

Parasitic Characteristics of Ticks (Acari: Ixodidae) Collected from Water Deer (Hydropotes inermis argyropus) and Spatiotemporal Distribution Prediction within Host-Influenced Cultivated Areas


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Capturing Water Deer and Detection Site Coordinates
2.3. Collection and Identification of Water Deer and Parasitic Ixodid Ticks
2.4. Statistical Analysis
2.5. Cropland Use Characterization by Water Deer and Parasitic Ixodid Tick Distribution Prediction
3. Results
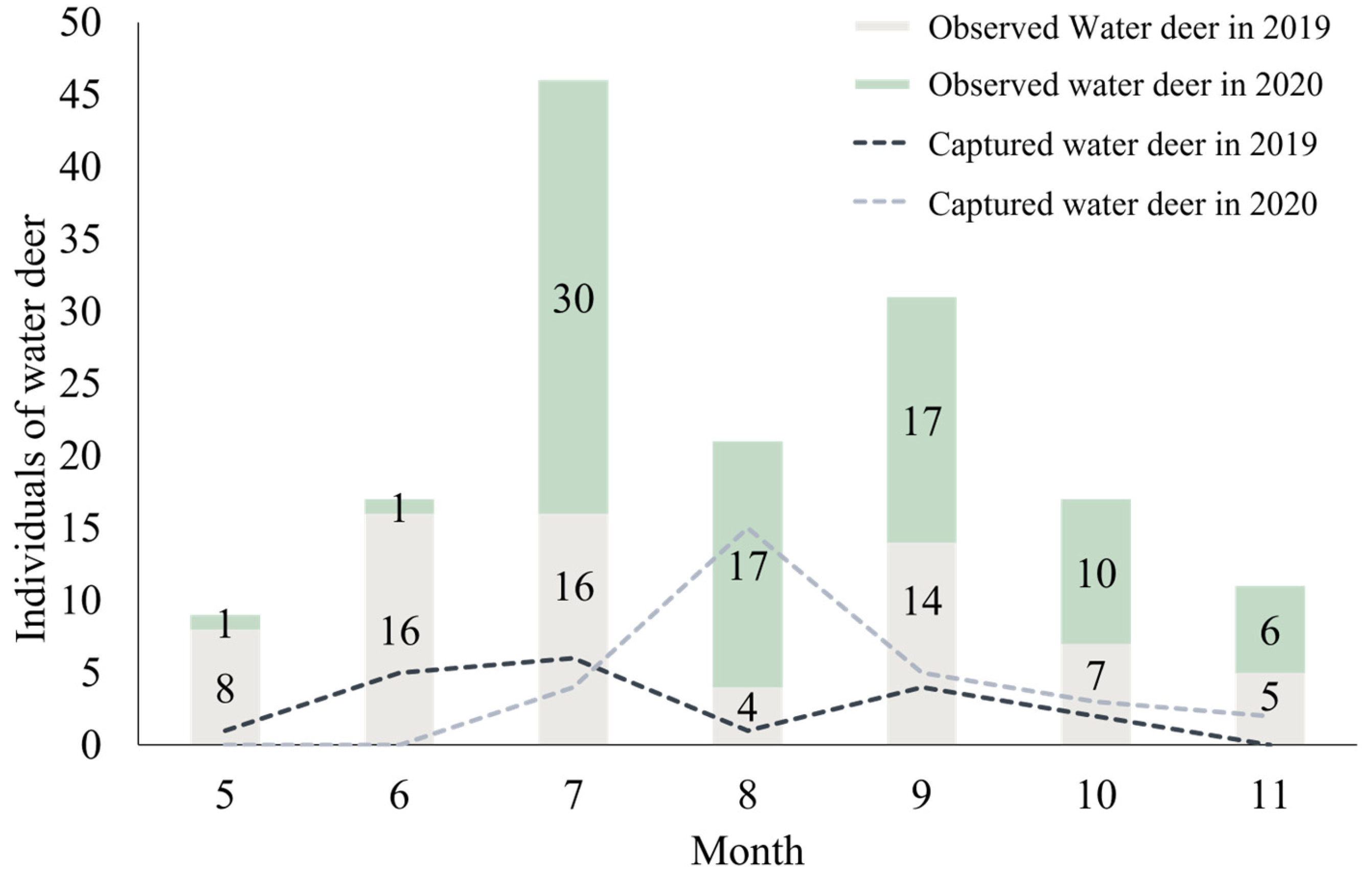
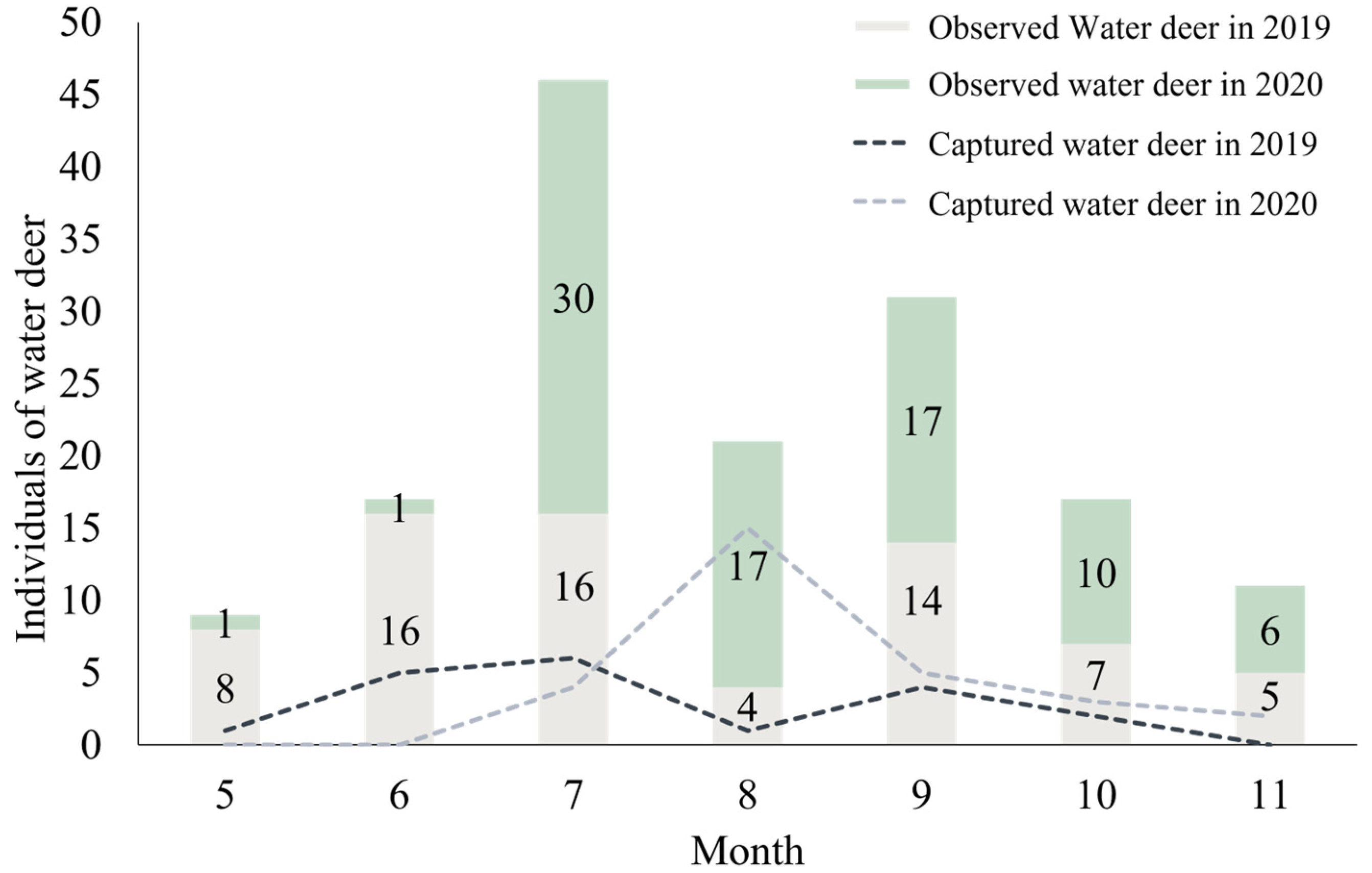
3.1. Capturing Water Deer
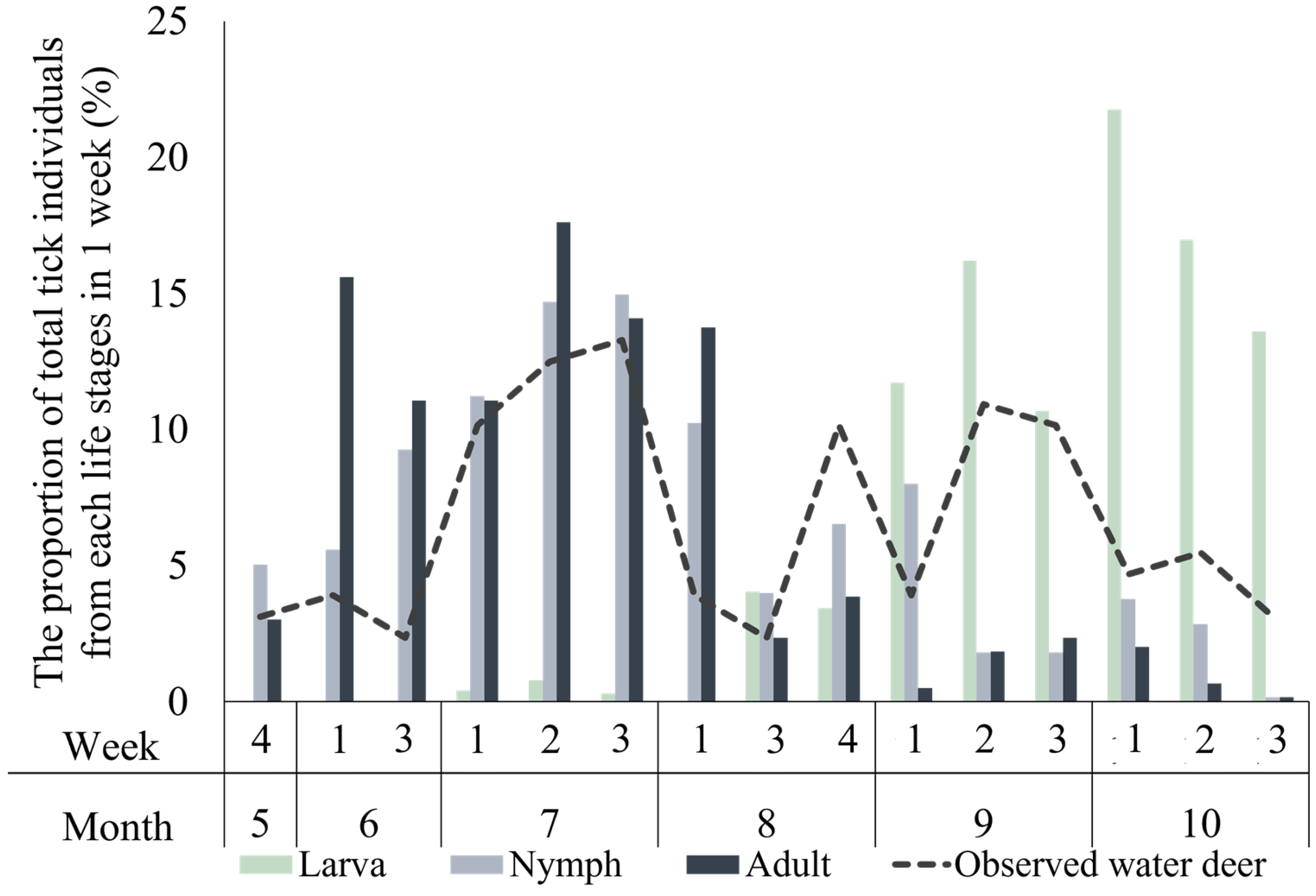
3.2. Water Deer and Parasitic Ixodid Tick Communities
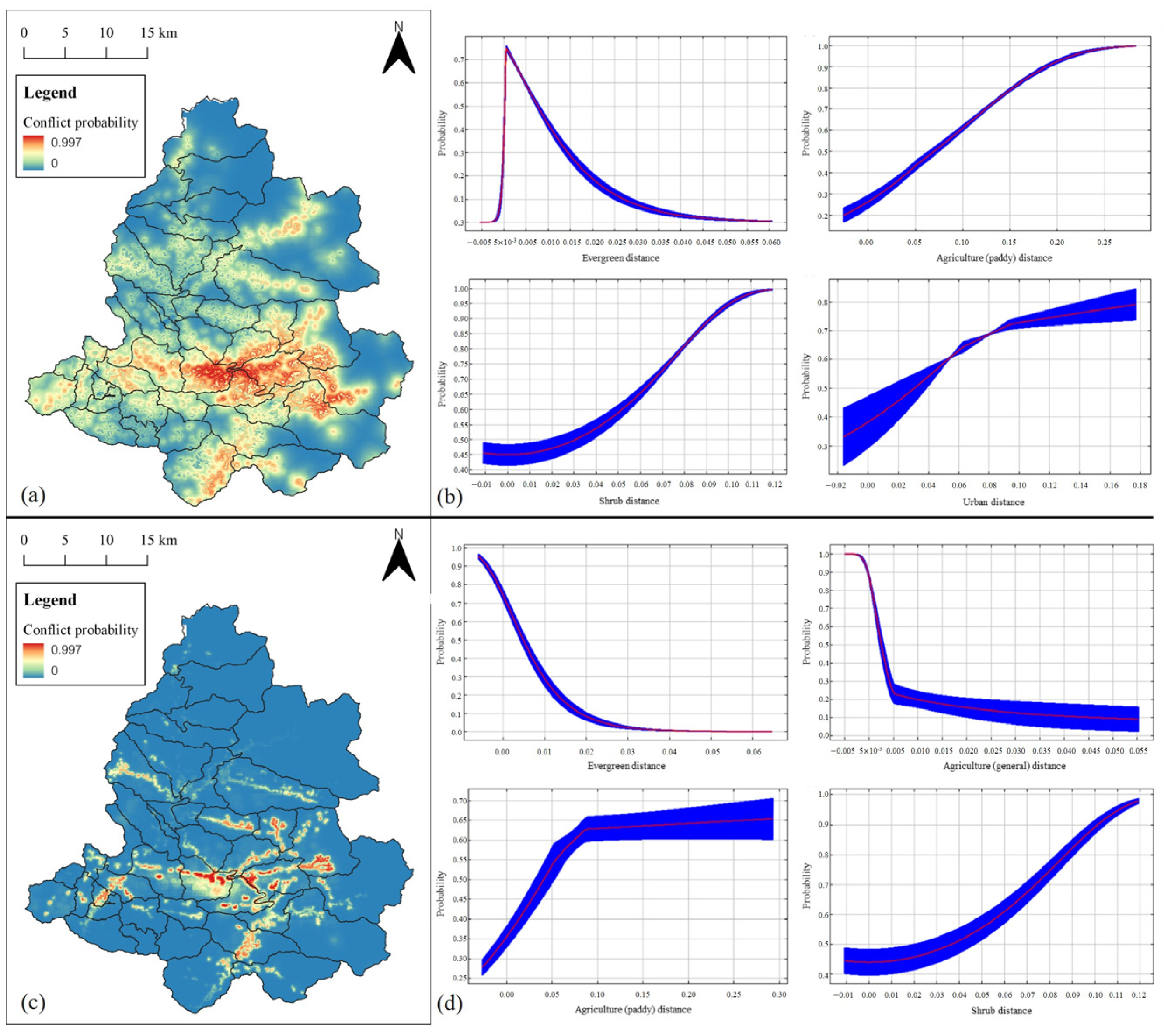
3.3. Prediction of Water Deer Distribution
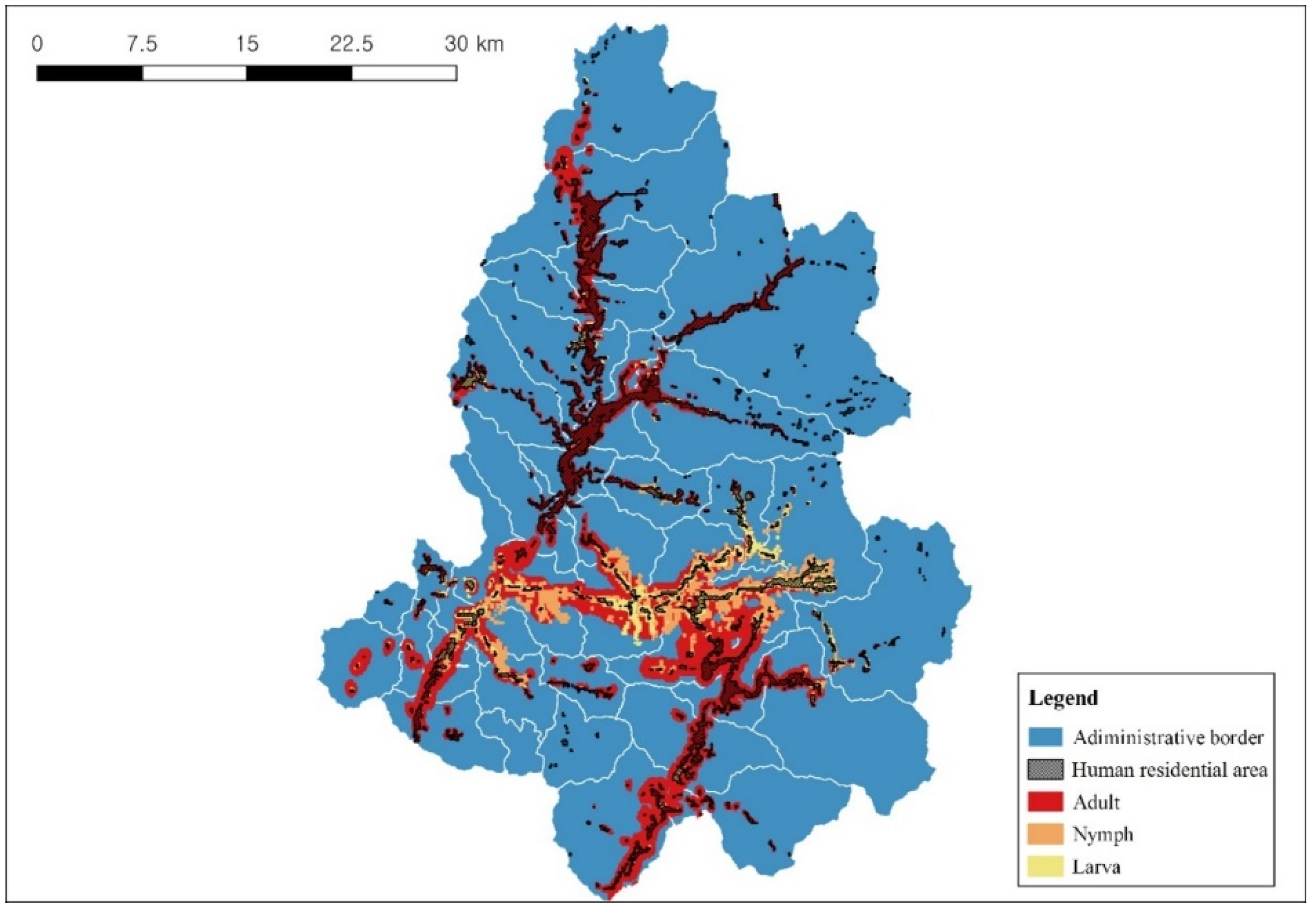
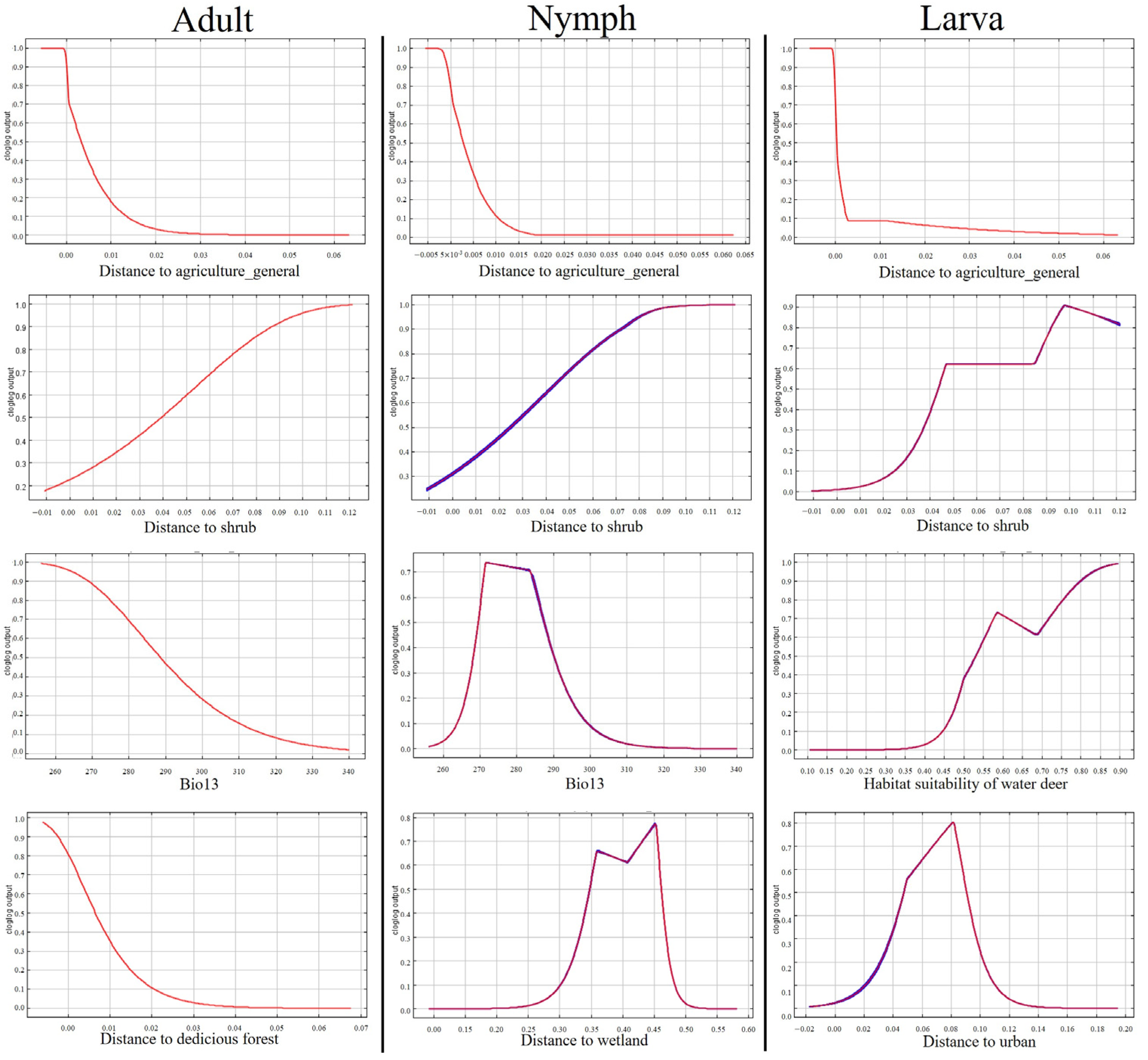
3.4. Distribution Prediction of Parasitic Ixodid Ticks
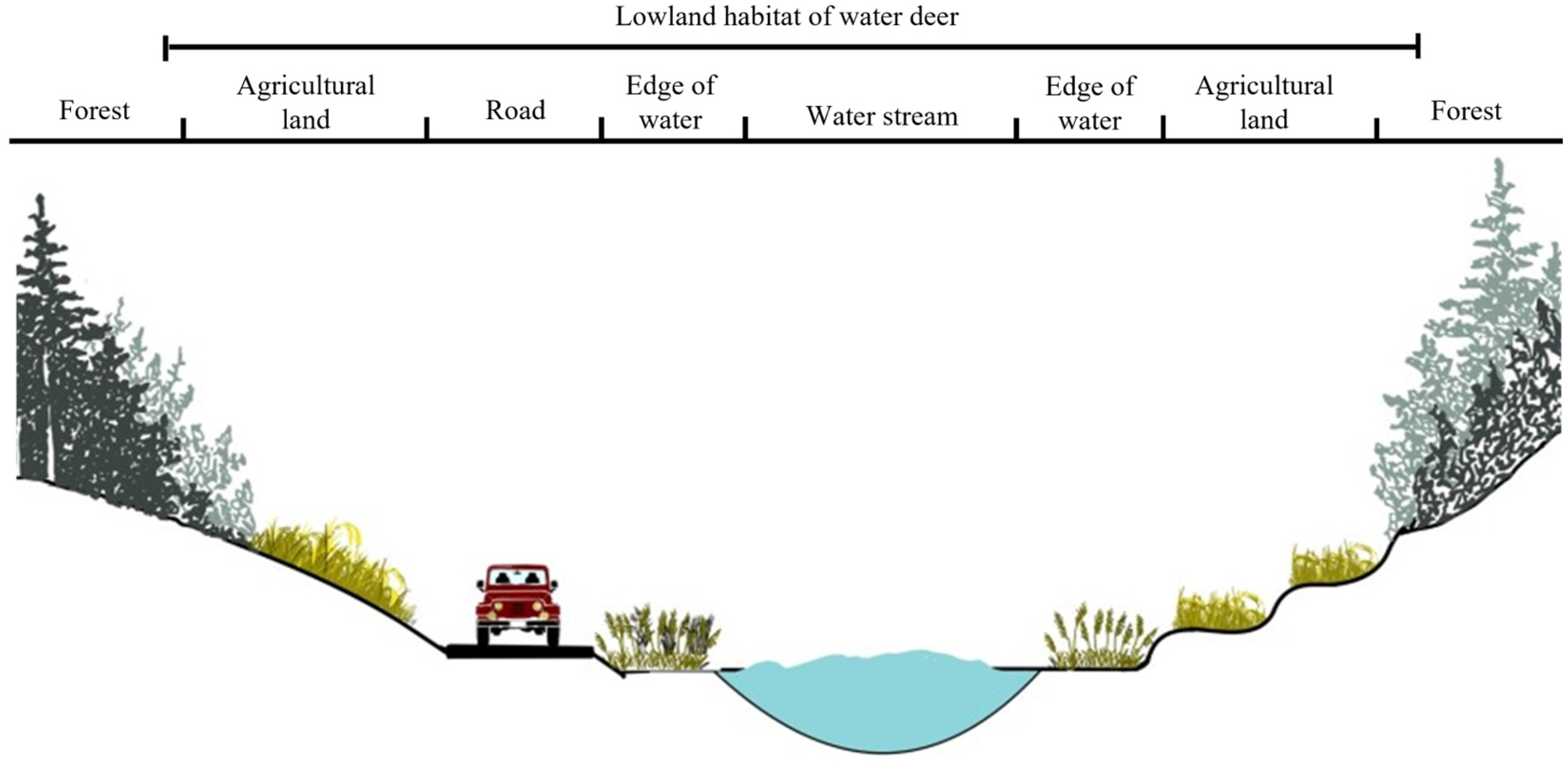
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paquet, P.C.; Shelley, M.A. Habitat loss: Changing how animals think? In Animal Welfare in a Changing World; CAB International: London, UK, 2018; pp. 4–13. [Google Scholar]
- Escobar, L.E.; Awan, M.N.; Qiao, H. Anthropogenic disturbance and habitat loss for the red-listed Asiatic black bear (Ursus thibetanus): Using ecological niche modeling and nighttime light satellite imagery. Biol. Conserv. 2015, 191, 400–407. [Google Scholar] [CrossRef]
- Nyhus, P.J. Human–wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 2016, 41, 143–171. [Google Scholar] [CrossRef]
- World Health Organization; WHO Expert Committee on the Selection; Use of Essential Medicines; World Health Organization. The Selection and Use of Essential Medicines: Report of the WHO Expert Committee; Including the 18th WHO Model List of Essential Medicines and the 4th WHO Model List of Essential Medicines for Children, 985; WHO Press: Geneva, Switzerland, 2013. [Google Scholar]
- Korea Disease Control and Prevention Agency. Guidelines for Managing Tick & Rodent Borne Disease; KCDC: Cheongju, Republic of Korea, 2020.
- Patrick, C.D.; Hair, J.A. Oviposition behavior and larval longevity of the lone star tick, Amblyomma americanum (Acarina: Ixodidae), in different habitats. Ann. Entomol. Soc. Am. 1979, 72, 308–312. [Google Scholar] [CrossRef]
- Bowman, A.S.; Ball, A.; Sauer, J.R. Tick salivary glands: The physiology of tick water balance and their role in pathogen trafficking and transmission. Ticks Biol. Contr. 2008, 73–91. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.C.; Golovljova, I.; Jaenson, T.G.; Jensen, J.K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasit. Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H.; Aeschlimann, A. Tick-host specificity. Bull. Société Entomol. Suisse 1982, 55, 5–32. [Google Scholar]
- Van Buskirk, J.; Ostfeld, R.S. Controlling Lyme disease by modifying the density and species composition of tick hosts. Ecol. Appl. 1995, 5, 1133–1140. [Google Scholar] [CrossRef]
- Kilpatrick, H.J.; Labonte, A.M.; Stafford III, K.C. The relationship between deer density, tick abundance, and human cases of Lyme disease in a residential community. J. Med. Entomol. 2014, 51, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Camin, J.H.; Drenner, R.W. Climbing behavior and host-finding of larval rabbit ticks (Haemaphysalis leporispalustris). J. Parasitol. 1978, 64, 905–909. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.F.; Schmidtmann, E.T. Dispersal of blacklegged tick (Acari: Ixodidae) nymphs and adults at the woods–pasture interface. J. Med. Entomol. 1996, 33, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Sumner, T.; Orton, R.J.; Green, D.M.; Kao, R.R.; Gubbins, S. Quantifying the roles of host movement and vector dispersal in the transmission of vector-borne diseases of livestock. PLoS Comput. Biol. 2017, 13, e1005470. [Google Scholar] [CrossRef] [PubMed]
- Johannesen, J.; Foissac, X.; Kehrli, P.; Maixner, M. Impact of vector dispersal and host-plant fidelity on the dissemination of an emerging plant pathogen. PLoS ONE 2012, 7, e51809. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Pietzsch, M.E.; Rice, N.V.P.; Jones, L.; Kerrod, E.; Avenell, D.; Los, S.; Ratcliffe, N.; Leach, S.; Butt, T. Investigation of ecological and environmental determinants for the presence of questing Ixodes ricinus (Acari: Ixodidae) on Gower, South Wales. J. Med. Entomol. 2008, 45, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Ellis, V.A.; Collins, M.D.; Medeiros, M.C.; Sari, E.H.; Coffey, E.D.; Dickerson, R.C.; Lugarini, C.; Stratford, J.A.; Henry, D.R.; Merrill, L.; et al. Local host specialization, host-switching, and dispersal shape the regional distributions of avian haemosporidian parasites. Proc. Natl. Acad. Sci. USA 2015, 112, 11294–11299. [Google Scholar] [CrossRef]
- Gaitan, J.; Millien, V. Stress level, parasite load, and movement pattern in a small-mammal reservoir host for Lyme disease. Can. J. Zool. 2016, 94, 565–573. [Google Scholar] [CrossRef]
- Jo, Y.S.; Baccus, J.T.; Koprowski, J.L. Artiodactyla. In Mammals of Korea; National Institute of Biological Resources: Incheon, Republic of Korea, 2018; pp. 293–295. [Google Scholar]
- Kim, B.J.; Lee, N.S.; Lee, S.D. Feeding diets of the korean water deer (Hydropotes inermis argyropus) based on a 202 bp rbcL sequence analysis. Conserv. Genet. 2011, 12, 851–856. [Google Scholar] [CrossRef]
- Nam, T.W.; Park, S.C.; Han, B.H. A study on the characteristics of inhabitation environment of Hydropotes inermis in Daebudo Island, Ansan-si. J. Korean Environ. Restor. Technol. 2020, 23, 45–58. [Google Scholar]
- Kim, C.M.; Park, S.Y.; Kim, D.M.; Park, J.W.; Chung, J.K. First report of Borrelia burgdorferi sensu stricto detection in a commune genospecies in Apodemus agrarius in Gwangju, South Korea. Sci. Rep. 2021, 11, 18199. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Y.; Li, H.; Zhu, W.; Darman, Y.; Lee, D.K.; Wang, T.; Sedash, G.; Pandey, P.; Borzée, A.; et al. Prediction of range expansion and estimation of dispersal routes of water deer (Hydropotes inermis) in the transboundary region between China, the Russian Far East and the Korean Peninsula. PLoS ONE 2022, 17, e0264660. [Google Scholar] [CrossRef] [PubMed]
- Ministry of the Environment. Status of Damage by Harmful Wildlife by Year (14~18). Available online: https://me.go.kr (accessed on 27 October 2022).
- Kim, K.; Woo, D.G.; Seo, H.; Park, T.; Song, E.G.; Choi, T.Y. Korea road-kill observation system: The first case to integrate road-kill data in national scale by government. J. For. Env. Sci. 2019, 35, 281–284. [Google Scholar] [CrossRef]
- Li, Z.; Hu, J.; Bao, C.; Li, P.; Qi, X.; Qin, Y.; Wang, S.; Tan, Z.; Zhu, Y.; Tang, F.; et al. Seroprevalence of antibodies against SFTS virus infection in farmers and animals, Jiangsu, China. J. Clin. Virol. 2014, 60, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Fukuma, A.; Shimojima, M.; Yamashita, Y.; Mizota, F.; Yamashita, M.; Otsuka, Y.; Kan, M.; Fukushi, S.; Tani, H.; et al. Seroprevalence of severe fever with thrombocytopenia syndrome (SFTS) virus antibodies in humans and animals in Ehime Prefecture, Japan, an endemic region of SFTS. J. Infect. Chemother. 2018, 24, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Korea Disease Control Agency. Available online: http://www.kdca.ac.kr (accessed on 27 October 2022).
- Kim, H.C.; Han, S.H.; Chong, S.T.; Klein, T.A.; Choi, C.Y.; Nam, H.Y.; Chae, H.Y.; Lee, H.; Ko, S.; Kang, J.G.; et al. Ticks collected from selected mammalian hosts surveyed in the Republic of Korea during 2008–2009. Korean J. Parasitol. 2011, 49, 331–335. [Google Scholar] [CrossRef]
- Kim, B.J.; Kim, H.; Won, S.; Kim, H.C.; Chong, S.T.; Klein, T.A.; Kim, K.G.; Seo, H.Y.; Chae, J.S. Ticks collected from wild and domestic animals and natural habitats in the Republic of Korea. Korean J. Parasitol. 2014, 52, 281–285. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.S.; Chae, J.B.; Kang, J.G.; Kim, H.C.; Chong, S.T.; Shin, J.H.; Hur, M.S.; Suh, J.H.; Oh, M.D.; Jeong, S.M.; et al. Detection of severe fever with thrombocytopenia syndrome virus from wild animals and Ixodidae ticks in the Republic of Korea. Vector Borne Zoonotic Dis. 2016, 16, 408–414. [Google Scholar] [CrossRef]
- VanBik, D.; Lee, S.H.; Seo, M.G.; Jeon, B.R.; Goo, Y.K.; Park, S.J.; Rhee, M.H.; Kwon, O.D.; Kim, T.H.; Geraldino, P.J.L.; et al. Borrelia species detected in ticks feeding on wild Korean water deer (Hydropotes inermis) using molecular and genotypic analyses. J. Med. Entomol. 2017, 54, 1397–1402. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Song, B.G.; Shin, E.H.; Yun, S.M.; Han, M.G.; Park, M.Y.; Park, C.; Ryou, J. Prevalence of severe fever with thrombocytopenia syndrome virus in Haemaphysalis longicornis ticks in South Korea. Ticks Tick-Borne Dis. 2014, 5, 975–977. [Google Scholar] [CrossRef] [PubMed]
- Im, J.H.; Baek, J.; Durey, A.; Kwon, H.Y.; Chung, M.H.; Lee, J.S. Current status of tick-borne diseases in South Korea. Vector Borne Zoonotic Dis. 2019, 19, 225–233. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, T.K.; Kim, T.Y.; Lee, H.I. Geographical distribution of Borrelia burgdorferi sensu lato in ticks collected from wild rodents in the Republic of Korea. Pathogens 2020, 9, 866. [Google Scholar] [CrossRef] [PubMed]
- Yamaguti, N.; Tipton, V.J.; Keegan, H.L.; Toshioka, S. Ticks of Japan, Korea, and the Ryukyu Islands; Brigham Young University Sience Bulletin, Biological Series; Brigham Young University: Provo, UT, USA, 1971; Volume 15, p. 1. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Zhang, E. Daytime activity budgets of the Chinese water deer. Mammalia 2000, 64, 163–172. [Google Scholar] [CrossRef]
- Park, J.E.; Kim, B.J.; Oh, D.H.; Lee, H.; Lee, S.D. Feeding habit analysis of the Korean water deer. Korean J. Environ. Ecol. 2011, 25, 836–845. [Google Scholar]
- Carroll, J.F.; Klun, J.A.; Schmidtmann, E.T. Evidence for kairomonal influence on selection of host-ambushing sites by adult Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 1995, 32, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Rechav, Y. Migration and dispersal patterns of three African ticks (Acari: Ixodidae) under field conditions. J. Med. Entomol. 1979, 16, 150–163. [Google Scholar] [CrossRef]
- Yano, Y.; Shiraishi, S.; Uchida, T.A. Effects of temperature on development and growth in the tick, Haemaphysalis longicornis. Exp. Appl. Acarol. 1987, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Despins, J.L. Effects of temperature and humidity on ovipositional biology and egg development of the tropical horse tick, Dermacentor (Anocentor) nitens. J. Med. Entomol. 1992, 29, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Buczek, A. Experimental teratogeny in the tick Hyalomma marginatum marginatum (Acari: Ixodida: Ixodidae): Effect of high Humidity on embryonic development. J. Med. Entomol. 2000, 37, 807–814. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Moon, K.; Kim, M.; Lee, W.G.; Lee, H.I.; Park, J.K.; Kim, Y.H. Seasonal distribution of Haemaphysalis longicornis (Acari: Ixodidae) and detection of SFTS virus in Gyeongbuk Province, Republic of Korea, 2018. Acta Trop. 2021, 221, 106012. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Lee, D.H. Population dynamics of hard ticks (Acari: Ixodidae) and their harboring rates of Severe Fever with Thrombocytopenia Syndrome (SFTS) virus in four landscapes of Gyeonggi Province, South Korea. Exp. Appl. Acarol. 2023, 91, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Suh, J.H.; Kim, H.C.; Yun, S.M.; Lim, J.W.; Kim, J.H.; Chong, S.T.; Kim, D.H.; Kim, H.T.; Kim, H.; Klein, T.A.; et al. Detection of SFTS virus in Ixodes nipponensis and Amblyomma Testudinarium (Ixodida: Ixodidae) collected from reptiles in the Republic of Korea. J. Med. Entomol. 2016, 53, 584–590. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, K.Y.; Kim, G.; Moon, S.; Park, Y.C.; Cho, H.S.; Oh, Y. Potential tick defense associated with skin and hair characteristics in Korean water deer (Hydropotes inermis argyropus). Animals 2024, 14, 185. [Google Scholar] [CrossRef] [PubMed]
- Chaves, P.P.; Ruokolainen, K.; Van doninck, J.; Tuomisto, H. Impact of spatial configuration of training data on the performance of amazonian tree species distribution models. Forest Ecol. Manag. 2022, 504, 119838. [Google Scholar] [CrossRef]
- Kim, K.Y. Spatiotemporal Distribution of Ectoparasites in Relation to the Habitat Use Characteristics of Mammal Hosts in Cultivated Land and Nearby Habitats in Gangwon-do. Ph.D. Thesis, Kangwon National University, Chuncheon, Republic of Korea, 2023. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Explanation |
|---|---|
| Bio1 | Annual mean temperature |
| Bio2 | Mean diurnal range (mean of monthly (max temp − min temp)) |
| Bio3 | Isothermality (Bio2)/(Bio7) × 100 |
| Bio4 | Temperature seasonality (standard deviation × 100) |
| Bio5 | Maximum temperature of the warmest month |
| Bio6 | Minimum temperature of the coldest month |
| Bio7 | Annual temperature range (Bio5 − Bio6) |
| Bio8 | Mean temperature of the wettest quarter |
| Bio9 | Mean temperature of the driest quarter |
| Bio10 | Mean temperature of the warmest quarter |
| Bio11 | Mean temperature of the coldest quarter |
| Bio12 | Annual precipitation |
| Bio13 | Precipitation of the wettest month |
| Bio14 | Precipitation of the driest month |
| Bio15 | Precipitation seasonality (coefficient of variation) |
| Bio16 | Precipitation of the wettest quarter |
| Bio17 | Precipitation of the driest quarter |
| Bio18 | Precipitation of the warmest quarter |
| Bio19 | Precipitation of the coldest quarter |
| Elevation | Digital elevation model (DEM; GMTED provided by USGS) |
| Slope | Derived from the DEM using the Slope tool in ArcMap |
| Ruggedness | Derived from the DEM—standard deviation of the slope using focal statistics |
| Landcover | Extracted from World Land Cover 30 m (BaseVue 2013), Source: MDA information systems (MDAUS) |
| Deciduous forest (d) | Euclidean distance to deciduous forest land cover class (BaseVue 2013) |
| Evergreen forest (d) | Euclidean distance to evergreen forest land cover class (BaseVue 2013) |
| Shrub (d) | Euclidean distance to shrub (BaseVue 2013) |
| Grassland (d) | Euclidean distance to grassland (BaseVue 2013) |
| Agriculture_general (d) | Euclidean distance to general agricultural distance (BaseVue 2013) |
| Agriculture_paddy (d) | Euclidean distance to agricultural paddy distance (BaseVue 2013) |
| Wetland (d) | Euclidean distance to wetland (BaseVue 2013) |
| Water (d) | Euclidean distance to water (BaseVue 2013) |
| Urban (d) | Euclidean distance to an urban area (BaseVue 2013) |
| H. longicornis | H. flava | H. japonica | I. nipponensis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Month | AF | AM | N | AF | AM | N | AF | AM | N | AF | AM | N |
| 5 | 8.0 | 7.0 | 89.0 | 1.0 | 2.0 | 3.0 | ||||||
| 6 | 44.2 ± 13.7 | 37.6 ± 8.5 | 127.4 ± 56.2 | 1.0 | 3.0 ±0.0 | 1.0 | ||||||
| 7 | 70.5 ± 10.0 | 19.4 ± 3.9 | 267.8 ± 28.9 | 70.5 ± 10.0 | 19.4 ± 3.9 | 3.5 ± 0.6 | 1.5 ± 0.5 | |||||
| 8 | 19.4 ± 9.3 | 7.2 ± 3.5 | 100.2 ± 24.9 | 6.2 ± 3.2 | 1.0 | 3.3 ± 0.9 | 4.0 ± 1.0 | 1.5 ± 0.5 | 4.0 | |||
| 9 | 2.0 ± 0.5 | 54.4 ± 16.1 | 4.0 | 7.5 ± 2.8 | 21.8 ± 5.9 | 4.0 ± 2.0 | 5.8 ± 4.1 | 2.0 | 1.0 | 6.3 ± 3.4 | ||
| 10 | 3.6 ± 1.0 | 1.0 | 5.2 ± 2.2 | 33.5 ± 6.9 | 1.0 | 20.5 ± 6.5 | ||||||
| Month | ||||||
|---|---|---|---|---|---|---|
| 6 | 7 | 8 | 9 | 10 | ||
| Number of Captured Water Deer | 5 (5) | 10 (10) | 16 (9) | 9 (9) | 5 (5) | |
| MI of ixodid ticks | Larva | abc* 2.8 ± 0.7 | ab 47.6 ± 8.7 | a 278.8 ± 4.5 | ac 809.3 ± 235.9 | c 1080.8 ± 82.6 |
| Nymph | a 128.6 ± 56.9 | b 271.6 ± 29.2 | a 101.1 ± 24.5 | a 44.7 ± 9.6 | a 38.6 ± 9.3 | |
| Adult | a 82.2 ± 21.1 | b 91.3 ± 13.0 | ac 24.6 ± 8.8 | c 8.4 ± 3.2 | ac 5.6 ± 2.1 | |
| Season | Larva | Nymph | Adult | |
|---|---|---|---|---|
| Summer | Water deer | ** 0.819 | * 0.609 | 0.138 |
| Larva | 0.446 | −0.144 | ||
| Nymph | * 0.549 | |||
| Fall | Water deer | −0.065 | * 0.583 | 0.187 |
| Larva | −0.469 | 0.014 | ||
| Nymph | 0.297 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.; Kim, K.; Yoon, K.; Park, Y. Parasitic Characteristics of Ticks (Acari: Ixodidae) Collected from Water Deer (Hydropotes inermis argyropus) and Spatiotemporal Distribution Prediction within Host-Influenced Cultivated Areas. Animals 2024, 14, 1153. https://doi.org/10.3390/ani14081153
Kim K, Kim K, Yoon K, Park Y. Parasitic Characteristics of Ticks (Acari: Ixodidae) Collected from Water Deer (Hydropotes inermis argyropus) and Spatiotemporal Distribution Prediction within Host-Influenced Cultivated Areas. Animals. 2024; 14(8):1153. https://doi.org/10.3390/ani14081153
Chicago/Turabian StyleKim, Kiyoon, Kyungmin Kim, Kwangbae Yoon, and Yungchul Park. 2024. "Parasitic Characteristics of Ticks (Acari: Ixodidae) Collected from Water Deer (Hydropotes inermis argyropus) and Spatiotemporal Distribution Prediction within Host-Influenced Cultivated Areas" Animals 14, no. 8: 1153. https://doi.org/10.3390/ani14081153
APA StyleKim, K., Kim, K., Yoon, K., & Park, Y. (2024). Parasitic Characteristics of Ticks (Acari: Ixodidae) Collected from Water Deer (Hydropotes inermis argyropus) and Spatiotemporal Distribution Prediction within Host-Influenced Cultivated Areas. Animals, 14(8), 1153. https://doi.org/10.3390/ani14081153