

Grain Type Impacts Feed Intake, Milk Production and Body Temperature of Dairy Cows Exposed to an Acute Heat Event in Early Lactation

 , , , ,
, , , , 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Experimental Design and Heat-Challenge Schedule
2.3. Feeding Regime, Feed Intake and Feed Composition
2.4. Milk Production
2.5. Physiology
2.6. Blood
2.7. Animal Welfare
2.8. Statistical Analyses
3. Results
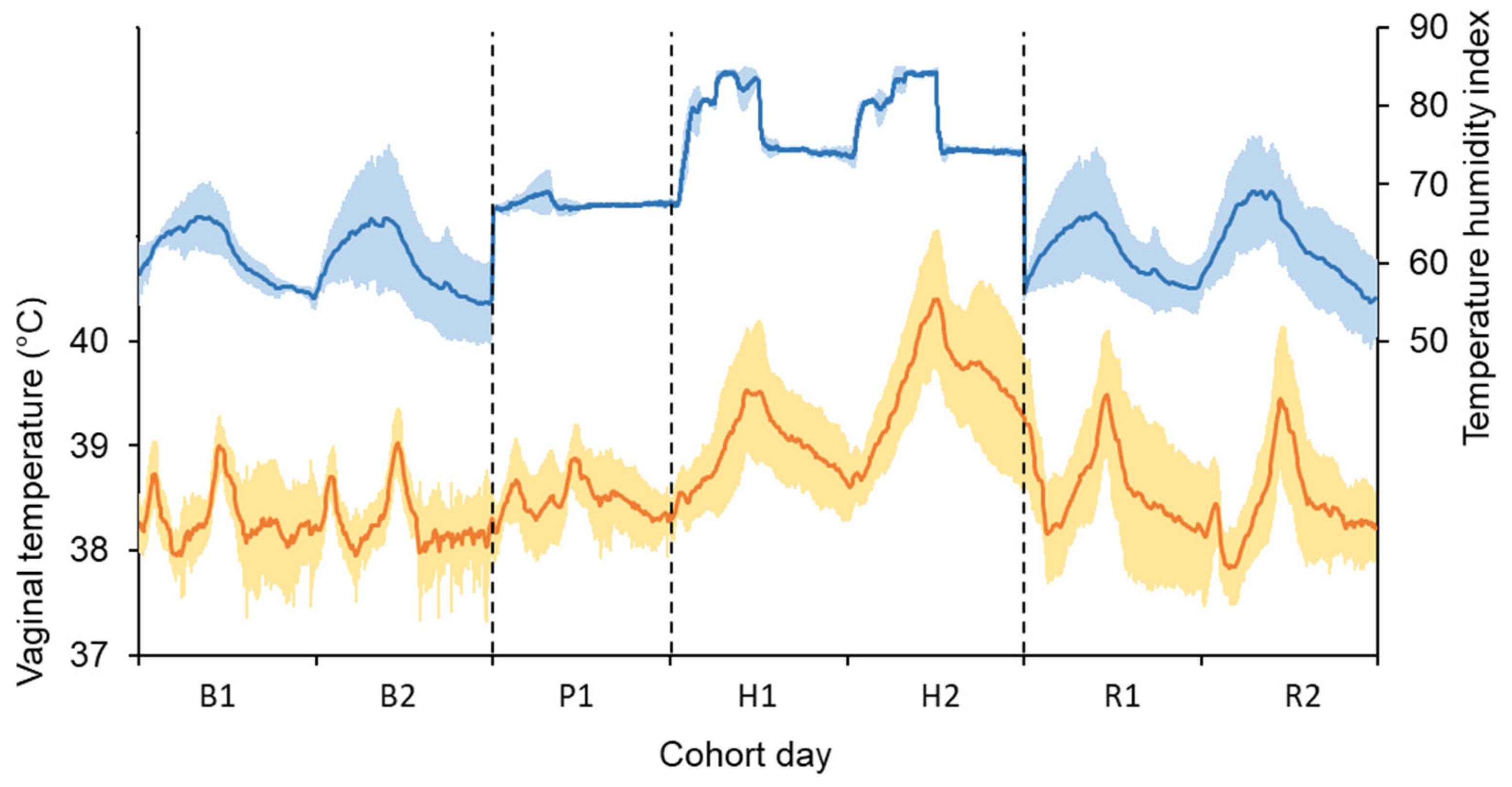
3.1. Conditions Experienced During the Experiment
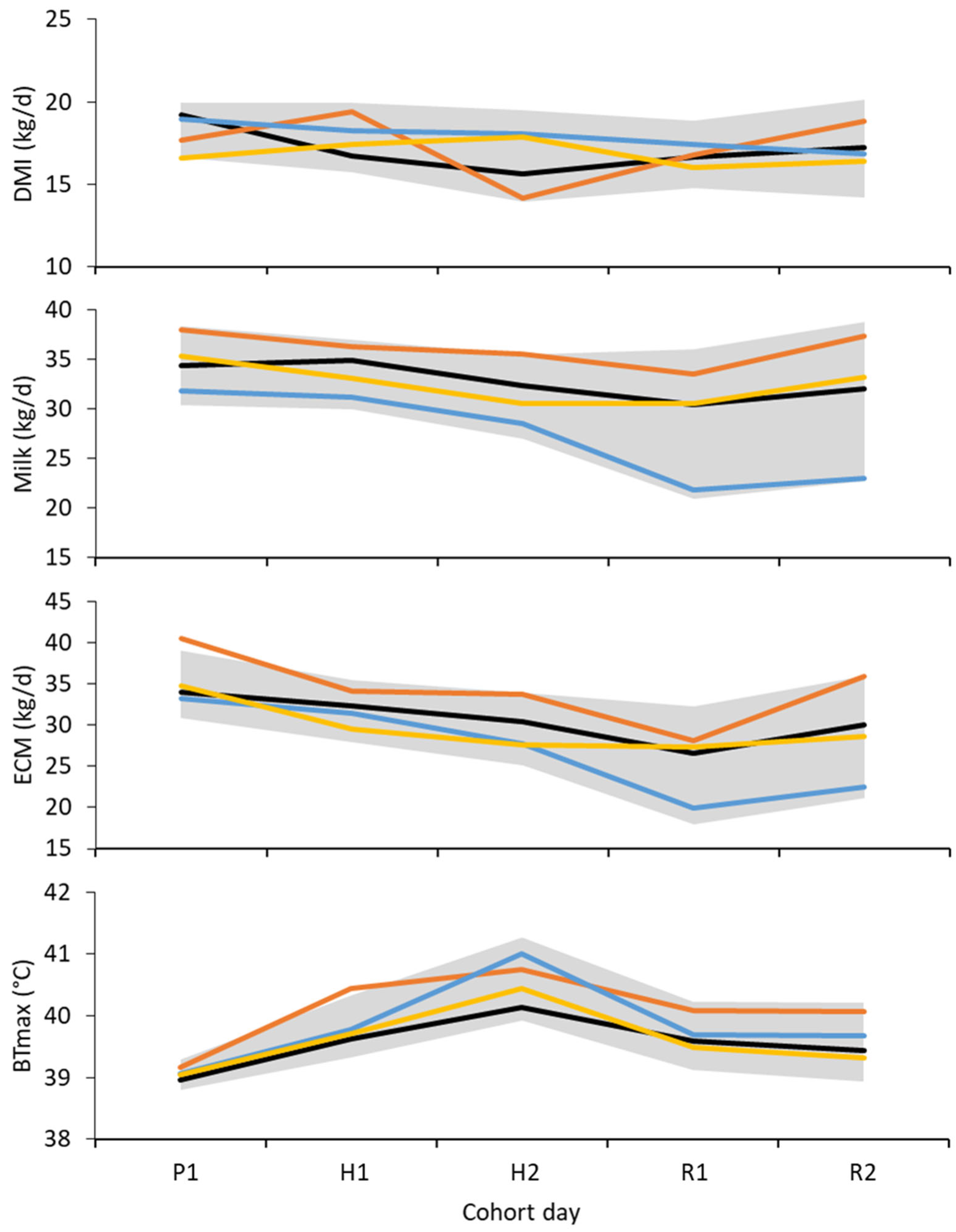
3.2. Dry Matter Intake
3.3. Milk Yield
3.4. Physiology
3.5. Blood
4. Discussion
4.1. Canola
4.2. Barley, Corn and Wheat
4.3. Dietary Heat
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Auldist, M.; Marett, L.; Greenwood, J.; Hannah, M.; Jacobs, J.; Wales, W. Effects of different strategies for feeding supplements on milk production responses in cows grazing a restricted pasture allowance. J. Dairy Sci. 2013, 96, 1218–1231. [Google Scholar] [PubMed]
- Smith, P.; Martino, D.; Cai, Z.; Gwary, D.; Janzen, H.; Kumar, P.; McCarl, B.; Ogle, S.; O’Mara, F.; Rice, C.; et al. Agriculture; Cambridge University: Cambridge, UK, 2007. [Google Scholar]
- Bureau of Meteorology. State of the Climate 2020; Bureau of Meteorology: Canberra, Australia, 2020.
- Silanikove, N. Effects of heat stress on the welfare of extensively managed domestic ruminants. Livest. Prod. Sci. 2000, 67, 1–18. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [PubMed]
- Baumgard, L.; Rhoads, R. Ruminant nutrition symposium: Ruminant production and metabolic responses to heat stress. J. Anim. Sci. 2012, 90, 1855–1865. [Google Scholar]
- Coppock, C. Energy nutrition and metabolism of the lactating dairy cow. J. Dairy Sci. 1985, 68, 3403–3410. [Google Scholar]
- Conrad, H.; Weiss, W.; Odwongo, W.; Shockey, W. Estimating net energy lactation from components of cell solubles and cell walls. J. Dairy Sci. 1984, 67, 427–436. [Google Scholar] [CrossRef]
- Allen, M.S.; Bradford, B.J.; Harvatine, K.J. The cow as a model to study food intake regulation. Annu. Rev. Nutr. 2005, 25, 523–547. [Google Scholar]
- Coppock, C.; Wilks, D. Supplemental fat in high-energy rations for lactating cows: Effects on intake, digestion, milk yield, and composition. J. Anim. Sci. 1991, 69, 3826–3837. [Google Scholar]
- Williams, S.R.O.; Milner, T.C.; Garner, J.B.; Moate, P.J.; Jacobs, J.L.; Hannah, M.C.; Wales, W.J.; Marett, L.C. Dietary Fat and Betaine Supplements Offered to Lactating Cows Affect Dry Matter Intake, Milk Production and Body Temperature Responses to an Acute Heat Challenge. Animals 2021, 11, 3110. [Google Scholar] [CrossRef]
- Garner, J.B.; Williams, S.R.O.; Moate, P.J.; Jacobs, J.L.; Hannah, M.C.; Morris, G.L.; Wales, W.J.; Marett, L.C. Effects of heat stress in dairy cows offered diets containing either wheat or corn grain during late lactation. Animals 2022, 12, 2031. [Google Scholar] [CrossRef]
- Hou, Y.; Zhang, L.; Dong, R.Y.; Liang, M.Y.; Lu, Y.; Sun, X.Q.; Zhao, X. Comparing responses of dairy cows to short-term and long-term heat stress in climate-controlled chambers. J. Dairy Sci. 2021, 104, 2346–2356. [Google Scholar] [CrossRef] [PubMed]
- Moe, P. Energy metabolism of dairy cattle. J. Dairy Sci. 1981, 64, 1120–1139. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, R.; Allison, M. Rumen metabolism. J. Anim. Sci. 1983, 57, 461–477. [Google Scholar] [PubMed]
- Murphy, M.R.; Baldwin, R.L.; Koong, L.J. Estimation of Stoichiometric Parameters for Rumen Fermentation of Roughage and Concentrate Diets. J. Anim. Sci. 1982, 55, 411–421. [Google Scholar] [CrossRef]
- Erickson, P.S.; Kalscheur, K.F. Nutrition and feeding of dairy cattle. In Animal Agriculture; Elsevier: Amsterdam, The Netherlands, 2020; pp. 157–180. [Google Scholar]
- Eastridge, M.L. Major Advances in Applied Dairy Cattle Nutrition. J. Dairy Sci. 2006, 89, 1311–1323. [Google Scholar] [CrossRef]
- Auldist, M.J.; Marett, L.C.; Greenwood, J.S.; Wright, M.M.; Hannah, M.; Jacobs, J.L.; Wales, W.J. Replacing wheat with canola meal in a partial mixed ration increases the milk production of cows grazing at a restricted pasture allowance in spring. Anim. Prod. Sci. 2014, 54, 869–878. [Google Scholar] [CrossRef]
- West, J. Effects of Heat-Stress on Production in Dairy Cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Gonzalez-Rivas, P.A.; Sullivan, M.; Cottrell, J.J.; Leury, B.J.; Gaughan, J.B.; Dunshea, F.R. Effect of feeding slowly fermentable grains on productive variables and amelioration of heat stress in lactating dairy cows in a sub-tropical summer. Trop. Anim. Health Prod. 2018, 50, 1763–1769. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes, 8th ed.; National Health and Medical Research Council: Canberra, Australia, 2013. [Google Scholar]
- NRC. Nutrient Requirements of Dairy Cattle: Seventh Revised Edition; National Academies Press: Washington, DC, USA, 2001. [Google Scholar] [CrossRef]
- Harville, D.A. Nearly optimal allocation of experimental units using observed covariate values. Technometrics 1974, 16, 589–599. [Google Scholar] [CrossRef]
- Williams, S.; Moate, P.; Hannah, M.; Ribaux, B.; Wales, W.; Eckard, R. Background matters with the SF6 tracer method for estimating enteric methane emissions from dairy cows: A critical evaluation of the SF6 procedure. Anim. Feed Sci. Technol. 2011, 170, 265–276. [Google Scholar] [CrossRef]
- Garner, J.B.; Douglas, M.L.; Williams, S.R.O.; Wales, W.J.; Marett, L.C.; Nguyen, T.T.T.; Reich, C.M.; Hayes, B.J. Genomic selection improves heat tolerance in dairy cattle. Sci. Rep. 2016, 6, 34114. [Google Scholar] [CrossRef]
- Kibler, H.H. Environmental Physiology and Shelter Engineering with Special Reference to Domestic Animals. LXVII, Thermal Effects of Various Temperature-Humidity Combinations on Holstein Cattle as Measured by Eight Physiological Responses; University of Missouri: Columbia, MO, USA, 1964. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 17th ed.; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000. [Google Scholar]
- Dairy One. Analytical Procedures. Available online: https://dairyone.com/download/forage-forage-lab-analytical-procedures/ (accessed on 7 October 2019).
- Weiss, W.P.; Conrad, H.R.; St. Pierre, N.R. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- Free, M.; Dove, H.; Nolan, J.V. (Eds.) Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Collingwood, VIC, Australia, 2007; p. 270. [Google Scholar]
- Tyrrell, H.F.; Reid, J.T. Prediction of the Energy Value of Cow’s Milk. J. Dairy Sci. 1965, 48, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Gaughan, J.B.; Mader, T.L.; Holt, S.M.; Lisle, A. A New Heat Load Index for Feedlot Cattle. J. Anim. Sci. 2008, 86, 226–234. [Google Scholar] [CrossRef]
- Cooke, R.F.; Arthington, J.D. Concentrations of haptoglobin in bovine plasma determined by ELISA or a colorimetric method based on peroxidase activity. J. Anim. Physiol. Anim. Nutr. 2013, 97, 531–536. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Moate, P.J.; Garner, J.B.; Hannah, M.C.; Giri, K.; Wales, W.J.; Marett, L.C. Dairy Cows Offered Fresh Chicory Instead of Ensiled Pasture during an Acute Heat Challenge Produced More Milk and Had Lower Body Temperatures. Animals 2023, 13, 867. [Google Scholar] [CrossRef]
- Saville, D.J. Multiple Comparison Procedures—Cutting the Gordian Knot. Agron. J. 2015, 107, 730–735. [Google Scholar] [CrossRef]
- Reed, K.F.; Bonfá, H.C.; Dijkstra, J.; Casper, D.P.; Kebreab, E. Estimating the energetic cost of feeding excess dietary nitrogen to dairy cows. J. Dairy Sci. 2017, 100, 7116–7126. [Google Scholar] [CrossRef]
- Moran, J.B. Cereal grains in complete diets for dairy cows: A comparison of rolled barley, wheat and oats and of three methods of processing oats. Anim. Prod. 1986, 43, 27–36. [Google Scholar] [CrossRef]
- Jeffery, H.; Buesnel, R.; O’Neill, G. Short-term studies on the response of grazing dairy cows to dietary supplementation. Aust. J. Exper. Agric. 1976, 16, 445–451. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Rode, L.M. Minimum Versus Optimum Concentrations of Fiber in Dairy Cow Diets Based on Barley Silage and Concentrates of Barley or Corn. J. Dairy Sci. 1997, 80, 1629–1639. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A.; Rode, L.M.; Yang, W.Z. Effects of Nonstructural Carbohydrates and Source of Cereal Grain in High Concentrate Diets of Dairy Cows. J. Dairy Sci. 1997, 80, 1640–1650. [Google Scholar] [CrossRef] [PubMed]
- Kargar, S.; Ghorbani, G.R.; Khorvash, M.; Sadeghi-Sefidmazgi, A.; Schingoethe, D.J. Reciprocal combinations of barley and corn grains in oil-supplemented diets: Feeding behavior and milk yield of lactating cows. J. Dairy Sci. 2014, 97, 7001–7011. [Google Scholar] [CrossRef] [PubMed]
- Moate, P.J.; Williams, S.R.O.; Deighton, M.H.; Hannah, M.C.; Ribaux, B.E.; Morris, G.L.; Jacobs, J.L.; Hill, J.; Wales, W.J. Effects of feeding wheat or corn and of rumen fistulation on milk production and methane emissions of dairy cows. Anim. Prod. Sci. 2019, 59, 891–905. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Cammell, S.B.; Humphries, D.J.; Beever, D.E.; Sutton, J.D.; Newbold, J.R. Effects of Postrumen Starch Infusion on Milk Production and Energy Metabolism in Dairy Cows. J. Dairy Sci. 2001, 84, 2250–2259. [Google Scholar] [CrossRef]
- Offner, A.; Bach, A.; Sauvant, D. Quantitative review of in situ starch degradation in the rumen. Anim. Feed Sci. Technol. 2003, 106, 81–93. [Google Scholar] [CrossRef]
- Tamminga, S.; van Vuuren, A.M.; van der Koelen, C.J.; Ketelaar, R.S.; van der Togt, P.L. Ruminal behaviour of structural carbohydrates, non-structural carbohydrates and crude protein from concentrate ingredients in dairy cows. Neth. J. Agric. Sci. 1990, 38, 513–526. [Google Scholar] [CrossRef]
- Gengler, W.R.; Martz, F.A.; Johnson, H.D.; Krause, G.F.; Hahn, L. Effect of Temperature on Food and Water Intake and Rumen Fermentation. J. Dairy Sci. 1970, 53, 434–437. [Google Scholar] [CrossRef]
- Bhattacharya, A.N.; Warner, R.G. Influence of Varying Rumen Temperature on Central Cooling or Warming and on Regulation of Voluntary Feed Intake in Dairy Cattle. J. Dairy Sci. 1968, 51, 1481–1489. [Google Scholar] [CrossRef]
- Gonzalez-Rivas, P.A.; DiGiacomo, K.; Russo, V.M.; Leury, B.J.; Cottrell, J.J.; Dunshea, F.R. Feeding slowly fermentable grains has the potential to ameliorate heat stress in grain-fed wethers. J. Anim. Sci. 2016, 94, 2981–2991. [Google Scholar] [CrossRef]
- Gonzalez-Rivas, P.A.; Digiacomo, K.; Giraldo, P.A.; Leury, B.J.; Cottrell, J.J.; Dunshea, F.R. Reducing rumen starch fermentation of wheat with three percent sodium hydroxide has the potential to ameliorate the effect of heat stress in grain-fed wethers. J. Anim. Sci. 2017, 95, 5547–5562. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Saldana, R.E.; Huber, J.T.; Poore, M.H. Dry Matter, Crude Protein, and Starch Degradability of Five Cereal Grains. J. Dairy Sci. 1990, 73, 2386–2393. [Google Scholar]
- Plaizier, J.C.; Krause, D.O.; Gozho, G.N.; McBride, B.W. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. Vet. J. 2008, 176, 21–31. [Google Scholar] [PubMed]
- Russo, V.M.; Leury, B.J.; Kennedy, E.; Hannah, M.C.; Auldist, M.J.; Wales, W.J. Forage type influences milk yield and ruminal responses to wheat adaptation in late-lactation dairy cows. J. Dairy Sci. 2018, 101, 9901–9914. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Chamber 1 | Chamber 2 | Chamber 3 | Chamber 4 | Chamber 5 | Chamber 6 | |
|---|---|---|---|---|---|---|
| Cohort 1 | WHT | BLY | CAN | WHT | CRN | CAN |
| Cohort 2 | CRN | CAN | BLY | BLY | WHT | WHT |
| Cohort 3 | CAN | WHT | CRN | CRN | BLY | BLY |
| Cohort 4 | BLY | CRN | WHT | CAN | CAN | CRN |
| BLY 1 | CAN | CRN | WHT | |
|---|---|---|---|---|
| Crude protein | 154 | 184 | 144 | 159 |
| Soluble protein (% CP) | 39.2 | 41.0 | 39.7 | 41.8 |
| Acid detergent fiber | 236 | 241 | 226 | 227 |
| Neutral detergent fiber | 354 | 352 | 326 | 337 |
| Acid detergent lignin | 45.8 | 49.2 | 41.9 | 43.6 |
| Non-fiber carbohydrate | 392 | 364 | 428 | 410 |
| Starch | 211 | 184 | 263 | 237 |
| Crude fat | 28.8 | 28.3 | 35.4 | 27.4 |
| Ash | 72.6 | 72.9 | 66.6 | 68.0 |
| Total digestible nutrients | 672 | 669 | 698 | 683 |
| Calcium | 5.4 | 6.0 | 5.2 | 5.4 |
| Magnesium | 2.0 | 2.4 | 2.0 | 2.0 |
| Sodium | 2.3 | 2.3 | 2.3 | 2.3 |
| Potassium | 20.1 | 20.6 | 19.7 | 19.7 |
| Chloride | 7.8 | 7.6 | 7.5 | 7.6 |
| DCAD 2 (meq./100 g DM) | 24.0 | 22.1 | 24.1 | 23.0 |
| Copper (mg/kg DM) | 7.6 | 8.7 | 7.2 | 8.8 |
| Sulfur | 2.5 | 3.1 | 2.4 | 2.6 |
| Metabolizable energy (ME, MJ/kg DM) | 11.3 | 11.4 | 11.5 | 11.5 |
| BLY 1 | CAN | CRN | WHT | SED | |
|---|---|---|---|---|---|
| Animals (n) | 5 | 3 | 6 | 4 | |
| 5-day mean | |||||
| Dry Matter Intake | 18.2 b | 16.2 a | 17.9 b | 17.0 ab | 0.701 |
| Crude Protein | 2.02 ab | 2.26 b | 1.83 a | 1.95 a | 0.134 |
| Neutral Detergent Fiber | 2.97 ab | 3.27 ab | 3.32 b | 2.55 a | 0.409 |
| Non-Fiber Carbohydrate | 8.13 c | 3.99 a | 5.69 b | 7.05 c | 0.711 |
| Starch | 3.99 b | 3.26 a | 5.02 d | 4.51 c | 0.186 |
| Fat | 0.24 a | 0.25 a | 0.41 b | 0.26 a | 0.046 |
| Metabolizable Energy (MJ/d) | 166 b | 147 a | 165 b | 158 ab | 6.396 |
| Pre-challenge | |||||
| Dry Matter Intake | 20.5 b | 16.9 a | 18.8 ab | 16.6 a | 1.25 |
| Crude Protein | 2.34 ab | 2.46 b | 2.02 a | 1.94 a | 0.215 |
| Neutral Detergent Fiber | 3.68 b | 3.62 ab | 3.80 b | 2.74 a | 0.537 |
| Non-Fiber Carbohydrate | 8.91 c | 4.38 a | 5.87 c | 6.72 b | 0.830 |
| Starch | 4.28 b | 3.34 a | 4.91 c | 4.21 b | 0.292 |
| Fat | 0.29 a | 0.28 a | 0.43 b | 0.26 a | 0.056 |
| Metabolizable Energy (MJ/d) | 184 b | 154 a | 174 ab | 155 a | 11.4 |
| Heat challenge | |||||
| Dry Matter Intake | 17.3 ab | 15.4 a | 18.0 b | 17.7 b | 0.931 |
| Crude Protein | 1.89 | 2.10 | 1.86 | 2.05 | 0.167 |
| Neutral Detergent Fiber | 2.80 | 3.14 | 3.42 | 2.88 | 0.457 |
| Non-Fiber Carbohydrate | 7.66 c | 3.53 a | 5.68 b | 7.17 c | 0.753 |
| Starch | 3.66 b | 2.96 a | 4.93 d | 4.47 c | 0.229 |
| Fat | 0.21 a | 0.23 a | 0.40 b | 0.27 a | 0.049 |
| Metabolizable Energy (MJ/d) | 158 ab | 141 a | 167 b | 164 b | 8.51 |
| Recovery | |||||
| Dry Matter Intake | 18.1 | 16.7 | 17.3 | 16.6 | 0.929 |
| Crude Protein | 1.99 ab | 2.33 b | 1.70 a | 1.86 a | 0.161 |
| Neutral Detergent Fiber | 2.80 | 3.14 | 3.42 | 2.88 | 0.443 |
| Non-Fiber Carbohydrate | 8.21 c | 4.26 a | 5.62 b | 7.09 c | 0.732 |
| Starch | 4.16 b | 3.52 a | 5.16 d | 4.70 c | 0.237 |
| Fat | 0.25 a | 0.26 a | 0.40 b | 0.24 a | 0.0473 |
| Metabolizable Energy (MJ/d) | 165 | 151 | 159 | 154 | 8.512 |
| Pre-challenge to heat 2 | |||||
| Δ Dry Matter Intake | −2.8 a | −1.5 ab | −0.8 ab | 1.1 b | 1.190 |
| Δ Crude Protein | −0.45 a | −0.36 ab | −0.16 ab | 0.11 b | 0.189 |
| Δ Neutral Detergent Fiber | −0.87 a | −0.48 ab | −0.38 ab | 0.14 b | 0.395 |
| Δ Non-Fiber Carbohydrate | −1.25 a | −0.84 ab | −0.19 bc | 0.45 c | 0.512 |
| Δ Starch | −0.62 a | −0.38 ab | 0.02 b | 0.26 b | 0.297 |
| Δ Fat | −0.08 a | −0.05 ab | −0.03 ab | 0.01 b | 0.037 |
| Δ Metabolizable Energy (MJ/d) | −26 a | −13 ab | −6 ab | 9 b | 10.8 |
| BLY 1 | CAN | CRN | WHT | SED | |
|---|---|---|---|---|---|
| Animals (n) | 5 | 3 | 6 | 4 | |
| 5-day mean | |||||
| Milk yield | 31.7 ab | 35.2 b | 31.4 a | 32.7 ab | 1.52 |
| ECM 2 yield | 29.5 a | 34.9 b | 29.2 a | 30.2 a | 1.58 |
| Fat yield | 1.16 a | 1.44 b | 1.21 a | 1.18 a | 0.083 |
| Protein yield | 0.88 a | 1.06 b | 0.83 a | 0.91 a | 0.053 |
| Fat concentration | 41.0 | 37.2 | 40.7 | 35.7 | 6.34 |
| Protein concentration | 29.8 b | 27.6 ab | 28.8 ab | 27.5 a | 1.33 |
| Pre-challenge | |||||
| Milk yield | 33.3 | 37.0 | 33.7 | 35.6 | 1.90 |
| ECM yield | 32.6 a | 41.3 b | 33.6 a | 34.5 a | 2.22 |
| Fat yield | 1.32 a | 1.81 b | 1.48 a | 1.47 a | 0.135 |
| Protein yield | 1.01 a | 1.22 b | 0.92 a | 1.05 b | 0.076 |
| Fat concentration | 43.4 | 45.1 | 46.2 | 41.0 | 7.17 |
| Protein concentration | 31.4 | 31.2 | 29.2 | 29.1 | 1.50 |
| Heat challenge | |||||
| Milk yield | 32.5 | 34.9 | 32.5 | 32.0 | 1.69 |
| ECM yield | 30.1 ab | 34.5 b | 30.7 ab | 29.2 a | 1.84 |
| Fat yield | 1.18 ab | 1.42 b | 1.30 ab | 1.13 a | 0.107 |
| Protein yield | 0.92 ab | 1.04 b | 0.88 a | 0.88 a | 0.064 |
| Fat concentration | 40.1 ab | 36.7 ab | 41.8 b | 35.2 a | 6.86 |
| Protein concentration | 29.7 | 27.1 | 29.0 | 27.2 | 1.39 |
| Recovery | |||||
| Milk yield | 30.2 a | 34.5 b | 29.1 a | 31.9 ab | 1.70 |
| ECM yield | 27.2 a | 32.2 b | 25.5 a | 28.5 ab | 1.87 |
| Fat yield | 1.06 ab | 1.28 b | 0.98 a | 1.08 ab | 0.108 |
| Protein yield | 0.78 ab | 0.98 c | 0.72 a | 0.87 bc | 0.064 |
| Fat concentration | 40.6 | 33.7 | 36.7 | 33.5 | 6.88 |
| Protein concentration | 29.1 | 26.2 | 28.3 | 27.0 | 1.39 |
| Before challenge with heat 3 | |||||
| Δ Milk yield | −0.84 | −2.10 | −1.23 | −3.52 | 1.523 |
| Δ ECM yield | −2.47 | −6.81 | −2.85 | −6.24 | 2.153 |
| Δ Fat yield | −0.14 | −0.39 | −0.19 | −0.34 | 0.142 |
| Δ Protein yield | −0.09 | −0.19 | −0.05 | −0.17 | 0.075 |
| Δ Fat concentration | −3.38 | −8.39 | −4.37 | −5.88 | 3.286 |
| Δ Protein concentration | −1.71 b | −4.06 a | −0.18 b | −1.88 ab | 1.003 |
| BLY 1 | CAN | CRN | WHT | SED | |
|---|---|---|---|---|---|
| Animals (n) | 5 | 3 | 6 | 4 | |
| 5-day mean | |||||
| Mean | 38.4 a | 38.9 b | 38.9 b | 38.7 ab | 0.20 |
| Minimum | 37.74 a | 38.07 ab | 38.14 b | 37.91 ab | 0.142 |
| Maximum | 39.3 a | 40.0 b | 39.9 b | 39.5 ab | 0.29 |
| Duration > 38.8 °C | 318 a | 675 b | 625 b | 433 ab | 141 |
| Respiration | 65.6 | 78.3 | 65.2 | 70.2 | 12.70 |
| Skin temperature | 33.5 a | 35.6 b | 35.2 b | 34.3 ab | 0.75 |
| Pre-challenge | |||||
| Mean | 38.2 | 38.6 | 38.7 | 38.4 | 0.24 |
| Minimum | 37.8 a | 38.0 ab | 38.2 b | 37.8 a | 0.19 |
| Maximum | 38.7 | 39.2 | 39.2 | 38.9 | 0.34 |
| Duration > 38.8 °C | 12 | 404 | 251 | 113 | 191 |
| Respiration | 39.2 | 45.2 | 42.4 | 43.9 | 14.50 |
| Skin temperature | 31.2 a | 33.0 ab | 34.2 b | 32.5 ab | 1.43 |
| Heat challenge | |||||
| Mean | 38.9 a | 39.3 ab | 39.5 b | 39.2 ab | 0.22 |
| Minimum | 38.1 a | 38.3 ab | 38.5 b | 38.3 ab | 0.16 |
| Maximum | 39.7 a | 40.5 b | 40.4 b | 39.9 ab | 0.31 |
| Duration > 38.8 °C | 777 a | 1068 ab | 1157 b | 934 ab | 160 |
| Respiration | 93.8 | 97.1 | 91.0 | 101 | 13.48 |
| Skin temperature | 35.6 | 35.8 | 36.7 | 35.6 | 1.03 |
| Recovery | |||||
| Mean | 38.0 a | 38.7 b | 38.5 ab | 38.3 a | 0.22 |
| Minimum | 37.4 a | 38.0 b | 37.7 ab | 37.6 a | 0.17 |
| Maximum | 39.2 a | 40.0 b | 39.7 ab | 39.3 a | 0.31 |
| Duration > 38.8 °C | 14 a | 417 b | 281 ab | 93 ab | 161 |
| Respiration | 50.7 | 76.0 | 50.8 | 52.9 | 13.45 |
| Skin temperature | 32.5 a | 36.6 b | 34.3 ab | 33.9 ab | 1.41 |
| Before challenge with heat 2 | |||||
| Δ Mean | 0.70 | 0.67 | 0.81 | 0.80 | 0.165 |
| Δ Minimum | 0.28 | 0.25 | 0.27 | 0.45 | 0.158 |
| Δ Maximum | 1.04 | 1.26 | 1.15 | 1.01 | 0.251 |
| Δ Duration > 38.8 °C | 764 | 664 | 906 | 820 | 182 |
| Δ Respiration | 54.6 | 51.8 | 48.6 | 56.8 | 9.63 |
| Δ Skin temperature | 4.4 | 2.8 | 2.4 | 3.1 | 1.76 |
| BLY 1 | CAN | CRN | WHT | SED | |
|---|---|---|---|---|---|
| Animals (n) | 5 | 3 | 6 | 4 | |
| Pre-challenge | |||||
| pH | 7.11 | 7.13 | 7.18 | 7.15 | 0.055 |
| BHB | 0.97 | 0.60 | 0.93 | 0.53 | 0.257 |
| NEFA | 0.17 | 0.13 | 0.11 | 0.08 | 0.061 |
| Glucose | 3.08 | 2.98 | 3.02 | 3.39 | 0.287 |
| Haptoglobin | 0.25 b | 0.03 a | 0.17 ab | 0.13 ab | 0.097 |
| Na | 138 ab | 139 ab | 135 a | 140 b | 2.2 |
| Heat challenge | |||||
| pH | 7.15 | 7.12 | 7.14 | 7.14 | 0.054 |
| BHB | 0.60 | 0.71 | 0.63 | 0.44 | 0.257 |
| NEFA | 0.32 b | 0.46 c | 0.19 a | 0.17 a | 0.061 |
| Glucose | 3.26 | 3.13 | 3.43 | 3.41 | 0.287 |
| Haptoglobin | 0.17 | 0.03 | 0.16 | 0.12 | 0.097 |
| Na | 137 | 135 | 135 | 133 | 2.1 |
| Recovery | |||||
| pH | 7.23 | 7.16 | 7.21 | 7.26 | 0.061 |
| BHB | 0.77 | 0.73 | 0.80 | 0.42 | 0.253 |
| NEFA | 0.09 | 0.16 | 0.15 | 0.06 | 0.060 |
| Glucose | 2.93 | 2.90 | 2.87 | 3.07 | 0.283 |
| Haptoglobin | 0.03 | 0.11 | 0.16 | 0.06 | 0.095 |
| Na | 133 | 135 | 134 | 134 | 2.4 |
| Before challenge with heat 2 | |||||
| Δ pH | 0.04 | −0.01 | −0.05 | −0.01 | 0.076 |
| Δ BHB | −0.37 | −0.11 | −0.30 | −0.08 | 0.274 |
| Δ NEFA | 0.15 a | 0.33 b | 0.08 a | 0.09 a | 0.078 |
| Δ Glucose | 0.18 | 0.16 | 0.40 | 0.01 | 0.336 |
| Δ Haptoglobin | −0.08 | 0.00 | −0.02 | −0.01 | 0.116 |
| Δ Na | −1.4 ab | −4.2 ab | 0.1 b | −6.4 a | 2.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, S.R.O.; Knight, M.I.; Milner, T.C.; Garner, J.B.; Moate, P.J.; Giri, K.; Hannah, M.C.; Jacobs, J.L.; Wales, W.J.; Marett, L.C. Grain Type Impacts Feed Intake, Milk Production and Body Temperature of Dairy Cows Exposed to an Acute Heat Event in Early Lactation. Animals 2025, 15, 1045. https://doi.org/10.3390/ani15071045
Williams SRO, Knight MI, Milner TC, Garner JB, Moate PJ, Giri K, Hannah MC, Jacobs JL, Wales WJ, Marett LC. Grain Type Impacts Feed Intake, Milk Production and Body Temperature of Dairy Cows Exposed to an Acute Heat Event in Early Lactation. Animals. 2025; 15(7):1045. https://doi.org/10.3390/ani15071045
Chicago/Turabian StyleWilliams, S. Richard O., Matthew I. Knight, Tori C. Milner, Josie B. Garner, Peter J. Moate, Khageswor Giri, Murray C. Hannah, Joe L. Jacobs, William J. Wales, and Leah C. Marett. 2025. "Grain Type Impacts Feed Intake, Milk Production and Body Temperature of Dairy Cows Exposed to an Acute Heat Event in Early Lactation" Animals 15, no. 7: 1045. https://doi.org/10.3390/ani15071045
APA StyleWilliams, S. R. O., Knight, M. I., Milner, T. C., Garner, J. B., Moate, P. J., Giri, K., Hannah, M. C., Jacobs, J. L., Wales, W. J., & Marett, L. C. (2025). Grain Type Impacts Feed Intake, Milk Production and Body Temperature of Dairy Cows Exposed to an Acute Heat Event in Early Lactation. Animals, 15(7), 1045. https://doi.org/10.3390/ani15071045