Simple Summary
Procambarus clarkii is typically viewed as a predator rather than prey, due to its strong anti-predatory capabilities. However, during its juvenile stage, it remains vulnerable to predators. Nonetheless, complex outdoor habitats can offer some protection. By examining the effects of three different types of shelters with various coverages and combinations on the predation rhythm, behavior, and efficiency of Silurus asotus on juvenile Procambarus clarkii, we found that the presence and complexity of shelters in the environment have a limited impact on the predatory activities of Silurus asotus. Among the shelter types, PVC pipes offered the best protection, followed by stone, with water grass providing the least protection. Furthermore, the protective effect of shelters increased with coverage rate. Therefore, we suggest that the type and abundance of shelters significantly influence the predation rhythm and behavior of Silurus asotus on Procambarus clarkii in outdoor environments.
Abstract
Procambarus clarkii is adept at using natural shelters and caves to evade attacks from predators. However, the concealment abilities and mechanisms of P. clarkii for different types of shelters under predation pressure have not yet been reported. In this study, laboratory experiments were carried out to determine the effects of different coverages (25%, 50%, and 75%) and different combinations (I–VII) of three types of shelters (PVC pipes, water grass, and stone) on the predation rhythm, behavior, and abilities of Silurus asotus on P. clarkii. The results indicated that the predation of S. asotus on P. clarkii exhibited significant rhythmicity under shelter conditions, excluding PVC pipes, 75% stone, and combination VI. Among the three types of shelters, PVC pipes provided the strongest concealment, followed by stone and water grass. With the increase in shelter coverage, the anti-predation ability of P. clarkii continued to increase, and the optimal shade rate for water grass was 50%. In the different shelter combinations, the environmental complexity had little effect on the predation activity of S. asotus on P. clarkii. These findings demonstrated that the type and abundance of shelters in the wild environment can affect the predation rhythm and activities of S. asotus on P. clarkii.
1. Introduction
Procambarus clarkii is a typical cave-dwelling animal with social behavior [1], known for its aggressiveness, burrowing habits, and robust anti-predation ability [2]. In the late 1920s, P. clarkii was introduced to China from Japan, and it has since established a wide distribution in the Yangtze River Basin, occupying an ecological niche in Poyang Lake [3]. From 2009 to 2013,the production of P. clarkii in Poyang Lake represented over 50% of the total shrimp fishing production, averaging 2.5 × 104 tons [4]. However, following the ban on fishing in the Yangtze River in 2021, P. clarkii in Poyang Lake exhibited a trend towards smaller individuals and a substantial decline in resources. For example, the results of a 2021~2022 survey indicated that the presence of P. clarkii was detected at only 5 of the 14 sampling sites, and the number of P. clarkii caught was significantly lower than before the closure [5]. In nature, the predation activities of numerous predators, including waterfowl, water snakes, and some fierce carnivorous fish (Anguilla anguilla, Silurus asotus, Perca fluviatilis, etc.) may contribute to the decline in P. clarkii resources [6,7,8]. The interactions between prey size and predator species can influence the feeding preferences of the predator [9]. For example, predators exhibit a clear preference for the size of Penaeus orientalis [10] and Cherax quadricarinatus [11]. For P. clarkii, the small larvae may face a higher risk of predation.
The natural shelters in the environment offer protection for prey from the threat of predation. For P. clarkii, various effective shelter types include substrates, caves, large plants [12], and even shadows [13]. The specifications of shelters are a crucial factor affecting their protective capacity, with larger shelters typically providing better protection for P. clarkii and influencing their efficiency in responding to predators [14]. The effectiveness of shelters is proportional to their density and complexity [15,16]. Shelters can reduce the frequency and duration of interspecies competition and the risk of interspecies predation, significantly improving the survival rate of P. clarkii [1,13]. Shelters are more effective in reducing the aggression of Cambarus than food [17]. P. clarkii exhibits an obvious preference for different types of shelters. Regarding shelter type, P. clarkii preferred narrow, dark, and breathable shelters, showing a clear selective preference for cave-type shelters [18]. In terms of shelter materials, P. clarkii preferred bamboo-type shelters [19].
However, shelters cannot consistently yield a positive effect for P. clarkii, whether in the natural environment or aquaculture process. In the environments with shelters, the different developmental stages of P. clarkii will compete for shelters, leading to the larvae of P. clarkii being confined to areas with sparse shelters, increasing the risk of predation [20]. Simultaneously, in order to reduce the probability of interspecies fighting and predation, P. clarkii feed only to satisfy their own minimum energy expenditure, resulting in diminished growth rates [21]. Additionally, some shelters like straw, mud, and dead leaves may decrease the hatching rate of fertilized oosperm [22]. Currently, numerous studies have focused on the effects of shelters on the growth of P. clarkii, but there is limited research on the impact of shelters on its other life activities. Poyang Lake is a typical representative of composite river and lake ecology, with a complex habitat and numerous predators. Whether shelters will affect the anti-predation activity of P. clarkii and the mechanism of action remain unknown.
In this study, P. clarkii were used as the prey and Silurus asotus were selected as the predator. Three types of shelters (PVC pipes, water grass, and stone) were used to simulate diverse habitat conditions in Poyang Lake. Moreover, the different coverages and combinations of three shelters were set up to analyze the feeding rhythm, anti-predation behavior, and activity of P. clarkii. These results will reveal the influence mechanism of different habitats on the anti-predation activity of P. clarkii and provide a scientific reference for the resource assessment and sustainable development of P. clarkii in Poyang Lake.
2. Materials and Methods
2.1. Source and Maintenance of Experimental Animals
The morphological indices of P. clarkii and S. asotus were shown in Table 1 (with mean ± standard error), which were taken from the aquaculture base of the Jiangxi Fisheries Research Institute (Nanchang, Jiangxi). Before the test, P. clarkii and S. asotus were separately maintained in 84 indoor aquaria (length 1 m × width 0.6 m × height 0.55 m, water depth 0.15 m, each equipped with a recirculating system) for two weeks. During the acclimation, water subjected to continuous aeration for more than 48 h was provided, and an appropriate amount of sediment was added to the water to simulate the water transparency of Poyang Lake during the dry water period. The water temperature was maintained at 23 ± 0.5 °C, with a photoperiod of 12 h dark:12 h light, pH of 7.5 ± 0.2, and dissolved oxygen ≥ 6 mg/L. Commercial crayfish pellets (32% crude protein, 4% crude lipid, 8% crude fiber, 3% calcium, 1% phosphorus, 1.5% lysine, 15% ash and 14% moisture) were used to feed P. clarkii every three days, and P. clarkii of the above size were employed to feed the S. asotus to saturation once every other day.

Table 1.
Morphological indices of two experimental aquatic animals.
2.2. Experimental Design and Procedure
Three levels of shelter coverage (25%, 50%, 75%) and seven different combinations of three types of shelters (PVC pipes: φ 0.028 m, length 0.1 m; water grass: length 0.1 m; stone: length 0.08 m, width 0.06 m) with 50% coverage were set up (Table 2), including 14 groups with three replicates each, and the control group without shelters. The experiment lasted for 24 h. All experimental conditions were consistent with the acclimation and the photoperiod consisted of 12 h light and 12 h dark. A transparent perforated acrylic plate separating areas A and B of the blue test device (Figure 1) was used, and two cameras were mounted directly above and at the bottom. A small flushing device (length 0.4 m × width 0.75 m × height 0.8 m) was installed in front to simulate the water flow rate.
Table 2.
Combination of three different types of shelters.
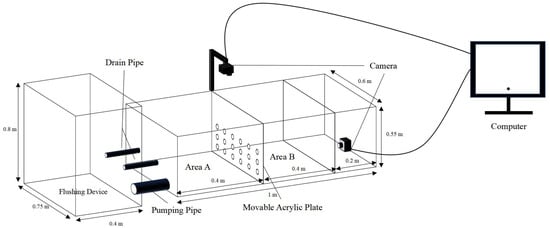
Figure 1.
Predator–prey test device.
The 16 P. clarkii and one S. asotus subjected to 48 h starvation were randomly placed in areas A–B of the test device for two hours before the experiment. After two hours, the plate was withdrawn and the video recording system was activated to document the experiment. The remaining number of P. clarkii in the test device was recorded at 12 h intervals. After 24 h, the number of prey of P. clarkii was counted. The number of attacks and predation time of S. asotus were recorded and analyzed for the predation behavior of S. asotus and the anti-predation behavior of P. clarkii in the recorded video.
2.3. Data Statistical Analysis
The behavior of S. asotus when approaching, attacking, and handling P. clarkii was considered as a complete predation event. The number of prey (NOP), relative predation rate (RPR) of P. clarkii, and the success rate of foraging (SROF) of S. asotus were calculated as follows:
where, N0: Initial number of P. clarkii, Ni: Remaining number of P. clarkii, D: Number of predators, NOA: Number of attacks.
The results were expressed as mean ± standard error (SE). IBM SPSS Statistics 27.0 was used to conduct one-way ANOVA and multiple comparisons to analyze differences between groups, p < 0.05 indicates a significant difference. Results were plotted with Graphpad Prism 8.
3. Results
3.1. Predation Rhythm
The predation results of daytime and night (12 h:12 h) indicated that the RPR for the other experimental groups of S. asotus on P. clarkii was significantly higher at night (p < 0.05) with the exception of 25% and 75% PVC pipes, 75% stone, and combination VI (Table 3). Similarly, except for 75% PVC pipes, the NOA of S. asotus was significantly higher at night (p < 0.05) (Table 3).
Table 3.
Predation rhythm of S. asotus on P. clarkii under different shelter conditions.
3.2. Predation and Anti-Predation Behavior
Fish aggression was an adaptive behavior exhibited by individual animals of the same or different species to compete for space or food resources, which was an innate and prominent characteristic of fish [23]. Predation strategies and behaviors of S. asotus were found to be consistent across species and coverage of shelters during the course of the experiment. Sample images from videos, depicting the predation behavior of S. asotus on P. clarkii, are shown in figure. 2. Based on the analysis of the video recordings, the predation process of S. asotus on P. clarkii was divided into three stages, including the search, attack, and prey handling. Stage I was the search stage, in which S. asotus persisted in searching for prey along the wall of the test device and most P. clarkii remained stationary on the wall or in a corner during the attack latency. Stage II was the attack stage. Upon contact with the P. clarkii, the S. asotus immediately accelerated and attacked the body of P. clarkii (Figure 2A,B). Simultaneously, the P. clarkii rapidly raised their pedes chela and swung their tails to escape along the wall or toward the opposite direction. It remained stationary on another safe corner or wall if successful, and S. asotus would continue to swim along the tank wall in search of the next prey. Stage III was the prey handling stage. After capturing the prey, S. sotus gradually swallowed it for chewing upon capturing the prey, and consistently pushed the prey against the wall and shook its body rhythmically until it completely devoured its prey during the swallowing process (Figure 2C,D). After finishing its meal, S. asotus left along the wall of device.

Figure 2.
Predatory process of S. asotus on P. clarkii. The letters (A–D) indicated different serial numbers, and the arrows pointed to the momentary behavior of S. asotus preying on P. clarkii.
3.3. Effects of Different Coverages of Shelter on the Anti-Predation of P. clarkii
Under the effect of three types of shelters with varying coverage rates, except for 25% stone, the RPR of P. clarkii in the remaining experimental groups was significantly lower than that in the control group (p < 0.05) (Figure 3). The NOA of S. asotus on P. clarkii in the remaining experimental groups was significantly lower than that in the control group with the exception of 25% PVC pipes and 75% water grass and 25% stone (p < 0.05) (Figure 4). And, apart from the 25% PVC pipes, 50% stone, and 75% water grass, the SROF of S. asotus on P. clarkii in the remaining experimental groups was significantly lower than that in the control group (p < 0.05) (Figure 5)
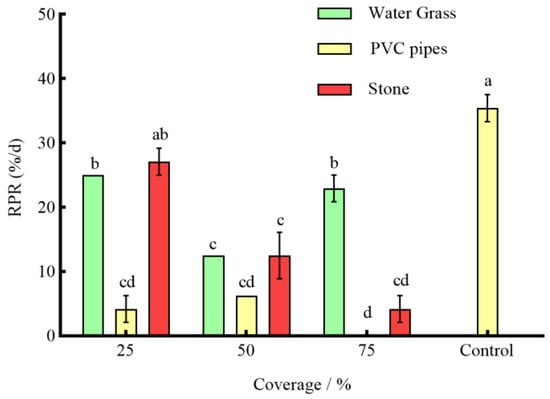
Figure 3.
The RPR of P. clarkii under different coverage rates. The difference in letters indicated that there were significant differences between groups (p < 0.05), and the same applies below.
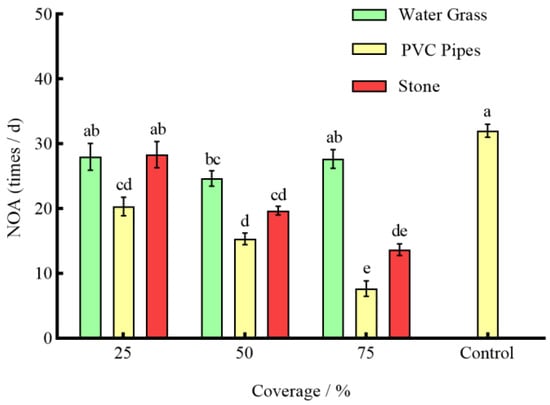
Figure 4.
The NOA of S. asotus on P. clarkii under different coverage rates.
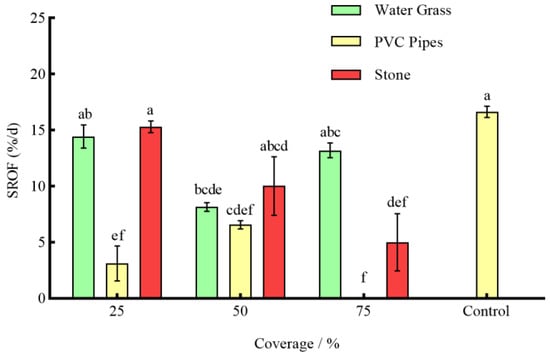
Figure 5.
The SROF of S. asotus on P. clarkii under different coverage rates.
In the presence of water grass sheltering, the RPR of P. clarkii with 50% coverage was significantly lower than that with 25% and 75% coverage (p = 0.0008; p = 0.0021) (Figure 3). The NOA of S. asotus on P. clarkii showed no significant difference under three coverage rates (p > 0.05) (Figure 4). The SROF of S. asotus on P. clarkii with 50% coverage was significantly lower than that with 25% coverage (p = 0.0143) (Figure 5). In the presence of PVC pipe sheltering, there was no significant difference in the RPR and SROF under three coverage rates (p > 0.05) (Figure 3 and Figure 5). The NOA with 75% coverage was significantly lower than that with 25% and 50% coverage (p < 0.0001; p = 0.0201) (Figure 4). In the presence of stone sheltering, the RPR and NOA of 50% and 75% coverage were significantly lower than that of 25% coverage (p < 0.05) (Figure 3 and Figure 4) and the SROF with 75% coverage was significantly lower than that with 25% coverage (p = 0.0009) (Figure 5).
The RPR, NOA, and SROF of S. asotus on P. clarkii with PVC pipes were consistently lower than those of water grass and stone at an equal coverage rate. Specifically, at the coverage rate of 25%, the RPR, NOA, and SROF of PVC pipes were significantly lower than those of water grass and stone (p < 0.05) (Figure 3, Figure 4 and Figure 5). The NOA of PVC pipes was significantly lower than that of water grass at the coverage rate of 50% (p = 0.0031) and there was no significant difference in the SROF of three types of shelters (p > 0.05) (Figure 5). At the coverage rate of 75%, the RPR, NOA, and SROF of PVC pipes and stone were significantly lower than those of water grass (p < 0.05) (Figure 3, Figure 4 and Figure 5).
As seen above, 25%, 50% and 75%, respectively, represent the coverage of the three kinds of shelters. The control is the control group, the same applies below.
3.4. Effects of Different Combinations of Shelter on the Predation of S. asotus on P. clarkii
In the combinations of different shelters with a coverage rate of 50%, the RPR of seven combinations was significantly lower than that of the control group (p < 0.05) (Figure 6). Except for combination I, the NOA of S. asotus on P. clarkii in the remaining combinations was significantly lower than that in the control group (p < 0.05) (Figure 7). With the exception of combination V, the SROF of S. asotus on P. clarkii in the remaining combinations was significantly lower than that in the control group (p < 0.05) (Figure 6).
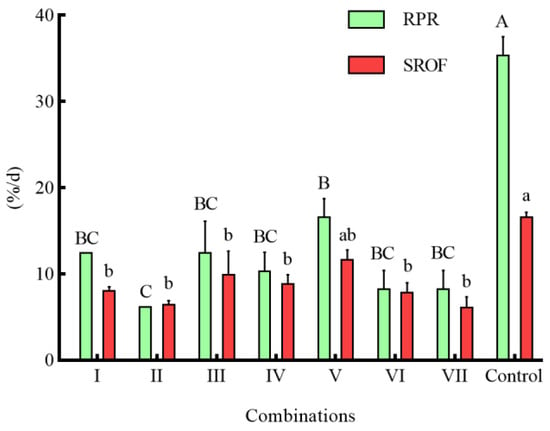
Figure 6.
The RPR of P. clarkii and SROF of S. asotus on P. clarkii under different combinations of shelters. The difference in case letters indicated that there were significant differences between groups (p < 0.05).
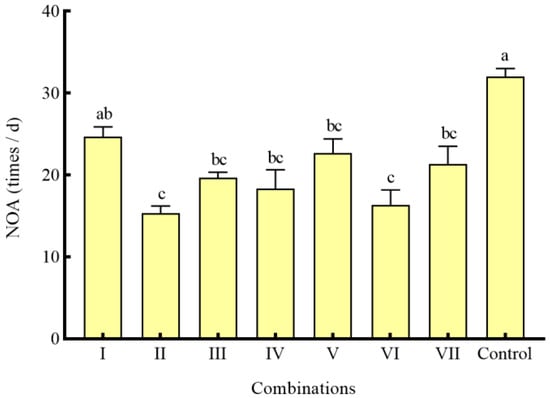
Figure 7.
The NOA of P. clarkii under different combinations of shelters. The difference in case letters indicated that there were significant differences between groups (p < 0.05).
Within three single shelter combinations (I, II, III), there was no significant difference in the RPR and SROF of three combinations (p > 0.05) (Figure 6), The NOA of combination II was significantly lower than that of combination I (p = 0.0139) (Figure 7) and there was no significant difference in the RPR NOA and SROF among combinations IV–VI (p > 0.05) (Figure 6 and Figure 7). The RPR, NOA, and SROF showed no significant differences with the increasing type of shelters among the four groups (I, IV, V, VII) containing water grass. Similarly, no notable distinctions were observed among the intergroups that included stone (II, IV, VI, VII) and PVC pipes (III, V, VI, VI) (Figure 6 and Figure 7).
4. Discussion
4.1. Predation Rhythm of S. asotus under Different Shelter Conditions
The photoperiod can directly or indirectly regulate the diurnal cycle of dissolved oxygen and temperature of water, and in turn, affects the feeding activity of fish [24,25]. For example, the feeding frequency and amount of Cyprinus carpio and Abalone can be significantly affected by the photoperiod [26,27]. In this study, the total predation activity of S. asotus on P. clarkii in all shelter conditions except 25%, 75% PVC tube, 75% stone, and combination VI showed a clear predation rhythm, consistent with the negatively correlated feeding intensity of Channa argus with illumination [28]. S. asotus, which mainly relies on olfaction and mechanical touch for predation, is a typical demersal fish with negative phototropism for light [29]. Nevertheless, the photoperiod can also assist some fishes that rely on vision for predation in better locating their prey. The reason for the differences in daytime and night feeding rhythms among the five groups mentioned above may be related to the type and coverage rates of shelters. The P. clarkii has a sufficient number of shelters to avoid the potential attack threat of the predator after addressing the predation threat with a high coverage rate of PVC pipes, reducing interactions with S. asotus, consequently affecting the predation rhythm of the S. asotus.
4.2. Effects of Different Coverage of Shelter on the Predation of S. asotus on P. clarkii
The change in the habitat environment can change the predator’s ability to search and process prey [30,31] and the structure of the hidden area of prey [32]. In this study, the RPR, NOA, and SROF of S. asotus on P. clarkii were reduced to varying degrees by the presence of the three shelters. Among them, the inhibition ability of PVC pipes was the strongest, followed by stone and water grass, which may be related to the concealment ability of shelters and the habitat habits of P. clarkii [33].
P. clarkii cling to water grass, using it to hide and escape from predators when threatened by predators [34]. It was found that both the RPR and SROF of S. asotus on P. clarkii exhibit a trend of initially decreasing and then increasing with the increase in the coverage rate of shelter in this study. This may be attributed to the insufficient coverage rate of water grass, so that P. clarkii had no enough shelters for concealment. Some P. clarkii can only stay on the device wall or corner and expose to S. asotus, increasing the predation risk by S. asotus. As the coverage rate increased to 50%, there are enough shelters for P. clarkii to avoid the pursuit of S. asotus, resulting in a decrease in the RPR of P. clarkii and the NOA of S. asotus. The increase in the biomass of large plants affects the ability of visual predators to detect prey [35]. As the coverage rate increased to 75%, excess water grass contacted with S. asotus—which relies on olfactory and mechanical touch for predation—increased the NOA, and provided a good hiding effect for S. asotus [36], which reduced its exposure opportunities to P. clarkii, resulting in a decrease in the anti-predation awareness of P. clarkii and an increase in its predation risk. Ostrand et al. proposed that prey species was the key factor affecting the predation efficiency of predators, followed by vegetation density [37]. This may be the reason why the results of this study are inconsistent with Gotceitas’ conclusion that the increase in vegetation density will significantly reduce the predation efficiency of predator [38].
As a typical burrowing animal, P. clarkii is timid and prefers solitude [39]. It avoids predation by seeking shelter in the cave after feeling the threat of predation [2]. Similarly, Palaemonetes pugio can use coarse wood chips as a cover when facing predatory fish [40]. In this study, the NOA and SROF of S. asotus on P. clarkii decreased with the increasing coverage rates of PVC pipes. This decline was attributed to potential conflicts among P. clarkii individuals in the cave with a low coverage rate of PVC pipes, where stronger individuals displaced weaker counterparts from shelters, creating opportunities for S. asotus to attack P. clarkii. As the coverage rate of PVC pipes increased, enough PVC pipes provided sufficient shelter for P. clarkii, which reduced its exposure opportunity to S. asotus. This is consistent with the research conclusions of Gotceitas et al. [38].
The principle of the stone is similar to that of the PVC tube. We found that the P. clarkii will lie under the stone and in crevices to avoid the predator’s attack when the type of shelter is stone, This is consistent with the conclusion that Homarus americanus can use stone crevices and caves to evade predators [41]. In this study, both the RPR, NOA, and SROF of S. asotus on P. clarkii continued to decline with the increase in the coverage rate of stone. This was due to the fact that P. clarkii did not have enough stones to avoid predators when the coverage rate of stone was low, and this phenomenon improved with increasing stone coverage. The existence of stones will also increase the difficulty of S. asotus predation on P. clarkii [42].
4.3. Effects of Different Combinations of Shelter on the Predation of S. asotus on P. clarkii
In the wild environment, habitat complexity diminishes the predator’s predation efficiency by inhibiting the predator’s mobility and providing shelter for prey [43,44], The distribution and quality of shelters affects the strength, duration, and outcome of animal interactions [45,46]. However, some studies have indicated that higher environmental complexity is more conducive to ambushing the predator’s predation activities [36]. In this study, as the complexity of the shelter increased, no significant difference was observed between the experimental groups. This implies that the complexity of the shelters had little effect on the predation activity of S. asotus. The outcome is related to the unique biological characteristics of S. asotus, which preferred to use the complex environment to ambush prey. Simultaneously, the prey size, anti-predation ability, and defense structure affect the predation activity of S. asotus on P. clarkii [47,48].
4.4. Effects of Shelter on the Resources of P. clarkii
Hydrological changes in Poyang Lake significantly impact the distribution and diversity of wetland vegetation [49]. Poyang Lake experiences a dry season from November to March annually [50]. During this period, wetland vegetation is barren, constituting a critical phase for the growth and development of P. clarkii larvae [51], while other bait organisms such as Squaliobarbus curriculus and Pseudorasbora parva are larger [52]. P. clarkii larvae encounters heightened predation challenges in response to the predatory attacks of wintering ferocious fish during this period [53]. Its weak anti-predation and burrowing ability renders it incapable of evading attacks from ferocious fish. The vegetation in Poyang Lake from April to October is abundant and aggressive fish continue to pose a threat to P. clarkii after emerging from hibernation. Nevertheless, the rich vegetation provides natural shelters for the mature P. clarkii with a strong anti-predation ability, allowing it to escape from the fierce fish. At the peak of spawning and juvenile growth of many bait fish at this stage, aggressive fishes preferentially prey on juveniles with low anti-predator capacity when multiple bait organisms coexist in Poyang Lake [54], which reduces the risk of P. clarkii predation.
In the wild environment, the seasonal burrowing behavior of P. clarkii is primarily concentrated from May to October [55]. Sufficient caves can provide shelter for P. clarkii, mitigating the risk of predator attacks during this stage. On the contrary, in November to April, little caves can be used by P. clarkii to evade predator attacks, greatly increasing its risk of predation. Furthermore, the burrowing ability of P. clarkii positively correlates with its growth [56]. P. clarkii larvae exhibit poor burrowing ability. Faced with a predation threat, low-quality caves fail to offer effective shelter, leaving them vulnerable to predator attacks.
4.5. Effects of Environmental Conditions on the Predatory Activities of Fishes
The effect of water transparency on the predation activity of fish is directly related to the predation mode of fish [57,58] such as Lateolabrax japonicus and other fish relying on vision to locate the prey. Lower water transparency will seriously affect its normal predation activity and reduce its predation efficiency [59]. The water transparency of Poyang Lake changes periodically with seasons, i.e., the water transparency is higher during the abundant water period and lower during the dry water period [49]. In this study, sediment was added to simulate the water transparency of Poyang Lake during the dry water period, which resulted in the experimental results only reflecting the predation mechanism of S. asotus on P. clarkii during the dry water period.
Both S. asotus and P. clarkii prefer to live in stone crevices and caves in the field, and engage in life activities in the evening and at night [60,61]. This overlap in habitat and active periods could be a contributing factor to S. asotus predating on P. clarkii. Meanwhile, many fierce fishes in Poyang Lake migrate to the shores to spawn during the spring, meaning that the P. clarkii larvae inhabiting the shallow water and mudflats may face a greater threat of predation [62].
The size of the oral fissure is a crucial factor limiting the feeding preferences of predators [63], the smaller size of P. clarkii larvae expands the spectrum of potential predators. According to the Optimal Foraging Theory [64], predators expend significantly more energy attacking adult P. clarkii than they gain in return. Conversely, preying on P. clarkii larvae optimizes the predator’s energy acquisition [54]. This increased efficiency may contribute to the heightened predation risk faced by P. clarkii larvae.
5. Conclusions
Generally, this study revealed that different shelter combinations and coverage rates of three types of shelters (PVC pipes, water grass and stone) exerted varying inhibitory effects on the predation activity of S. asotus on P. clarkii. Among the three types of shelter, PVC pipes exhibit the strongest sheltering ability, followed by stone and water grass. In different shelter combinations, environmental complexity had little impact on the predation activity of S. asotus on P. clarkii. Due to the size limitations of the experimental device and single species of prey, it is impossible to fully simulate the natural habitat, which may lead to limitations in the experimental results and an incomplete explanation of the predatory mechanisms of ferocious fish on P. clarkii in the wild. Additionally, Poyang Lake harbors many ferocious fish species, whose predation may still impact the resources of P. clarkii. Future research should focus on the effects of predation by different carnivorous fish on P. clarkii to refine the understanding of how ferocious fish predation affects the resources of P. clarkii.
Author Contributions
M.Z. performed writing—original draft, validation, conceptualization, data curation, and formal analysis. Y.F. and J.C. performed formal analysis, software, data collection, and data analysis. H.W., Y.Z., H.Z. and Y.Y. helped with data collection, the literature search, software, and investigation. G.F. performed writing—review and editing, conceptualization, methodology, supervision, and project administration. C.S. and W.X. performed writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Innovative and Entrepreneurial Talents Project of Jiangxi Province, the funding number is jxsq2020102109.
Institutional Review Board Statement
The experiments comply with current laws in China.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the article. Further information is available upon request from the corresponding author.
Acknowledgments
The authors would like to thank the members of the Jiangxi Fisheries Research Institute for helping with managing the crayfish and catfish.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Chen, T.; Guo, J.Y.; Tang, J.Q.; Huang, C. The differences of growth and living strategies between burrow and corner habitat in Procambarus clarkii. J. Nanjing Univ. 2011, 47, 635–641. [Google Scholar]
- Figler, M.H.; Blank, G.S.; Peeke, H.V.S. Shelter competition between resident male red swamp crayfish Procambarus clarkii (Girard) and conspecific intruders varying by sex and reproductive status. Mar. Freshw. Behav. Physiol. 2005, 38, 237–248. [Google Scholar] [CrossRef]
- Zou, J.; Wang, Y.; Zhong, Y.; Zhu, C.; Zhang, X.; Shi, L.; Zhou, X. Current distribuiton of Procambarus clarkii (Crustacea, Decapoda, Cambaridae) in Poyang Lake and the preliminary morphological cluster analysis of different populaitons. Resour. Environ. Yangtze Basin 2014, 23, 415–421. [Google Scholar]
- Zhang, Y.P.; Chen, W.J.; Fang, C.L. Analysis of the fishing population structure of Procambarus clarkiii in Poyang Lake area. Jiangxi Fish. Sci. Technol. 2014, 2, 6–9. [Google Scholar]
- He, G.; Feng, G.P.; Fu, H.Y.; Wang, H.H.; Zhang, Y.P.; Zhang, A.F.; Ma, B.H.; Yuan, J.X. Distribution characteristics and morphological Differences of the Procambarus Clarkii population in Poyang Lake after the fishing ban. Jiangxi Fish. Sci. Technol. 2024, 1, 1–6. [Google Scholar]
- Wang, C.L.; Qin, J.P.; Huang, C. Behavioral responses of juvenile and subadult crayfishes(Procambarus clarkii) to its predator, the soft-shelled turtle Trionyx sinensis. J. Biosaf. 2013, 22, 169–172. [Google Scholar]
- Song, Z.K. A brief discussion on the methods of preventing natural enemies by cultivating crayfish in paddy field. Henan Agric. 2019, 1, 52. [Google Scholar]
- Blake, M.A.; Hart, P.J.B. The vulnerability of juvenile signal crayfish to perch and eel predation. Freshw. Biol. 1995, 33, 233–244. [Google Scholar] [CrossRef]
- Tierney, A.J.; Godleski, M.S.; Massanari, J.R. Comparative analysis of agonistic behavior in four crayfish species. J. Crustac. Biol. 1992, 20, 54–66. [Google Scholar] [CrossRef]
- Mace, M.M.; Iiirozas, L.P. Fish Predation on Juvenile Penaeid Shrimp: Examining Relative Predator Impact and Size-Selective Predation. Estuaries Coasts 2018, 41, 2128–2134. [Google Scholar] [CrossRef]
- Tyser, A.; Douthwaite, R. Predation on invasive redclaw crayfish Cherax quadricarinatus by native fishes in the Kafue River, Zambia. Afr. J. Aquat. Sci. 2014, 39, 473–477. [Google Scholar] [CrossRef]
- Jones, C.M.; Ruscoe, I.M. Assessment of Five Shelter Types in the Production of Redclaw Crayfish Cherax quadricarinatus (Decapoda: Parastacidae) Under Earthen Pond Conditions. J. World Aquac. Soc. 2001, 32, 41–52. [Google Scholar] [CrossRef]
- Huang, C.; Xiong, Q.H.; Tang, J.Q.; Wu, M. Shadow area and partitioning influencing mortality, healthiness and growth of juvenile Red Swamp Crayfish Procambarus clarkii (Decapoda). Aquac. Res. 2012, 43, 1677–1686. [Google Scholar] [CrossRef]
- Björn, S. Interspecific dominance relationship and aggressive interactions in the freshwater crayfishes Astacus astacus (L.) and Pacifastacus leniusculus (Dana). Can. J. Zool. 1991, 69, 1321–1325. [Google Scholar]
- Sáez-Royuela, M.; Carral, J.M.; Celada, J.D.; Pérez, J.R. Effects of shelter type and food supply frequency on survival and growth of stage-2 juvenile white-clawed crayfish (Austropotamobius pallipes Lereboullet) under laboratory conditions. Aquac. Int. 2001, 9, 489–497. [Google Scholar] [CrossRef]
- Amaral, V.; Paula, J.; Hawkins, S.; Jenkins, S. Cannibalistic interactions in two co-occurring decapod species: Effects of density, food, alternative prey and habitat. J. Exp. Mar. Biol. Ecol. 2009, 368, 88–93. [Google Scholar] [CrossRef]
- Capelli, G.M.; Hamilton, P.A. Effects of food and shelter on aggressive activity in the crayfish Orconectes rusticus (Girard). J. Crustac. Biol. 1984, 4, 252–260. [Google Scholar] [CrossRef]
- Tang, J.Q.; Song, S.L.; Pan, J.L.; Huang, C. The preference to Artificial caves by Procambarus clarkii. Fish. Sci. 2004, 23, 26–28. [Google Scholar]
- Luo, L.; Huang, J.; Liu, D.; Tan, L.; Jiang, S.; Zhou, F.; Jiang, S.; Yang, Q.; Yang, L. Study on selection and spatial distribution of two kinds of caverns for Procambarus clarkii. South China Fish. Sci. 2019, 15, 76–81. [Google Scholar]
- Lodge, D.M. Factors governing species composition, population size, and productivity of cool-water crayfishes. Nord. J. Freshw. Res. 1994, 69, 111–136. [Google Scholar]
- Song, G.; Ding, F.; Chen, J.; Wu, S.; Wang, X. Effects of broodstock sizes, shelter, illumination and stocking density on breeding in red swamp crayfish Procambarus clarkii. Fish. Sci. 2012, 31, 549–553. [Google Scholar]
- Jiang, G.Y.; Yan, W.H. Influence of Different Shroud on the Hatching Rate of the Fertilized Eggs of Procambarus clarkii. Shandong Fish. 2007, 32, 9–10. [Google Scholar]
- Jie, H.; Gao, Y.; Wang, W.; Xie, J.J.; Shi, H.; Wang, G.S.; Xu, W.J. Limb autotomy patterns in the juvenile swimming crab (Portunus trituberculatus) in earth ponds. Aquaculture 2016, 463, 189–192. [Google Scholar]
- Kitagawa, A.T.; Costa, L.S.; Paulino, R.R.; Luz, R.K.; Rosa, P.V.; Guerra-Santos, B.; Fortes-Silva, R. Feeding behavior and the effect of photoperiod on the performance and hematological parameters of the pacamã catfish (Lophiosilurus alexandri). Appl. Anim. Behav. Sci. 2015, 171, 211–218. [Google Scholar] [CrossRef]
- Mattos, B.O.D.; Nascimento-Filho, E.C.T.; Anjos-Santos, A.D.; Sánchez-Vázquez, F.J.; Fortes-Silva, R. Daily self-feeding activity rhythms and dietary self-selection of pirarucu (Arapaima gigas). Aquaculture 2016, 465, 152–157. [Google Scholar] [CrossRef]
- Chen, S.B.; Chen, W.X.; Fang, Z.T. Effect of Water Temperature on Feeding Rhythm in Common Carp (Cyprinus carpio haematopterus Temminck et Schlegel). J. Northeast Agric. Univ. 2012, 19, 57–61. [Google Scholar]
- Gao, X.; Pang, G.; Luo, X.; You, W.; Ke, C. Effects of light cycle on circadian feeding activity and digestive physiology in Haliotis discus hannai. Aquaculture 2021, 539, 736642. [Google Scholar] [CrossRef]
- Xie, C.X.; Xiong, C.X.; Zhou, J.; Wan, X.M.; Jin, H. Feeding intensity and dynamics of juvenile northern snakehead, Channa argus, under different illumination. Acta Hydrobiol. Sin. 1997, 21, 213–218. [Google Scholar]
- Qiao, Z.G.; Zhang, G.L.; Zhang, Y.Y.; Peng, X.L. Diel feeding rhythm of juvenile oriental sheatfish Silurus asotus in different photoperiods. Chin. J. Ecol. 2008, 27, 791–796. [Google Scholar]
- Savino, J.F.; Stein, R.A. Behavior of fish predators and their prey: Habitat choice between open water and dense vegetation. Environ. Biol. Fishes 1989, 24, 287–293. [Google Scholar] [CrossRef]
- Heck, K.L.; Crowder, L.B. Habitat structure and predator—Prey interactions in vegetated aquatic systems. In Habitat Structure; Springer: Dordrecht, The Netherlands, 1991. [Google Scholar]
- Alexander, M.E.; Dick, J.T.A.; O’Connor, N.E. Born to kill: Predatory functional responses of the littoral amphipod Echinogammarus marinus Leach throughout its life history. J. Exp. Mar. Biol. Ecol. 2013, 439, 92–99. [Google Scholar] [CrossRef]
- Yu, J.X.; Xiong, M.T.; Ye, S.W.; Li, W.; Zhang, T.L. Effects of stocking density and artificial macrophyte shelter on survival, growth and molting of juvenile red swamp crayfish (Procambarus clarkii) under experimental conditions. Aquaculture 2020, 521, 735001. [Google Scholar] [CrossRef]
- Wu, L.M.; Han, G.M.; Tan, B.L.; Zhang, J.H.; Wang, S.H. Effects of different aquatic plants on growth performance, physiological indices and muscle nutrients of red swamp crayfish (Procambarus clarkii). J. Dalian Ocean Univ. 2023, 38, 779–786. [Google Scholar]
- Stoner, A.W. The influence of benthic macrophytes on the foraging behavior of pinfish, Lagodon rhomboides (Linnaeus). J. Exp. Mar. Biol. Ecol. 1982, 58, 271–284. [Google Scholar] [CrossRef]
- Howard, R.K.; Koehn, J.D. Population dynamics and feeding ecology of pipefish (Syngnathidae) associated with eelgrass beds of Western Port, Victoria. Mar. Freshw. Res. 1985, 36, 361–370. [Google Scholar] [CrossRef]
- Ostrand, K.G.; Braeutigam, B.J.; Wahl, D.H. Consequences of Vegetation Density and Prey Species on Spotted Gar Foraging. Trans. Am. Fish. Soc. 2004, 133, 794–800. [Google Scholar] [CrossRef]
- Gotceitas, V. Variation in plant stem density and its effects on foraging success of juvenile bluegill sunfish. Environ. Biol. Fishes 1990, 27, 63–70. [Google Scholar] [CrossRef]
- Figler, M.H.; Cheverton, H.M.; Blank, G.S. Shelter competition in juvenile red swamp crayfish (Procambarus clarkii): The influences of sex differences, relative size, and prior residence. Aquaculture 1999, 178, 63–75. [Google Scholar] [CrossRef]
- Everett, R.A.; Ruiz, G.M. Coarse woody debris as a refuge from predation in aquatic communities: An experimental test. Oecologia 1993, 93, 475–486. [Google Scholar] [CrossRef]
- Karnofsky, E.B.; Atema, J.; Elgin, R.H. Natural Dynamics of Population Structure and Habitat Use of the Lobster, Homarus americanus, in a Shallow Cove. Biol. Bull. 1989, 176, 247–256. [Google Scholar] [CrossRef]
- Vince, S.; Valiela, I.; Backus, N.; Teal, J.M. Predation by the salt marsh killifish Fundulus heteroclitus (L.) in relation to prey size and habitat structure: Consequences for prey distribution and abundance. J. Exp. Mar. Biol. Ecol. 1976, 23, 255–266. [Google Scholar] [CrossRef]
- Stein, R.A. Selective Predation, Optimal Foraging, and Resource Depression within the Predator-Prey Interaction between Fish and Crayfish; University of Wisconsin-Madison: Madison, WI, USA, 1975. [Google Scholar]
- Valley, R.D.; Bremigan, M.T. Effects of Macrophyte Bed Architecture on Largemouth Bass Foraging: Implications of Exotic Macrophyte Invasions. Trans. Am. Fish. Soc. 2002, 131, 234–244. [Google Scholar] [CrossRef]
- Eggleston, D.B.; Lipcius, R.N. Shelter selection by spiny lobster under variable predation risk, social conditions, and shelter size. Ecology 1992, 73, 992–1011. [Google Scholar] [CrossRef]
- Mintz, J.D.; Lipcius, R.N.; Eggleston, D.B.; Seebo, M.S. Survival of juvenile caribbean spiny lobster: Effects of shelter size, geographic location and conspecific abundance. Mar. Ecol. Prog. Ser. 1994, 112, 255. [Google Scholar] [CrossRef]
- Nilsson, P.A.; Bronmark, C. Prey Vulnerability to a Gape-Size Limited Predator: Behavioural and Morphological Impacts on Northern Pike Piscivory. Oikos 2008, 88, 539–546. [Google Scholar] [CrossRef]
- Webb, P.W. Effect of Body Form and Response Threshold on the Vulnerability of Four Species of Teleost Prey Attacked by Largemouth Bass (Micropterus salmoides). Can. J. Fish. Aquat. Sci. 2011, 43, 763–771. [Google Scholar] [CrossRef]
- Huang, W.; Hu, T.; Mao, J.; Montzka, C.; Bol, R.; Wan, S.; Li, J.; Yue, J.; Dai, H. Hydrological Drivers for the Spatial Distribution of Wetland Herbaceous Communities in Poyang Lake. Remote Sens. 2022, 14, 4870. [Google Scholar] [CrossRef]
- Tan, Z.Q.; Xu, X.L.; Li, Y.L.; Jiang, J.H.; Zhang, Q. Wetland landscape pattern evolution of large yangtze-connected lakes in the middle reaches of yangtze river. Resour. Environ. Yangtze Basin 2017, 26, 1619–1624. [Google Scholar]
- Eversole, A.G.; Mazlum, Y. Comparative fecundity of three Procambarus species. J. Shellfish Res. 2002, 21, 255–258. [Google Scholar]
- Lei, X.Q.; Yan, B.H.; Yao, Y.; Jiang, Q.L.; Wang, Y.L. Study on biological characteristics and propagation techniques of wheat ear fish. Jiangxi Fish. Sci. Technol. 2013, 16–18. [Google Scholar]
- Xie, D.S.; Luo, L.Z. Diapause and its roles in the population dynamics of the beet webworm, Loxostege sticticalis. Plant Prot. 2011. [Google Scholar]
- Zhao, M.G.; Chen, J.H.; Feng, G.P.; Wang, H.H.; Zhang, Y.P. The Predation Selectivity of Ferocious Fish (Silurus asotus and Channa argus) to Procambarus clarkii Larvae in Poyang Lake. Acta Hydrobiol. Sin. 2024, 48, 799–807. [Google Scholar]
- Correia, A.M.; Ferreira, Ó. Burrowing behavior of the introduced red swamp crayfish Procambarus clarkii (Decapoda: Cambaridae) in Portugal. J. Crustac. Biol. 1995, 15, 248–257. [Google Scholar] [CrossRef]
- Groza, M.I.; Pop-Vancia, V.; Mireşan, V. Diel activity and use of multiple artificially constructed shelters in Astacus leptodactylus (Decapoda: Astacidae). Biologia 2016, 71, 1369–1379. [Google Scholar] [CrossRef]
- Li, R.R.; Kaufman, Y.J.; Gao, B.C.; Davis, C.O. Remote sensing of suspended sediments and shallow coastal waters. IEEE Trans. Geosci. Remote Sens. 2003, 41, 559–566. [Google Scholar]
- Levin, I.M.; Radomyslskay, T.M. Estimate of water inherent optical properties from secchi depth. Izv. Atmos. Ocean. Phys. 2012, 48, 214–221. [Google Scholar] [CrossRef]
- Zhu, J.Q.; Li, M.Y.; Wu, X.K. Study on the growth and feeding rhythm of larval and juvenile of Lateolabrax Japonicus. J. Zhejiang Ocean Univ. 2002, 21, 210–215. [Google Scholar]
- Li, K.G.; Zhen, D.S.; Fu, S.J. Effect of feeding on excess post-exercise oxygen consumption in juvenile chinese catfish (Silurus asotus Linnaeus). Acta Hydrobiol. Sin. 2012, 36, 1035–1040. [Google Scholar] [CrossRef]
- Perry, W.L.; Lodge, D.M.; Lamberti, G.A. Impact of crayfish predation on exotic zebra mussels and native invertebrates in a lake-outlet stream. Can. J. Fish. Aquat. Sci. 1997, 54, 120–125. [Google Scholar] [CrossRef][Green Version]
- Dorn, N.J.; Mittelbach, G.G. More than predator and prey: A review of interactions between fish and crayfish. Life Environ. 1999, 49, 229–237. [Google Scholar]
- Hoyle, J.A.; Allen, K. The effect of prey morphology and size on handling time in a piscivore, the largemouth bass (Micropterus salmoides). Can. J. Zool. 1987, 65, 1972–1977. [Google Scholar] [CrossRef]
- Gill, A.B. The dynamics of prey choice in fish: The importance of prey size and satiation. J. Fish Biol. 2010, 63, 105–116. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).