critical review. Anim. Behav. 1991, 41, 1015–1037. [Google Scholar] [CrossRef]
- Van Oers, K.; Drent, P.J.; De Jong, G.; van Noordwijk, A.J. Additive and nonadditive genetic variation in avian personality traits. Heredity 2004, 93, 496–503. [Google Scholar] [CrossRef][Green Version]
- Koehl, M.A.R. When does morphology matter? Annu. Rev. Ecol. Syst. 1996, 27, 501–542. [Google Scholar] [CrossRef]
- Huxley, T.H. The Crayfish: An Introduction to the Study of Zoology; C. Kegan Paul: London, UK, 1880. [Google Scholar]
- Gagneux, P.; Archer, J. The Behavioural Biology of Aggression; Cambridge University Press: Cambridge, UK, 1988; Volume 257, pp. 515–516. [Google Scholar]
- Colléter, M.; Brown, C. Personality traits predict hierarchy rank in male rainbowfish social groups. Anim. Behav. 2011, 81, 1231–1237. [Google Scholar] [CrossRef]
- Figler, M.H.; Holly, M.C.; Grant, S. Blank Shelter competition in juvenile red swamp crayfish (Procambarus clarkii): The influences of sex differences, relative size, and prior residence. Aquaculture 1999, 178, 63–75. [Google Scholar] [CrossRef]
- Dall, S.R. Behavioural biology: Fortune favours bold and shy personalities. Curr. Biol. 2004, 14, R470–R472. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Vanderschuren, L.J.; Niesink, R.J.; Spruijt, B.M.; Van Ree, J.M. Influence of environmental factors on social play behavior of juvenile rats. Physiol. Behav. 1995, 58, 119–123. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Winkler, H.; Leisler, B. The significance of ecological factors for exploration and neophobia in parrots. Ethology 2002, 108, 249–272. [Google Scholar] [CrossRef]
- Berrill, M.; Arsenault, M. The breeding behaviour of a northern temperate orconectid crayfish, Orconectes rusticus. Anim. Behav. 1984, 32, 333–339. [Google Scholar] [CrossRef]
- Moore, P.A. Agonistic behavior in freshwater crayfish. Evol. Ecol. Soc. Sex. Syst. 2007, 90–114. [Google Scholar] [CrossRef]
- Goessmann, C.; Hemelrijk, C.; Huber, R. The formation and maintenance of crayfish hierarchies: Behavioral and self-structuring properties. Behav. Ecol. Sociobiol. 2000, 48, 418–428. [Google Scholar] [CrossRef]
- Figler, M.H.; Twum, M.; Finkelstein, J.E.; Peeke, H.V.S. Maternal aggression in red swamp crayfish (Procambarus clarkii, Girard): The relation between reproductive status and outcome of aggressive encounters with male and female conspecifics. Behaviour 1995, 132, 107–125. [Google Scholar] [CrossRef]
- Bergman, D.A.; Moore, P.A. Field observations of intraspecific agonistic behavior of two crayfish species, Orconectes rusticus and Orconectes virilis, in different habitats. Biol. Bull. 2003, 205, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Herberholz, J.; Mccurdy, C.; Edwards, D.H. Direct benefits of social dominance in juvenile crayfish. Biol. Bull. 2007, 213, 21–27. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sex | Mutilation Degree | Sample Size | Exploration (cm) | Aggression Score |
|---|---|---|---|---|
| Female | Severe mutilation | 24 | 922.2 (±57.6) | 44.2 (±6.0) |
| Medium mutilation | 21 | 984.6 (±49.1) | 43.7 (±7.5) | |
| No mutilation | 18 | 1004.4 (±69.8) | 64.5 (±8.3) | |
| Male | Severe mutilation | 28 | 941.9 (±59.6) | 68.4 (±7.2) |
| Medium mutilation | 22 | 904.7 (±60.3) | 78.6 (±4.9) | |
| No mutilation | 21 | 922.6 (±44.7) | 83.8 (±6.2) |
| Sex | Repeatability | Exploration (cm) | ||
|---|---|---|---|---|
| Severe Mutilation | Medium Mutilation | No Mutilation | ||
| Female | R | 0.191 | 0.18 | 0.09 |
| Standard error | 0.129 | 0.13 | 0.11 | |
| 95% confidence interval | 0, 0.47 | 0, 0.45 | 0, 0.40 | |
| p-value | 0.06 | 0.66 | 0.207 | |
| Male | R | 0.12 | 0.38 | 0.40 |
| Standard error | 0.11 | 0.14 | 0.15 | |
| 95% confidence interval | 0, 0.35 | 0, 0.59 | 0.04, 0.61 | |
| p-value | 0.13 | 0.002 | 0.001 | |
| Sex | Repeatability | Aggression Score | ||
|---|---|---|---|---|
| Severe Mutilation | Medium Mutilation | No Mutilation | ||
| Female | R | 0.50 | 0.79 | 0.57 |
| Standard error | 0.15 | 0.12 | 0.17 | |
| 95% confidence interval | 0.19, 0.73 | 0.41, 0.88 | 0.23, 0.83 | |
| p-value | <0.001 | <0.001 | <0.001 | |
| Male | R | 0.56 | 0.57 | 0.96 |
| Standard error | 0.18 | 0.15 | 0.02 | |
| 95% confidence interval | 0.27, 0.89 | 0.22, 0.78 | 0.91, 0.98 | |
| p-value | <0.001 | <0.001 | <0.001 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.; Su, L.; Si, M.; Wang, G.; Li, C. Effects of Cheliped Amputation on the Personality of Crayfish. Animals 2024, 14, 1132. https://doi.org/10.3390/ani14071132
Lu L, Su L, Si M, Wang G, Li C. Effects of Cheliped Amputation on the Personality of Crayfish. Animals. 2024; 14(7):1132. https://doi.org/10.3390/ani14071132
Chicago/Turabian StyleLu, Leiyu, Li Su, Mengdi Si, Guangyao Wang, and Chunlin Li. 2024. "Effects of Cheliped Amputation on the Personality of Crayfish" Animals 14, no. 7: 1132. https://doi.org/10.3390/ani14071132
APA StyleLu, L., Su, L., Si, M., Wang, G., & Li, C. (2024). Effects of Cheliped Amputation on the Personality of Crayfish. Animals, 14(7), 1132. https://doi.org/10.3390/ani14071132