Simple Summary
Reducing reliance on antibiotic growth promoters (AGPs) is a priority to combat antimicrobial resistance (AMR). This study explores how temperature modification during egg incubation, a process called thermal manipulation (TM), can enhance broilers’ gut health and performance under lipopolysaccharide (LPS)-induced stress. Using 16S rRNA gene sequencing, we analyzed the gut microbiome and found that chicks exposed to TM had a different composition of gut bacteria, particularly by increasing beneficial bacterial taxa such as Oscillospirales. These microbial changes may improve the birds’ immune response to LPS-induced stress, as TM-treated birds avoided the hypothermic phase typically triggered by LPS. These findings suggest that TM could be a cost-effective and easily applicable way to improve poultry health, reduce antibiotic dependence, and support sustainable farming practices.
Abstract
Thermal manipulation (TM) during embryogenesis has emerged as a promising strategy to enhance post-hatch performance and improve resilience to environmental and bacterial stress, which offers a potential alternative to reduce the reliance on antibiotic growth promoters (AGPs) in broiler production. This study investigated TM’s ability to modulate broilers’ cecal microbiota and enhance resilience to lipopolysaccharide (LPS)-induced stress. Eggs in the control group (CON) were incubated at 37.8 °C and 56% relative humidity (RH), while TM eggs were exposed to 39 °C and 65% RH for 18 h daily from embryonic days 10–18. Post-hatch, the LPS subgroups (LPS-CON, LPS-TM) received intraperitoneal LPS injections, and body weight (BW) and temperature (BT) were monitored. Cecal samples were collected for microbiome sequencing. Alpha diversity showed no differences (p > 0.05), but beta diversity revealed differences between groups (PERMANOVA, p < 0.05). Firmicutes and Bacteroidota dominated the microbiota at the phylum level. Oscillospirales were enriched in the TM groups (p < 0.001) and Lactobacillales were increased in the LPS-CON group (p < 0.019). LPS reduced BT in the CON group (p < 0.01), but LPS-TM birds bypassed hypothermia. LPS significantly reduced BW (p < 0.001), while TM had no significant effect. These findings demonstrate TM’s enduring influence on gut microbiota and stress resilience, highlighting its potential to reduce antibiotic reliance and mitigate antimicrobial resistance (AMR) in poultry production.
1. Introduction
Chickens are essential for meeting the rising global demand for animal protein. Specifically, in 2021, chicken meat was the most produced meat worldwide, reaching approximately 120 million tons [1]. Artificial genetic selection is an essential strategy to improve the rate of meat production, resulting in a more than 400% increase in broiler growth rate over the past fifty years. Still, it has been linked with various disadvantages in the body, including negatively affected musculoskeletal, cardiac, and immune systems [2]. Additionally, it has been found to alter the gut microbiota, likely due to a modified mucosal ecological environment [3].
Antibiotic growth promoters (AGPs) have been widely used to enhance broiler performance, increasing growth rates by 4–8% and feed efficiency by 2–5% [4]. However, the broad use of AGPs leads to residues in meat and the development of antimicrobial resistance (AMR) [5]. By 2050, AMR is predicted to cause 10 million deaths annually if proper actions are not taken to slow down the threat [6]. Consequently, finding sustainable alternatives to AGPs has become a priority.
Thermal manipulation (TM) is a highly efficient and cost-effective method for improving broiler production and health, offering a promising alternative to reduce the reliance on AGPs. TM, achieved through epigenetic modifications by altering incubation temperature and humidity during the critical period of embryogenesis, has been shown to improve post-hatch performance [7,8]. Studies demonstrate that TM promotes muscle growth, strengthens resistance to bacterial challenges, and enhances immune responses under heat stress [9,10,11,12].
Lipopolysaccharide (LPS), a Gram-negative bacterial endotoxin, causes systemic inflammation and weight loss through appetite suppression and immunological pathway changes [13]. Previous research has demonstrated that TM influences immune responses by modulating splenic cytokine expression and heat shock protein (HSP) mRNA levels in LPS-challenged broiler chickens [14]. Similarly, Shanmugasundaram et al. [15] reported that TM in Pekin duck embryos significantly reduced bursal and splenic HSP70 and IL-6 mRNA levels following LPS challenge, supporting TM’s potential role in enhancing immune resilience.
The gut microbiome, composed of bacteria, fungi, archaea, protozoa, and viruses, plays a pivotal role in nutrient digestion, immunity, and growth performance in broilers [16]. TM may enhance gut microbiome resilience, potentially mitigating LPS-induced inflammation and supporting immune function. Recent studies have demonstrated significant microbial shifts during embryonic development; however, the effects of embryonic environmental conditions, such as TM, on the microbiome remain largely underexplored [17]. For instance, while TM has been shown to alter enterocyte proliferation and brush-border membrane enzyme activity—and when combined with post-hatch baicalein supplementation, it enhances volatile fatty acid production and improves gut health by promoting beneficial microbial communities—its influence on microbial communities during LPS-induced inflammation has not been thoroughly investigated [18,19]. Additionally, some studies have investigated post-hatch challenges like LPS-induced microbial change, reporting limited shifts in microbial diversity and increased pathogenic bacteria under LPS stress [20,21,22]. Research on the interaction between TM and post-hatch LPS challenge, as well as the impact on microbiome responses, remains limited.
This study investigates TM’s potential in modulating the cecal microbiome to enhance resilience against LPS-induced intestinal stress. By modifying embryonic development, TM may induce long-term shifts in gut microbial community composition and function. This approach could offer a sustainable alternative to AGPs to improve broiler health and performance.
2. Materials and Methods
The Animal Care and Use Committee of Jordan University of Science and Technology (Approval Number: 347/12/4/16) reviewed and approved all experimental procedures.
2.1. Study Population and Incubation
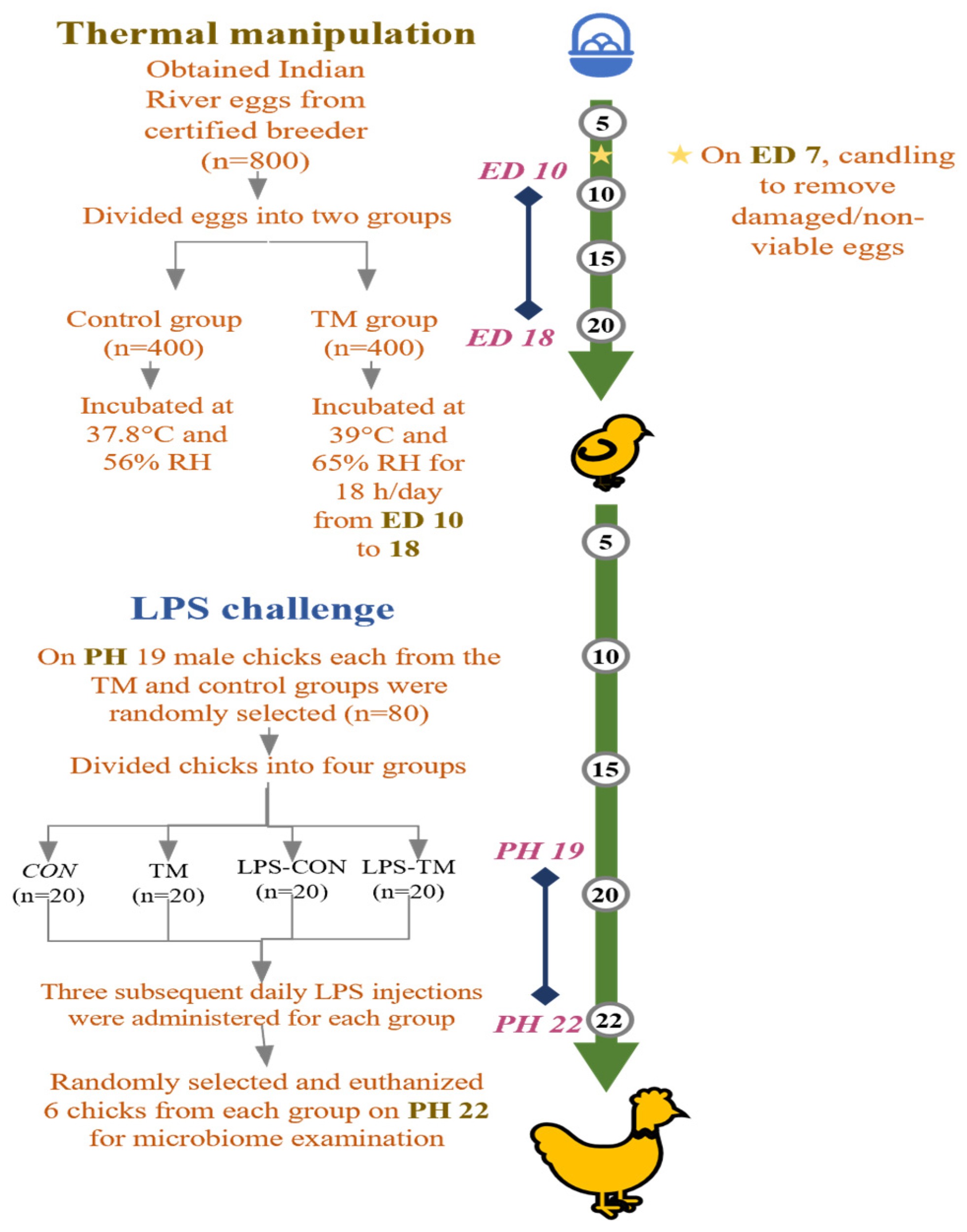
Figure 1 summarizes the experimental design used in the current study. After visual inspection to exclude damaged eggs, selected eggs (53.8 ± 3.49 g) were divided into two groups: the control (CON) and TM. The eggs in the CON group were kept at a regular 37.8 °C air temperature and 56% relative humidity (RH) throughout the embryogenesis period. The eggs in the TM group were incubated under regular incubation environments, except on embryonic days 10–18, when incubated at 39 °C and 65% RH for 18 h each day, consistently, from 12:00 a.m. to 8:00 a.m. All eggs were incubated in a Masalles Mod.25-I PDS incubator (Barcelona, Spain), with candling performed on embryonic day 7 to remove infertile eggs and those containing dead embryos.
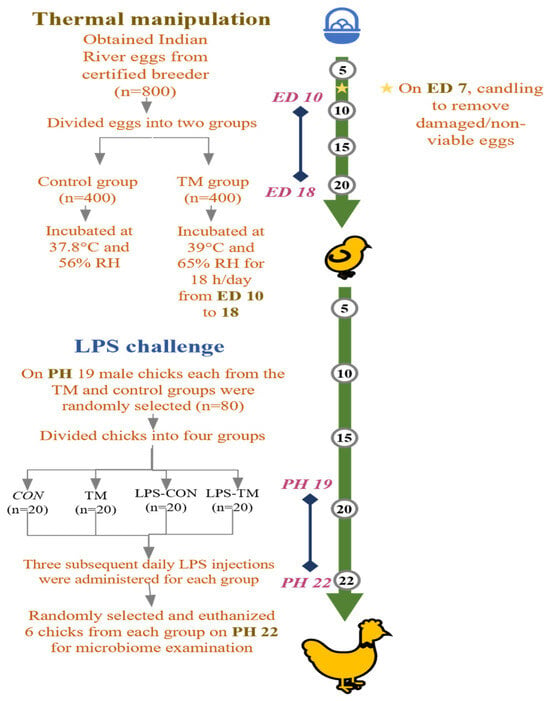
Figure 1.
Summary of the experimental design carried out in this study. TM: thermal manipulation, CON: control, RH: relative humidity, LPS: lipopolysaccharide, ED: embryonic day, PH: post-hatch.
According to our earlier research, using this TM procedure under heat stress in Cobb and Hubbard breeds results in changes to the expression of heat shock proteins and inflammatory cytokines, as well as a decrease in the expression of genes linked to oxidative stress and an increase in antioxidant capacity [23,24].
2.2. Hatching Management and Post-Hatching Rearing
Chicks were monitored hourly on the day of hatching. At 24 h post-hatching, the chicks were feather-sexed and then transferred to the animal house at Jordan University of Science and Technology, where the field experiments were conducted. The male broiler chicks were equally distributed into two groups. Each group contained 16 caged pens (80 × 40 × 100 cm). The pen was considered an experimental unit. Chicks were fed basal diets as per the National Research Council’s recommendations, fulfilling the standard nutrient requirements (Table 1), with the method adapted from Shakouri and Malekzadeh [25].

Table 1.
Experimental diet composition based on NRC (1994) guidelines.
Chicks received vaccination against Newcastle disease and infectious bronchitis. All chicks were raised under similar managerial practices. The experimental trial lasted 22 days, from 15 May to 5 June. Throughout the first week, the room temperature was kept at 33 ± 1 °C; it gradually decreased to 22 °C by the end of the third week. Throughout the field trial, the chicks received meal rations ad libitum and unlimited access to water [26].
2.3. LPS Challenge
On post-hatch day 19, the LPS challenge was administered. Both primary groups, TM and CON, were divided into two subgroups. The CON and TM subgroups did not have LPS injections; a second subgroup received the LPS treatment (LPS-CON and LPS-TM). On days 19, 20, and 21, the LPS subgroups (n = 20) received three intraperitoneal injections of LPS from E. coli O127 at a dose of 0.5 mg/kg. This dosing protocol, adapted from previous research by Wu et al. [27], demonstrated that subsequent LPS injections reliably induced intestinal mucosal damage, characterized by hemorrhagic lesions in the ileum and jejunum, severe villus structural damage (including epithelial fragmentation and mucosal erosion), and significant alterations in cecal microbial composition consistent with gut dysbiosis. Additionally, supporting this approach Zheng et al. [28] showed that three intraperitoneal LPS injections administered at 16, 18, and 20 days of age effectively induced oxidative stress, inflammatory responses, and intestinal morphological changes in broiler chicks.
After administering the injection, each subgroup was relocated to an experimental pen and maintained under thermoneutral conditions to ensure their physiological comfort. Eight hours post-injection, six new random chick samples were selected daily from each subgroup to measure body weight (BW) and temperature (BT). Additionally, eight hours after the last LPS injection, cecal content samples were collected for microbiome analysis. Chickens were humanely euthanized, and the ceca were aseptically excised using sterile instruments. The ceca were carefully and immediately snap-frozen on site to prevent DNA degradation. Samples were stored in a CryoCube® F570 Series Ultra-Low Temperature (ULT) Freezer (Eppendorf, Hamburg, Germany) at −80 °C to preserve microbial DNA integrity until extraction. A J/K/T thermocouple meter connected to a rat rectal probe (manufactured by Kent Scientific Corporation, Torrington, CT, USA; accuracy ±0.1 °C) was used to measure the chickens’ body temperature.
2.4. Microbiological Analysis
This study employed Illumina-based next-generation sequencing (NGS) to investigate the effects of embryonic TM on the cecum microbiome during a post-hatch LPS challenge. The Illumina platform, widely used for 16S rRNA gene sequencing, provides a highly accurate culture-independent approach for profiling microbial communities [29]. The 16S rRNA gene benefits this study due to its variable regions, which allow for the classification of microbial taxa. This method enables detailed insights into microbial diversity and community dynamics in environmental and host-associated samples.
2.4.1. DNA Isolation and Sequencing
Microbial DNA was isolated from about 250 mg of cecal contents collected from six randomly selected chicks per experimental subgroup using the DNeasy PowerSoil Pro Kit (QIAGEN, Inc., Valencia, CA, USA) according to the manufacturer’s procedures [30]. The use of 6 samples in this study was informed by recent findings in broiler chicken microbiome research. Weinroth and Oakley demonstrated that sampling 6–10 birds captures most microbial diversity, with less than 1% new amplicon sequencing variants (ASVs) added beyond this point [31].
The (V3-V4) region of the 16S rRNA gene was amplified using universal primers Bakt_341F (CCTACGGGNGGCWGCAG) and Bakt_805R (GACTACHVGGGTATCTAATC C). Macrogen Inc., Seoul, Republic of Korea, performed the sequencing commercially utilizing an Illumina MiSeq platform (Illumina, San Diego, CA, USA), following the 2 × 300 bp paired-end sequencing protocol.
2.4.2. Data Analysis by Bioinformatics Tools
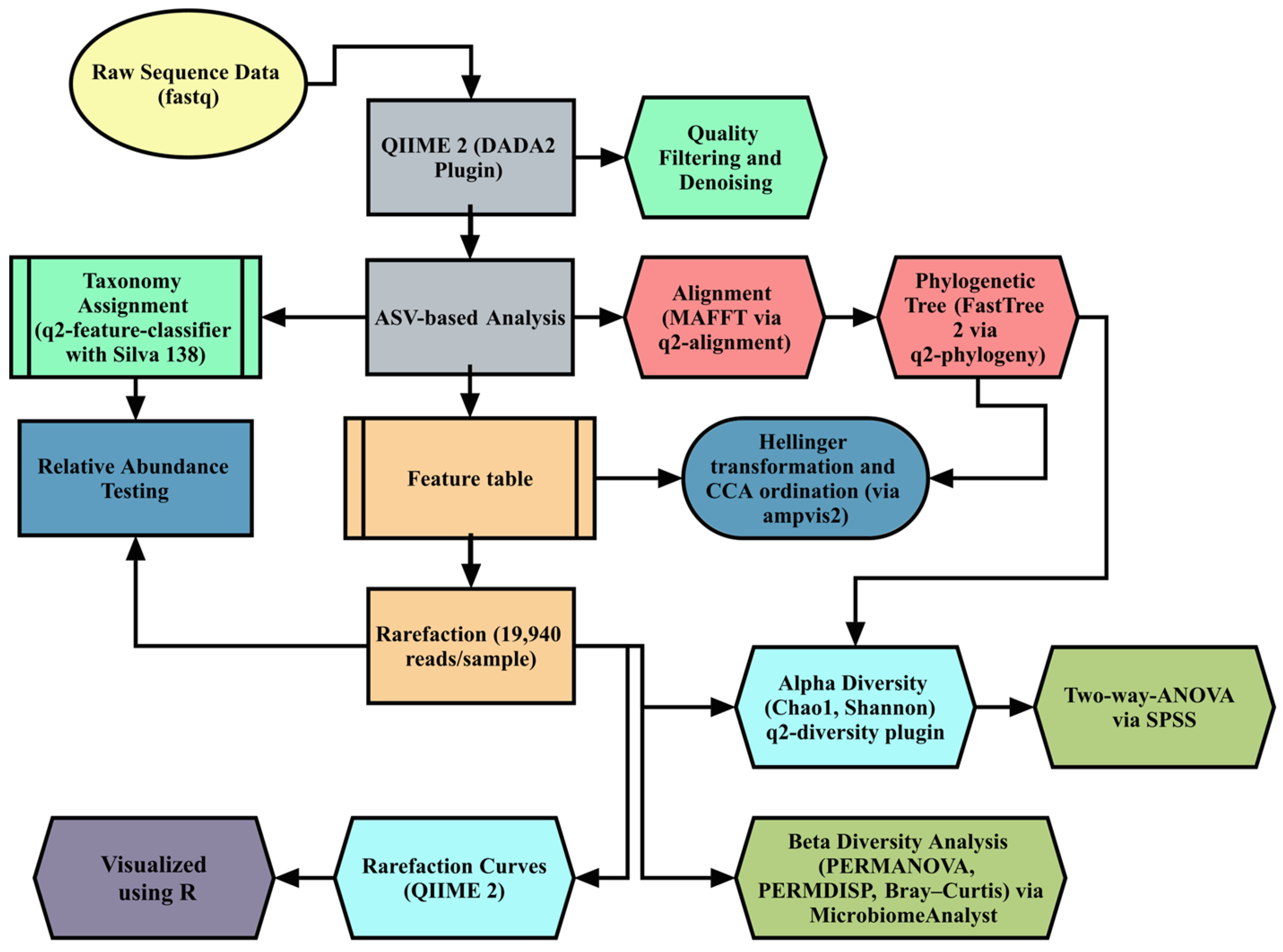
Figure 2 depicts the comprehensive pipeline utilized for the S16 microbiome sequencing of cecal content. QIIME 2 version 2024.8 was used for microbiome bioinformatics [32], using DADA2 for quality filtering, denoising, and amplicon sequence variant (ASV) generation. Taxonomic classification was performed with the Silva 138 reference database (99% OTUs). Rarefaction was performed with 19,940 reads per sample to standardize diversity metrics, including the Chao1, Shannon, and Bray–Curtis indices. Beta diversity was analyzed using Canonical Correspondence Analysis (CCA) on Hellinger-transformed data, implemented in the ampvis2 R package. Differences in microbial composition were tested using PERMANOVA (Permutational Multivariate Analysis of Variance) and validated by PERMDISP (permutational analysis of multivariate dispersions). Differential abundance analysis was conducted via LEfSe (LDA score > 2.0, p < 0.05) and STAMP for Kruskal–Wallis tests. The full bioinformatics workflow and references are detailed in Supplementary Materials S1.
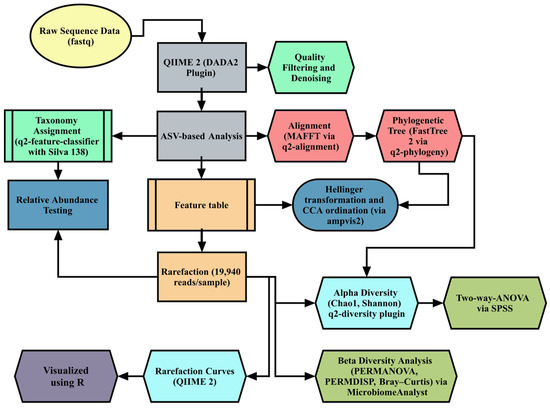
Figure 2.
Summary of the bioinformatics analysis carried out in this study. ASV: amplicon sequencing variant; CCA: canonical correspondence analysis; PERMANOVA: Permutational Multivariate Analysis of Variance; PERMDISP: permutational analysis of multivariate dispersions.
2.5. Statistical Analyses
BT and BW were analyzed using IBM SPSS Statistics 27.0 (IBM software, Chicago, IL, USA) and are represented as means ± SD. A 2 × 2 factorial design was employed, with random sampling for each measurement time point. A two-way ANOVA was used to compare variations in BT and BW within the same day between treatment groups. To analyze changes in BT over time, a one-way ANOVA was applied separately for each group (e.g., CON group: Day 1 vs. Day 2 vs. Day 3). In addition, BT was visualized using GraphPad Software, statistical version 10.0 (San Diego, CA, USA), while BW was visualized using Microsoft Excel. p values lower than 0.05 were considered significantly different.
3. Results
3.1. Effects of TM and LPS on Alpha Diversity
There were no significant differences (p > 0.05) in richness or diversity among the groups (Table 2), suggesting that neither TM nor LPS challenge had a direct impact on alpha diversity under the experimental conditions tested.
Table 2.
Alpha diversity indexes of microbial communities.
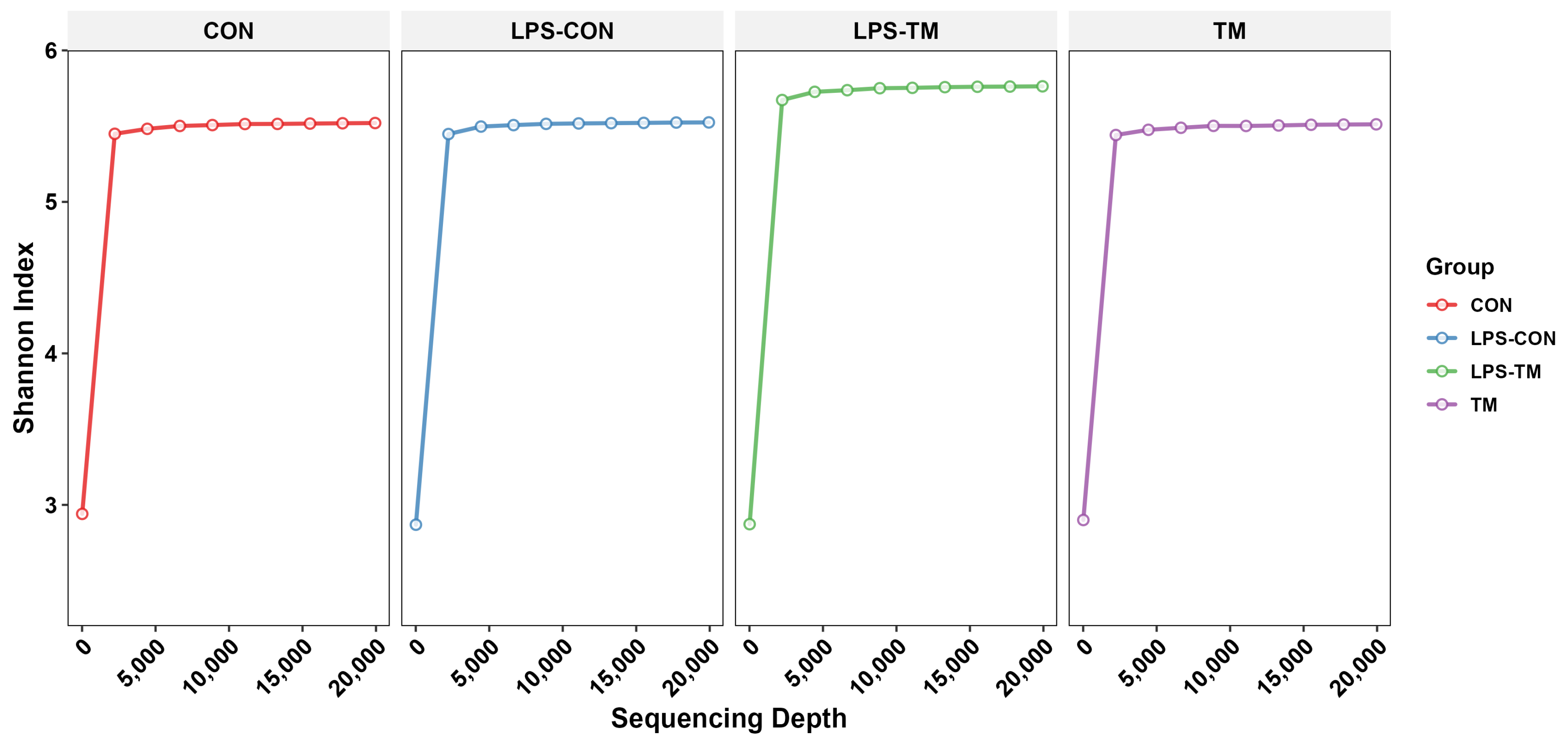
3.2. Rarefaction Curve Analysis
Figure 3 displays the rarefaction curves, which illustrate the Shannon index as a function of sequencing depth. The curves reach a plateau for all groups, indicating that sufficient sequencing depth was achieved to capture the diversity within each group.
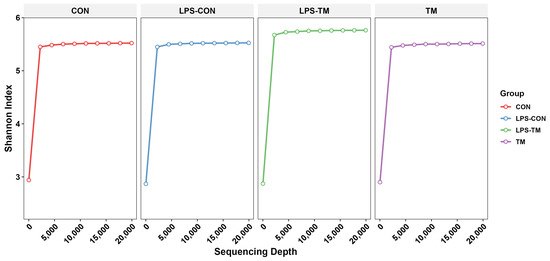
Figure 3.
The curves approach a plateau, indicating that the sequencing depth was sufficient to capture the most microbial diversity in all groups.
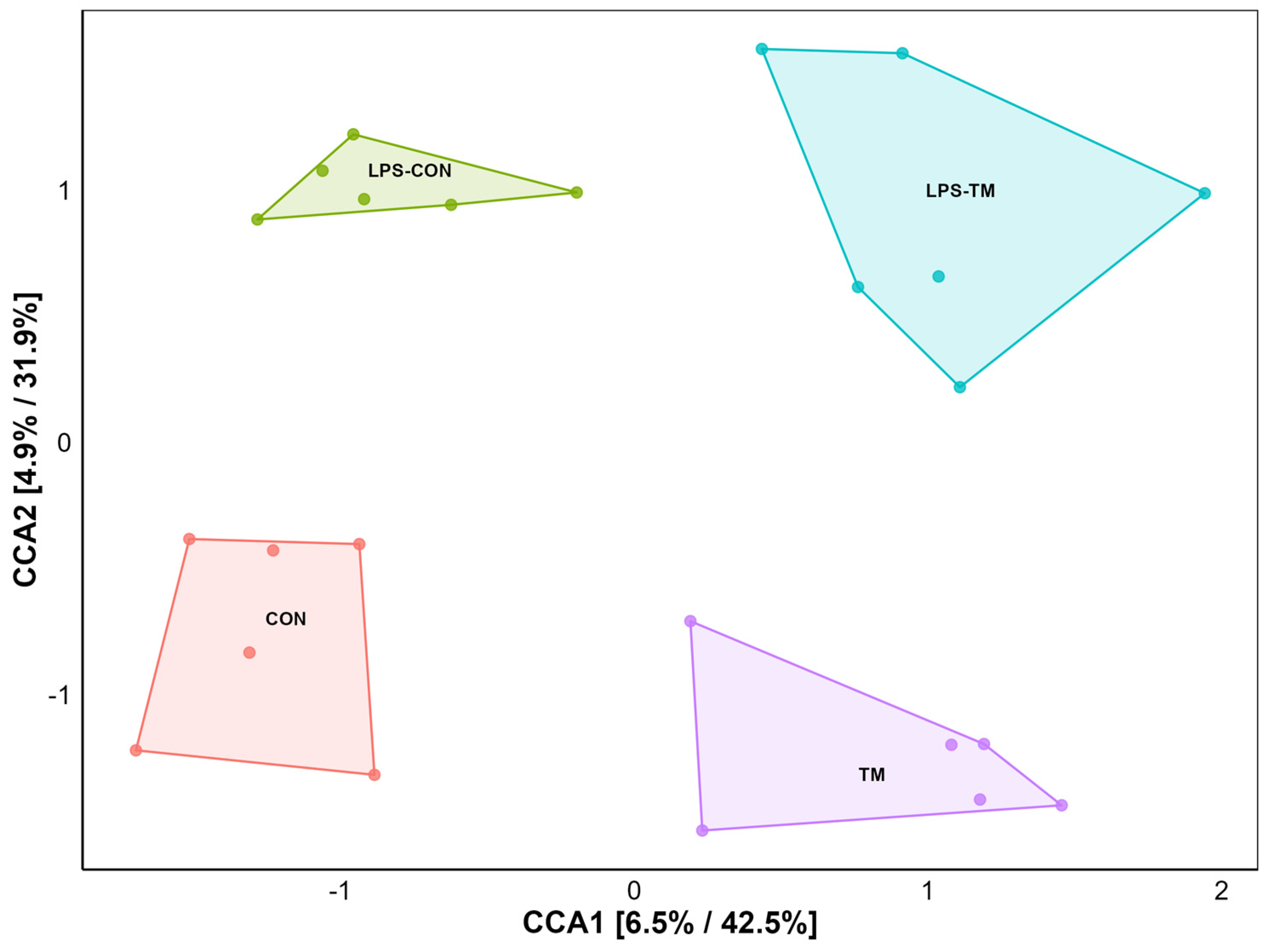
3.3. Effects of TM and LPS on Beta Diversity
CCA showed a significant separation among the cecal microbiota from the different experimental groups (Figure 4). Specifically, each group formed distinct clusters, indicating unique bacterial community compositions.
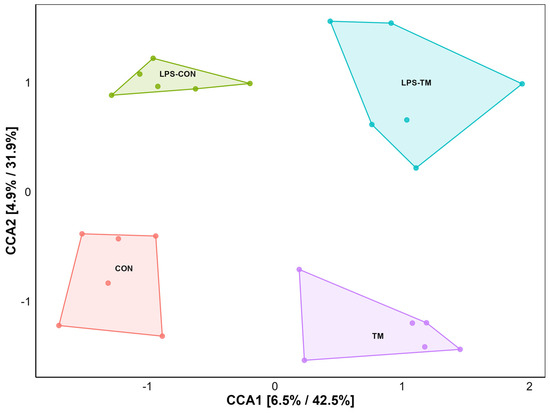
Figure 4.
Ordination plot for beta diversity generated by canonical correspondence analysis (CCA). The CCA plot illustrates the similarity between microbial communities based on beta diversity. Each point represents a sample.
Pairwise PERMANOVA analysis (Table 3) revealed significant differences between the TM and CON groups (F = 2.0033, p = 0.041), indicating that TM influenced microbial diversity. Similarly, a significant difference was observed between the TM and LPS-CON groups (F = 3.4913, p = 0.012).
Table 3.
Summary of PERMANOVA results for microbial community composition.
In contrast, the main effect of LPS was not significant (CON vs. LPS-CON: F = 1.0491, p = 0.398, and TM vs. LPS-TM: F = 1.5514, p = 0.080). PERMDISP confirmed homogeneity of variance (p = 0.21).
3.4. Effects of TM and LPS on Cecal Microbiota: Phylum-Level Composition
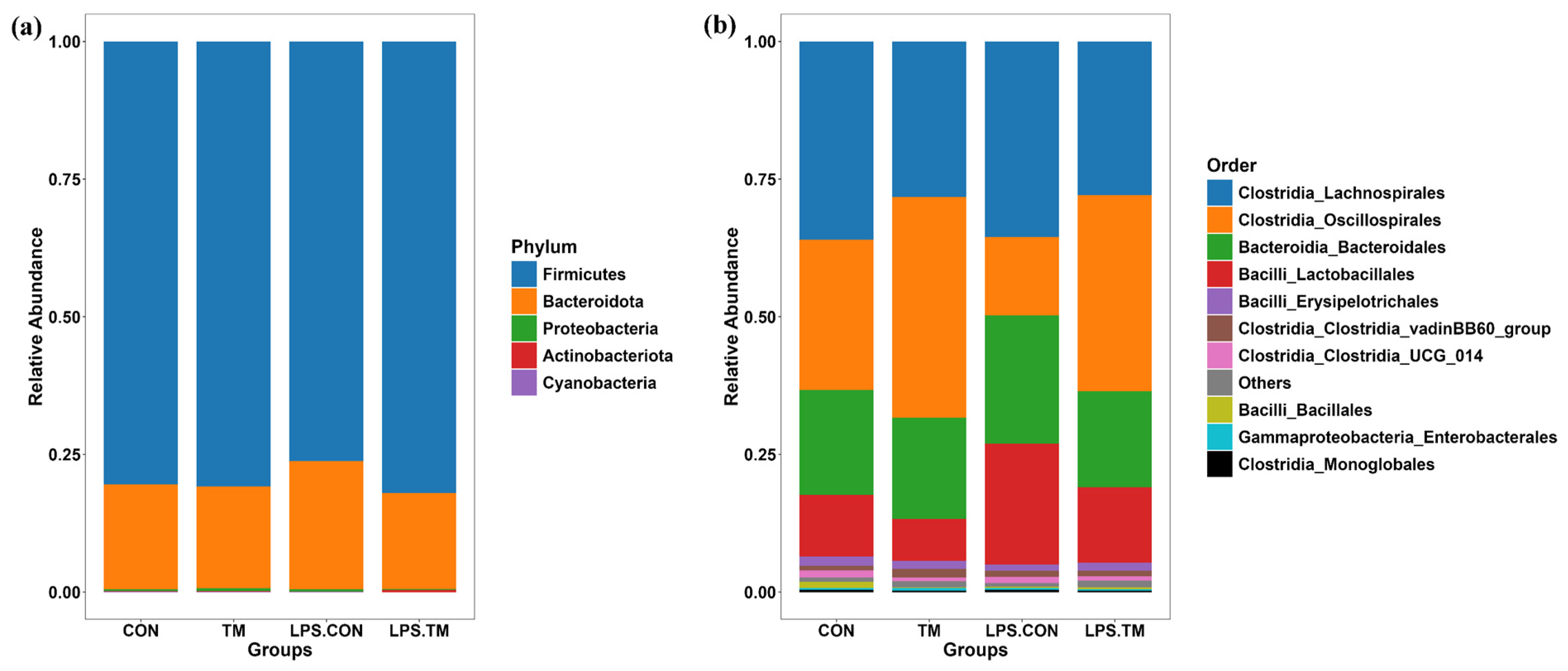
At the phylum level, Firmicutes (76.2–82%) and Bacteroidota (17.4–23.3%) were the dominant bacterial phyla across all groups. Minor contributions were observed from Proteobacteria (0.2–0.5%), Actinobacteriota (0.1–0.3%), and Cyanobacteria (0.0001–0.0009%) (Figure 5a).
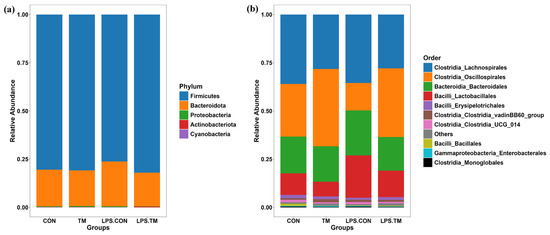
Figure 5.
Cecum microbiota composition at the phylum and order levels. (a) Composition at the phylum level. No significant differences were observed among groups. (b) Composition at the order level. Oscillospirales and Lactobacillales showed significantly different relative abundances (p < 0.05) between groups.
The LPS-CON group showed the lowest relative abundance (76.2%) of Firmicutes, while higher levels were observed in the CON (80%) and TM (80.1%) groups. Notably, the greatest relative abundance of Firmicutes (82%) occurred in the LPS-TM group. Conversely, Bacteroidota exhibited an inverse trend. The LPS-CON group had the highest relative abundance (23.3%), while the LPS-TM group showed the lowest (17.4%). Although these differences did not reach statistical significance (p > 0.05), this trend may highlight the potential of TM to modulate dominant bacterial phyla in response to immunological stress, such as LPS.
3.5. Effects of TM and LPS on Cecal Microbiota: Order-Level Composition
In all groups, the most prevalent bacterial orders were Lachnospirales (27.8–36%), Oscillospirales (14–40%), Bacteroidales (17.4–23.2%), and Lactobacillales (7.6–21.9%). Other bacterial orders, such as Erysipelotrichales, Clostridia vadinBB60—group, Clostridia UCG_014, Bacillales, Enterobacterales, and Monoglobales, were present at lower abundances (Figure 5b).
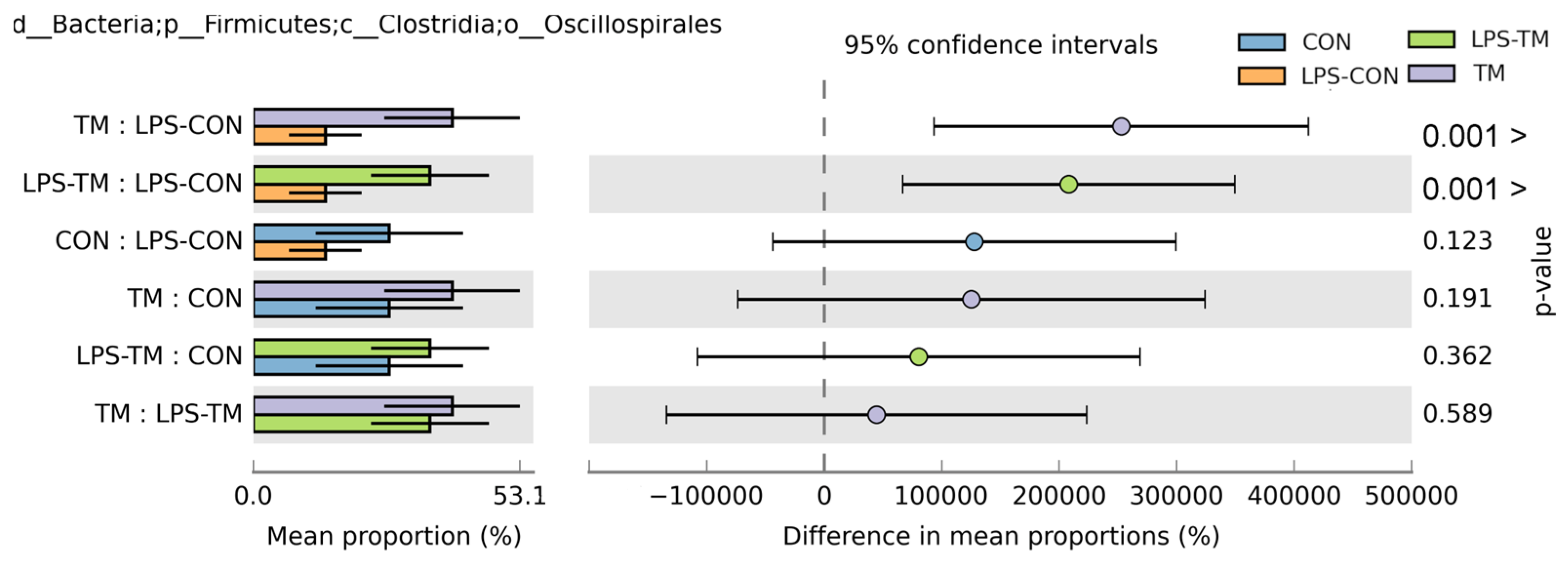
Oscillospirales showed statistically significant differences between the TM and LPS-TM groups compared to the LPS-CON group (Figure 6 and Figure 7b). The TM and LPS-TM groups exhibited higher relative abundances (40% and 35.6%, respectively) than the LPS-CON group (14.3%) (p < 0.001). These findings suggest that TM promotes the growth of Oscillospirales.
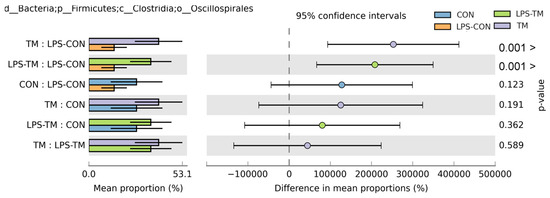
Figure 6.
Relative abundance of Oscillospirales across experimental groups. Results of the Kruskal–Wallis H-test indicate statistically significant differences (p < 0.05) in the relative abundance of Oscillospirales among the TM, LPS-TM, and LPS-CON groups. The TM and LPS-TM groups exhibited higher relative abundances (40% and 35.6%, respectively) than the LPS-CON group (14.3%). Error bars represent standard deviations.
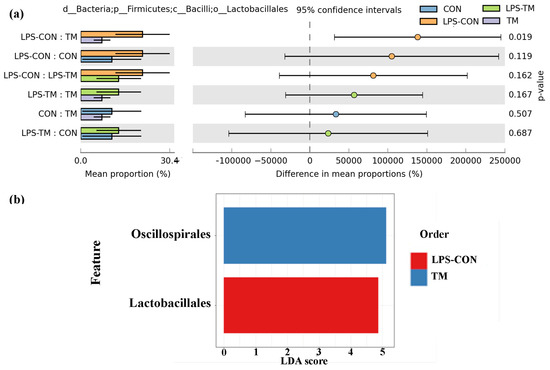
Figure 7.
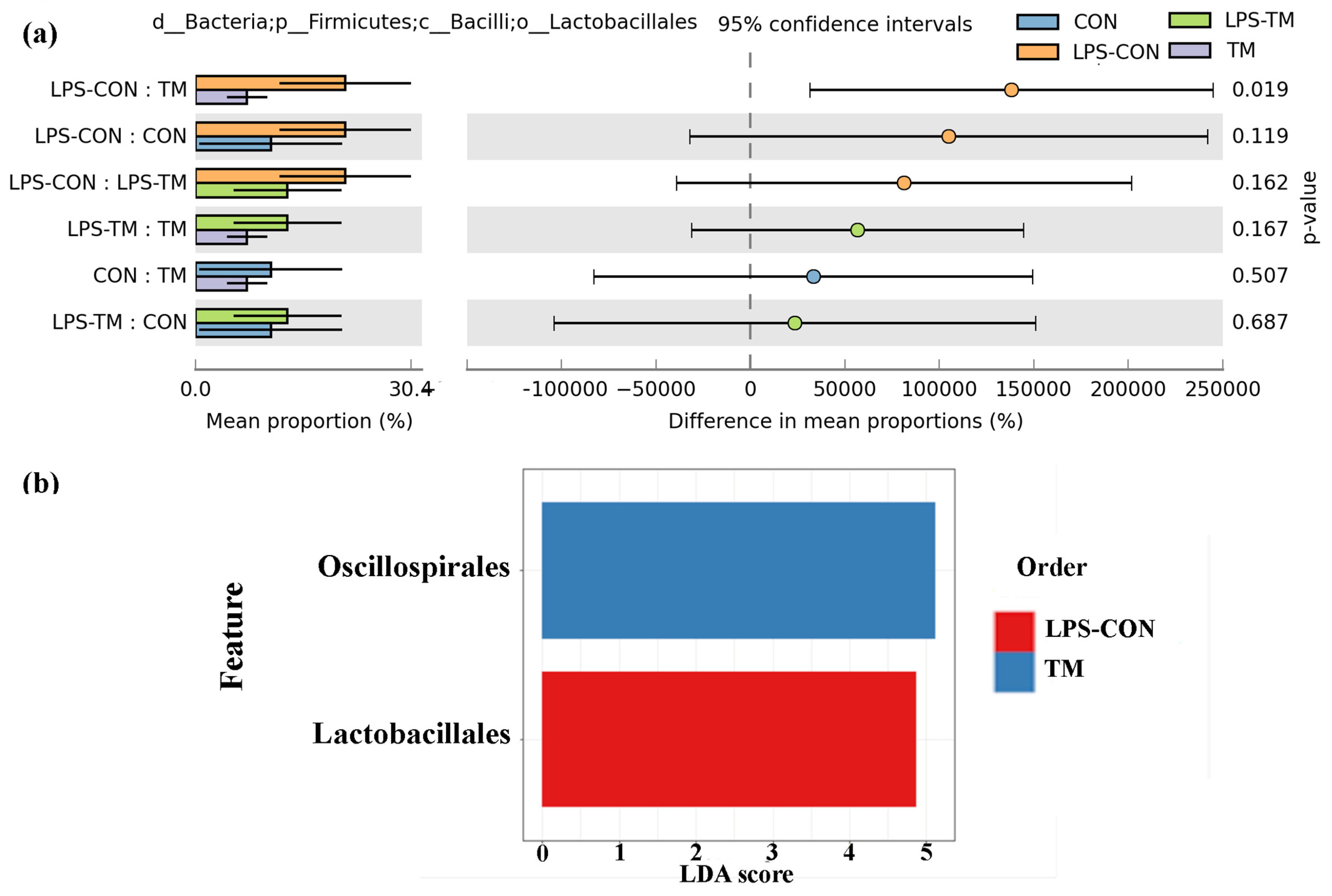
Representation of the relative abundance of Lactobacillales and LDA score across experimental groups. (a) A statistically significant difference (p < 0.019) in Lactobacillales abundance was observed between the TM and LPS-CON groups. (b) LDA further highlighted the enrichment of Lactobacillales in the LPS-CON group and the enrichment of Oscillospirales in the TM group.
The relative abundance of Lactobacillales in the LPS-CON group was 21.9%. A statistically significant difference (p < 0.05) in Lactobacillales abundance was observed between the TM and LPS-CON groups (Figure 7a). LDA further highlighted the enrichment of Lactobacillales in the LPS-CON group (Figure 7b).
3.6. TM and LPS Challenge Effects on Body Temperature (BT)
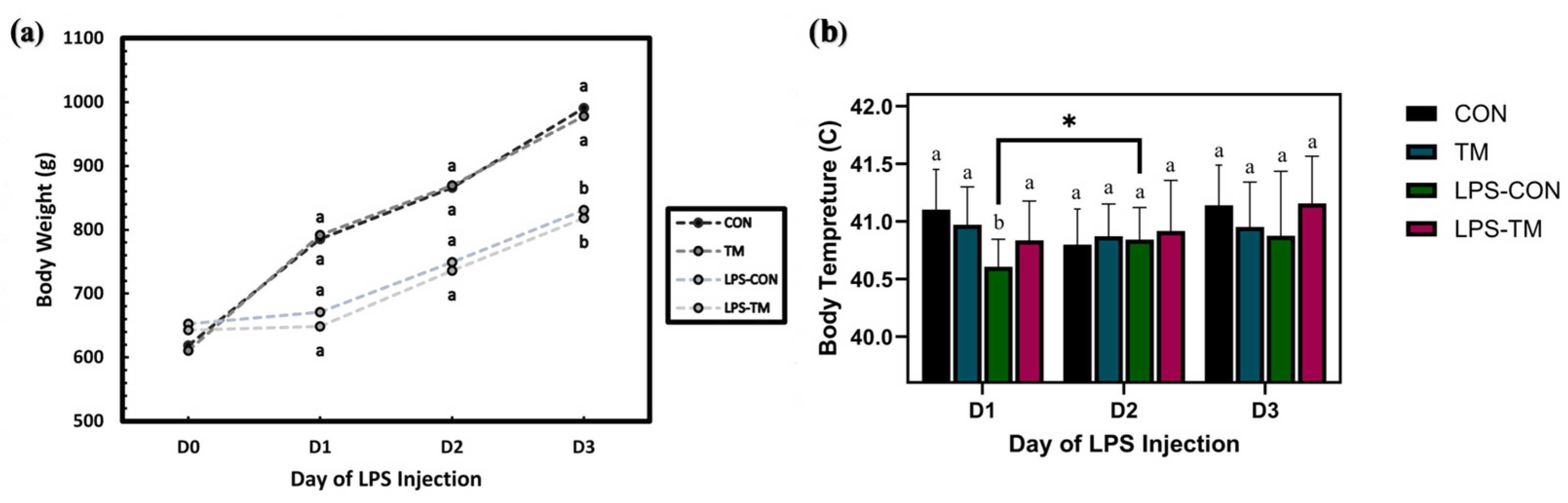
Figure 8a represents BT changes over time (Day 1 to Day 3), following LPS injections. In the LPS-CON group, BT initially decreased before increasing significantly (p = 0.013). This pattern reflects the typical response to LPS, which induces a hypothermic phase followed by fever as part of the acute-phase immune response. In contrast, the LPS-TM group bypassed the hypothermic phase, exhibiting a non-significant increase in body temperature over time (p > 0.05).
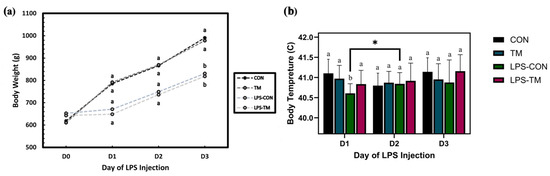
Figure 8.
Effect of LPS challenge on body weight (BW) and body temperature (BT) in broiler chickens subjected to temperature manipulation (TM) during embryogenesis. (a) LPS significantly reduced BW (p < 0.001), while TM did not significantly affect BW (p > 0.05). (b) In the LPS-CON group, BT initially decreased before increasing significantly (p = 0.013). In contrast, the LPS-TM group exhibited a non-significant increase in body temperature over time (p > 0.05). Groups labeled with different letters (a, b) differ significantly within the same day (p < 0.05); asterisks (*) indicate significant differences between days.
3.7. TM and LPS Challenge Effects on Body Weight (BW)
The line graph (Figure 8b) illustrates BW changes over time across the experimental groups in response to LPS injections. LPS significantly reduced BW (p < 0.001), highlighting its negative impact on growth performance. In contrast, TM did not significantly affect BW (p > 0.05), suggesting that TM may not mitigate the adverse effects of LPS on growth.
4. Discussion
This study provides new insights into how pre-hatch environmental manipulations, specifically TM, influence post-hatch microbial communities. It is among the first to investigate the impact of TM on gut microbiota composition, particularly under immunological challenges like exposure to LPS. While TM’s effects on broiler performance and heat stress resilience have been studied, its influence on gut microbial dynamics during LPS challenge has remained underexplored until now [9,10,11].
Population diversity is a crucial feature of a microbiome community that is closely linked to its environment. A change in diversity may indicate that the environment of the animal body has changed (from a healthy to a diseased state, for instance) or that certain conditions have disrupted the environment (e.g., a shift in the immune system or usage of antibiotics). Thus, one of the main goals of microbiome community research is to analyze the diversity of gut microbiota and discover host and/or environmental factors that can change the taxonomic composition [33].
Alpha diversity, which represents within-sample diversity, can be described using measures based on community richness (number of taxonomic groups), evenness (distribution of group abundances), or a combination of both [34]. The Chao1 index provides a nonparametric estimation of taxonomic richness based on abundance, while Shannon’s index (H) combines evenness and richness to estimate taxonomic diversity [35,36].
In this study, the Chao1 index was used to assess community richness, and Shannon’s index was used to represent community diversity. TM and LPS did not significantly affect overall microbial richness or diversity. These findings align with previous studies showing limited effects of LPS on alpha diversity [20,21,22].
Beta diversity measures differences in microbial community composition between groups or samples and is often more sensitive than alpha diversity metrics for detecting such distinctions [37]. In this study, a significant difference was detected between the TM and CON groups, underscoring TM’s role in shaping microbial community dynamics rather than changing microbial richness with the Bray–Curtis dissimilarity metric, which proved particularly effective in capturing these group-specific differences, highlighting the value of beta diversity in microbiome studies.
Research suggests that developmental stages and environmental factors during embryonic development significantly influence the diversity of gut microbiota. For example, Tong and Cui [38] reported that these factors shape microbiota composition in amphibians, while de Jong and Schokker [39] demonstrated that early-life environmental conditions affect gut microbial diversity in broilers. Among these factors, incubation temperature is a key regulator of epigenetic modifications, including DNA methylation and histone modifications, which influence microbial colonization and gut health [40,41]. TM specifically alters epigenetic regulation in embryos, leading to long-term physiological effects. According to David et al. [40], exposing chicken embryos to TM (39.5 °C for 12 h per day between the 7th and 16th day of embryogenesis) modifies post-translational histone markers, particularly H3K4me3 and H3K27me3, in the hypothalamus and muscle tissues. These changes contribute to environmental memory, impacting neurodevelopmental, metabolic, and immune regulatory pathways. Such epigenetic shifts likely extend to the gastrointestinal system, influencing gut microbiota composition and function.
Furthermore, the peri-hatching period is critical for microbiota establishment, as this window determines early microbial colonization, which can have long-lasting effects on birds’ health and development [41]. Early exposure to beneficial microbes allows advantageous bacterial populations to become dominant, and this is potentially modulated by TM-induced epigenetic mechanisms. Additionally, studies on in ovo microbiome stimulation with probiotics, prebiotics, and synbiotics suggest that these microbial interventions induce significant epigenetic changes, particularly in the liver and spleen. This further supports the link between early environmental interventions and gut microbiota regulation [42,43].
Animal health and performance at all levels of growth depend on maintaining an adequate balance of gut bacteria [44]. The cecum contains the most diverse microbial populations, which are essential for energy extraction and nutrient breakdown, ultimately affecting chickens’ intestinal health and growth efficiency [45,46]. In our study, Firmicutes and Bacteroidota phyla dominated the chickens’ cecal microbiota, which aligns with previous research [47]. However, the relative abundance of the dominant phyla can vary depending on factors such as diet and farming. For instance, free-range chickens tend to have higher levels of Bacteroidetes and Proteobacteria, while broiler chickens show a dominance of Firmicutes [48].
A high Firmicutes-to-Bacteroidota (F/B) ratio has previously been correlated with greater body weight in chickens [49]. Firmicutes are associated with butyrate and propionate production, which are short-chain fatty acids (SCFAs) linked to gut health and inflammation regulation [50]. In our study, the TM-LPS group exhibited a non-significantly higher F/B ratio, suggesting a potential trend toward improved growth performance.
At the order level, Oscillospirales were significantly enriched in the TM groups. These SCFA-producing bacteria are considered potential next-generation probiotics due to their role in synthesizing propionate, valerate, and butyrate [51,52]. Through the fermentation of dietary fibers and starch, Oscillospirales generate SCFAs that contribute to various physiological benefits, such as controlling apoptosis, lipid metabolism, and inflammation, influencing host DNA epigenetics, and maintaining the integrity of the epithelial barrier [53,54]. Specifically, as one of the principal butyrate producers, Oscillospirales play a vital role in gut health [55]. Butyrate serves as the primary energy source for enterocyte development and function [56]. Additionally, butyrate modulates the immune response by enhancing antioxidant enzyme activity, reducing pro-inflammatory cytokines, and inducing histone hyperacetylation through histone deacetylase (HDAC) inhibition—mechanisms that collectively regulate cell proliferation and likely contribute to the improved gut morphology previously observed in TM broilers [57,58,59].
Recent findings by Gałęcka et al. [60] further highlight the ecological significance of Oscillospirales, demonstrating that specific members (e.g., UCG-10) may respond to environmental contaminants like microplastics in porcine microbiomes, potentially contributing to both anti-inflammatory processes and contaminant degradation. Conversely, LPS-CON birds exhibited reduced Oscillospirales abundance, indicating that LPS exposure may selectively suppress these beneficial microbial taxa.
The increased abundance of Oscillospirales in the LPS-TM group may have contributed to the birds bypassing the hypothermic phase, indicating a stronger and more adaptive immune response. This could reflect enhanced cytokine production or other immunomodulatory effects induced by TM [9,14]. In contrast, the LPS-CON group displayed significantly lower BT post-LPS injection on Day 1 compared to the other groups, suggesting an initial hypothermic phase. This phase, characterized by reduced blood pressure, decreased circulating white blood cells, and increased apoptotic leukocytes, aligns with the early immune response to LPS [61].
Lactobacillales, a bacterial order well known for their probiotic properties, increased in the LPS-CON group in response to LPS-induced stress [62]. This microbial shift likely reflects a compensatory mechanism to mitigate gastrointestinal stress caused by LPS. Lactobacillales play a crucial role in immunoregulation, as their abundance has been positively correlated with SCFA production and CD4+ T-cell modulation, both of which contribute to immune homeostasis [63,64].
Additionally, Lactobacillales produce lactic acid, which lowers intestinal pH and creates an unfavorable environment for pathogenic bacteria, disrupting their outer membrane and limiting their colonization [65,66]. This shift in microbial composition may represent an adaptive host–microbiota interaction, where beneficial bacteria expand to protect the host from excessive inflammatory damage. Supporting this hypothesis, Alaqil [67] reported that Lactobacillus acidophilus (a key member of the Lactobacillales order) significantly alleviated stress markers and HSP70 expression in the spleen while increasing intestinal antioxidant capacity in LPS-challenged laying hens.
Furthermore, in the LPS-TM group, the abundance of Lactobacillales also did not significantly increase compared to the TM group, supporting the hypothesis that LPS may promote the growth of beneficial bacteria, such as Lactobacillales, to preserve gut health. According to Dai and Hackmann, LPS stimulation boosted the number of lactic acid-producing bacteria [68]. They also suggested that LPS would be detrimental to other bacteria that do not produce lactate, enhancing competition among lactate-producing bacteria and promoting their growth.
The observed effects of TM on gut microbiota composition may involve the microbiota–gut–brain (MGB) axis, a bidirectional communication system linking microbial dynamics with host stress responses [69]. TM has been shown to induce hypothalamic epigenetic adaptations and alter the levels of key hormones, such as plasma corticosterone, triiodothyronine (T3), and testosterone, especially during stressful situations [40,70]. These hormonal changes, mediated through mechanisms like the hypothalamic–pituitary–adrenal (HPA) axis, likely influence gut microbial composition and resilience [71,72]. Specifically, in chicken embryos, thyroid hormones and glucocorticoids work synergistically to promote gut maturation at the end of incubation. Thyroid hormones promote cellular differentiation and trigger the creation of digestive enzymes, whereas glucocorticoids aid in the development of intestinal glucose transport [73]. Interestingly, broiler chicks fed probiotic-supplemented diets have significantly increased blood T3 levels, highlighting the interrelated pathways between endocrine control, gut health, and microbial composition [74].
Compared to non-challenged groups, the LPS challenge reduced BW, consistent with previous studies demonstrating that LPS primarily affects appetite regulation and immune response, leading to weight loss. The mammalian target of rapamycin (mTOR) signaling pathway is activated by LPS, which causes anorexia by suppressing the expression of agouti-related protein (AgRP) in the hypothalamus, reducing food intake [13]. Furthermore, LPS exposure triggers the Toll-like receptor 4 (TLR4) signaling pathway, which might result in acute-phase responses and, consequently, due to immunological activation, lower body weight and feed intake may occur [75]. Remarkably, both the LPS-TM and LPS-CON groups exhibited similar growth suppression, suggesting that the LPS challenge diminished the beneficial effect of TM. However, microbiome analysis provides insight into how TM alters the intestinal ecology, indicating that TM might promote a more robust gut microbiota capable of responding to immunological stresses such as LPS.
TM shows promising effects on muscle development and growth-related hormones, but its success depends on incubation temperature, duration, and timing [76,77,78,79,80]. Studies suggest that specific TM protocols, such as incubating eggs at 38.2–38.4 °C for 2 h daily from day 18 until hatching or at 39.5 °C for 4 h daily from days 12 to 18, can increase BW [70,81]. However, other protocols, like 39.5 °C for 3 h daily during early, late, or combined embryonic periods, showed no significant effect on post-hatch BW [82]. Similarly, exposure to 39 °C and 65% RH for 12–18 h daily from days 12 to 18 significantly increased BW, while shorter durations did not [76]. These findings underscore the importance of optimizing TM conditions for the desired outcomes.
In our study, the significant decrease in BW post-LPS in the Indian River breed suggests that TM protocols may need further optimization for this specific breed. TM responses can vary across chicken breeds or types. For instance, TM in layer-type chickens has shown less favorable outcomes compared to broilers [83]. Limited literature exists on the effects of TM on the Indian River breed, highlighting the need for additional studies to optimize TM protocol for this breed.
This study underscores the potential of thermal manipulation (TM) as a non-invasive and cost-effective approach to enhancing gut microbiota resilience against immunological stress. TM-induced microbial shifts, particularly the enrichment of SCFA-producing taxa such as Oscillospirales, present promising avenues for improving poultry health and reducing antibiotic dependency. While TM enhances some microbial composition, its direct impact on growth performance during inflammatory stress remains unclear. Future research should prioritize metagenomic and metabolomic analyses to elucidate the functional pathways involved in TM-modulated gut microbiota. Such studies could provide a clearer understanding of TM’s potential in mitigating LPS-induced growth suppression. Integrating TM with nutritional strategies, including prebiotics, probiotics, phytogenic feed additives, and organic acids, offers a comprehensive solution to reduce antibiotic utilization and mitigate the risk of antimicrobial resistance (AMR).
5. Conclusions
The findings of this investigation demonstrate that thermal manipulation during embryogenesis dynamically and persistently modifies the gut microbiota. Moreover, by increasing beneficial bacteria and avoiding the hypothermic phase, TM significantly enhanced broiler response to the LPS challenge. These findings support TM as a viable antibiotic-free strategy for commercial poultry production. Future work should optimize protocols for different strains and integrate probiotic approaches. Standardized hatchery implementation could improve flock health while reducing antibiotic use, addressing critical industry needs.
6. Study Limitations
This study has several limitations that should be considered. First, the exclusive use of male chicks due to budget constraints may affect the generalizability of our findings, as established sex differences in broiler microbiome composition and function are well-documented. Second, while we focused on post-LPS microbiome changes, the characterization of microbial dynamics during the early post-hatch period (days 1–18) before the LPS challenge would have provided valuable insights into how TM shapes initial microbial colonization patterns. Additionally, although our findings suggest a potential link between TM, immune modulation, and metabolic efficiency, we did not directly measure muscle mass, carcass yield, or SCFA production. Future studies should incorporate muscle fiber histology, carcass composition analysis, metabolomic profiling, multi-timepoint microbiome analysis, and functional metagenomics to establish a more direct relationship between TM, microbial modulation, and overall broiler performance.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani15081149/s1: Detailed bioinformatics analysis (Word S1) and LDA scores with genus-level composition data (Word S2). Refs. [84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102] are cited in Supplementary Materials.
Author Contributions
Conceptualization, S.H. and M.B.A.-Z.; methodology, S.H., D.A., R.D., L.A. and M.A.M.; software, S.H., D.A., R.D. and M.A.M.; validation, M.B.A.-Z., L.A., M.A., D.E.G. and R.A.D.; formal analysis, S.H., D.A., M.M., M.A.M., M.B.A.-Z. and R.D.; investigation, D.A. and M.M.; resources, M.B.A.-Z. and M.A.; writing—original draft preparation, S.H.; writing—review and editing, M.B.A.-Z., M.A., L.A., R.A.D. and D.E.G.; visualization, S.H., D.A., R.D., M.A.M. and M.M.; supervision, M.B.A.-Z., L.A. and M.A.; funding acquisition, M.B.A.-Z. and M.A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Deanship of Research at Jordan University of Science and Technology, grant numbers 188/2019, 520/2023, and 400/2024.
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (Animal Care and Use Committee) of Jordan University of Science and Technology (JUST-ACUC) under protocol code 347/12/4/16.
Informed Consent Statement
Not applicable.
Data Availability Statement
All raw sequence data (FASTQ files) and QIIME 2 artifacts, including quality control files, feature tables, taxonomy tables, and rooted phylogenetic trees, associated with this study are publicly available and can be accessed through the following link: https://justedujo-my.sharepoint.com/:f:/g/personal/mbvlab_just_edu_jo/EhFXpakHEJhPgF-CzA2CqSEB0pvha6zKKggahhonMlQE0g?e=QsyF7j (accessed on 9 April 2025). Additionally, the raw sequence data are also available from the NCBI SRA database under accession number PRJNA1249260: https://www.ncbi.nlm.nih.gov/bioproject/1249260 (accessed on 26 March 2025).
Acknowledgments
The authors sincerely thank the Deanship of Research at Jordan University of Science and Technology for financially supporting this research project. They also express their sincere gratitude to Amany Al-Rashadieh and Eng. Ibrahim Alsukhni for their invaluable insights and exceptional technical support.
Conflicts of Interest
The authors have declared no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| TM | Thermal manipulation |
| CON | Control |
| LPS | Lipopolysaccharide |
| LPS-CON | Control with lipopolysaccharide challenge |
| LPS-TM | Thermal manipulation with lipopolysaccharide challenge |
| RH | Relative humidity |
| ED | Embryonic day |
| PH | Post-hatch |
| ASV | Amplicon sequencing variant |
| CCA | Canonical correspondence analysis |
| PERMANOVA | Permutational Multivariate Analysis of Variance |
| PERMDISP | Permutational analysis of multivariate dispersions |
References
- FAO. World Food and Agriculture—Statistical Yearbook 2023; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2023. [Google Scholar] [CrossRef]
- Zou, A.; Nadeau, K.; Wang, P.W.; Lee, J.Y.; Guttman, D.S.; Sharif, S.; Korver, D.R.; Brumell, J.H.; Parkinson, J. Accumulation of genetic variants associated with immunity in the selective breeding of broilers. BMC Genet. 2020, 21, 5. [Google Scholar] [CrossRef]
- Shterzer, N.; Sbehat, Y.; Poudel, B.; Rothschild, N.; Oloko, O.E.; Headrick, J.; Petersen, E.; Druyan, S.; Mills, E. Differences in gut bacterial community composition between modern and slower-growing broiler breeder lines: Implications of growth selection on microbiome composition. Front. Physiol. 2023, 14, 1151151. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, J.P.; Wilkie, D.C.; Van Kessel, A.G.; Drew, M.D. Potential strategies for controlling necrotic enteritis in broiler chickens in post-antibiotic era. Anim. Feed. Sci. Technol. 2006, 129, 60–88. [Google Scholar] [CrossRef]
- Nisha, A. Antibiotic residues—A global health hazard. Vet. World 2008, 1, 375. [Google Scholar] [CrossRef]
- O’Neill, J. Antimicrobial Resistance: Tackling a crisis for the health and wealth of nations. Rev. Antimicrob. Resist. 2014. [Google Scholar]
- Kisliouk, T.; Ziv, M.; Meiri, N. Epigenetic control of translation regulation: Alterations in histone H3 lysine 9 post-translation modifications are correlated with the expression of the translation initiation factor 2B (Eif2b5) during thermal control establishment. Dev. Neurobiol. 2010, 70, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Yahav, S.; Collin, A.; Shinder, D.; Picard, M. Thermal manipulations during broiler chick embryogenesis: Effects of timing and temperature. Poult. Sci. 2004, 83, 1959–1963. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Jaradat, Z.W.; Ababneh, M.M.; Okour, M.Z.; Saleh, K.M.M.; Alkofahi, A.; Alboom, M.H. Effects of embryonic thermal manipulation on the immune response to post-hatch Escherichia coli challenge in broiler chicken. Vet. World. 2023, 16, 918–928. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; El-Bahr, S.M. Thermal manipulation of the broilers embryos: Expression of muscle markers genes and weights of body and internal organs during embryonic and post-hatch days. BMC. Vet. Res. 2019, 15, 166. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Alliftawi, A.R.S.; Saleh, K.M.M.; Jaradat, Z.W. Expression of digestive enzyme and intestinal transporter genes during chronic heat stress in the thermally manipulated broiler chicken. Poult. Sci. 2019, 98, 4113–4122. [Google Scholar] [CrossRef]
- Loyau, T.; Bedrani, L.; Berri, C.; Metayer-Coustard, S.; Praud, C.; Coustham, V.; Mignon-Grasteau, S.; Duclos, M.J.; Tesseraud, S.; Rideau, N.; et al. Cyclic variations in incubation conditions induce adaptive responses to later heat exposure in chickens: A review. Animal 2015, 9, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Li, D.; Jiao, H.C.; Zhao, J.P.; Lin, H. Lipopolysaccharide inhibits hypothalamic Agouti-related protein gene expression via activating mechanistic target of rapamycin signaling in chicks. Gen. Comp. Endocrinol. 2021, 313, 113876. [Google Scholar] [CrossRef] [PubMed]
- Al-Zghoul, M.d.B.; Gerrard, D.; Dalloul, R. Dynamics of cytokines and heat shock proteins mRNA expression in thermally manipulated chicken challenged with intraperitoneally injection of Lipopolysaccharide (LPS). FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Wick, M.; Lilburn, M.S. Effect of embryonic thermal manipulation on heat shock protein 70 (HSP70) expression and subsequent immune response to post-hatch lipopolysaccharide challenge in Pekin ducklings. Poult. Sci. 2019, 98, 722–733. [Google Scholar] [CrossRef]
- Clavijo, V.; Florez, M.J.V. The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: A review. Poult. Sci. 2018, 97, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Akinyemi, F.T.; Ding, J.; Zhou, H.; Xu, K.; He, C.; Han, C.; Zheng, Y.; Luo, H.; Yang, K.; Gu, C.; et al. Dynamic distribution of gut microbiota during embryonic development in chicken. Poult. Sci. 2020, 99, 5079–5090. [Google Scholar] [CrossRef]
- Uni, Z.; Gal-Garber, O.; Geyra, A.; Sklan, D.; Yahav, S. Changes in Growth and Function of Chick Small Intestine Epithelium Due to Early Thermal Conditioning. Poult. Sci. 2001, 80, 438–445. [Google Scholar] [CrossRef]
- Amaz, S.A.; Chaudhary, A.; Mahato, P.L.; Jha, R.; Mishra, B. Pre-hatch thermal manipulation of embryos and post-hatch baicalein supplementation mitigated heat stress in broiler chickens. J. Anim. Sci. Biotechnol. 2024, 15, 8. [Google Scholar] [CrossRef]
- Wu, D.; Liang, S.; Du, X.; Xiao, J.; Feng, H.; Ren, Z.; Yang, X.; Yang, X. Effects of fecal microbiota transplantation and fecal virome transplantation on LPS-induced intestinal injury in broilers. Poult. Sci. 2024, 103, 103316. [Google Scholar] [CrossRef]
- Chen, J.Y.; Yu, Y.H. Bacillus subtilis-fermented products ameliorate the growth performance and alter cecal microbiota community in broilers under lipopolysaccharide challenge. Poult. Sci. 2021, 100, 875–886. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Lucke, A.; Doupovec, B.; Zebeli, Q.; Bohm, J. A multicomponent mycotoxin deactivator modifies the response of the jejunal mucosal and cecal bacterial community to deoxynivalenol contaminated feed and oral lipopolysaccharide challenge in chickens1. J. Anim. Sci. 2020, 98, skz377. [Google Scholar] [CrossRef] [PubMed]
- Al-Zghoul, M.B.; Mohammad Saleh, K.M. Effects of thermal manipulation of eggs on the response of jejunal mucosae to posthatch chronic heat stress in broiler chickens. Poult. Sci. 2020, 99, 2727–2735. [Google Scholar] [CrossRef] [PubMed]
- Al-Zghoul, M.B.; Sukker, H.; Ababneh, M.M. Effect of thermal manipulation of broilers embryos on the response to heat-induced oxidative stress. Poult. Sci. 2019, 98, 991–1001. [Google Scholar] [CrossRef]
- Shakouri, M.D.; Malekzadeh, M. Responses of broiler chickens to the nutrient recommendations of NRC (1994) and the Ross broiler management manual. Rev. Colom. Cienc. Pecua. 2016, 29, 91–98. [Google Scholar]
- Aviagen. Indian River Broiler Management Handbook. Available online: https://www.scribd.com/document/638577219/Untitled (accessed on 25 March 2024).
- Wu, Y.; Li, Q.; Liu, J.; Liu, Y.; Xu, Y.; Zhang, R.; Yu, Y.; Wang, Y.; Yang, C. Integrating Serum Metabolome and Gut Microbiome to Evaluate the Benefits of Lauric Acid on Lipopolysaccharide- Challenged Broilers. Front. Immunol. 2021, 12, 759323. [Google Scholar] [CrossRef]
- Zheng, Y.W.; Zhang, J.Y.; Zhou, H.B.; Guo, Y.P.; Ma, Q.G.; Ji, C.; Zhao, L.H. Effects of dietary pyrroloquinoline quinone disodium supplementation on inflammatory responses, oxidative stress, and intestinal morphology in broiler chickens challenged with lipopolysaccharide. Poult. Sci. 2020, 99, 5389–5398. [Google Scholar] [CrossRef]
- Pichler, M.; Coskun, O.K.; Ortega-Arbulu, A.S.; Conci, N.; Worheide, G.; Vargas, S.; Orsi, W.D. A 16S rRNA gene sequencing and analysis protocol for the Illumina MiniSeq platform. Microbiologyopen 2018, 7, e00611. [Google Scholar] [CrossRef]
- QIAGEN. DNeasy® PowerSoil® Pro Kit Handbook; QIAGEN: Hilden, Germany, 2023; Available online: https://www.qiagen.com/jp/resources/download.aspx?id=9bb59b74-e493-4aeb-b6c1-f660852e8d97&lang=en (accessed on 25 March 2024).
- Weinroth, M.D.; Oakley, B.; Ramirez, G.A.; Reyes, A.; Harris, C.E.; Buhr, R.J. 16S rRNA gene-based assessment of common broiler chicken sampling methods: Evaluating intra-flock sample size, cecal pair similarity, and cloacal swab similarity to other alimentary tract locations. Front. Physiol. 2022, 13, 996654. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, J. Alpha Diversity. In Bioinformatic and Statistical Analysis of Microbiome Data; Xia, Y., Sun, J., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 289–333. [Google Scholar]
- Willis, A.D. Rarefaction, Alpha Diversity, and Statistics. Front. Microbiol. 2019, 10, 2407. [Google Scholar] [CrossRef]
- Chao, A. Nonparametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Kers, J.G.; Saccenti, E. The power of microbiome studies: Some considerations on which alpha and beta metrics to use and how to report results. Front. Microbiol. 2022, 12, 796025. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Cui, L.Y.; Hu, Z.F.; Du, X.P.; Abid, H.M.; Wang, H.B. Environmental and host factors shaping the gut microbiota diversity of brown frog Rana dybowskii. Sci. Total Environ. 2020, 741, 140142. [Google Scholar] [CrossRef] [PubMed]
- de Jong, I.C.; Schokker, D.; Gunnink, H.; van Wijhe, M.; Rebel, J.M.J. Early life environment affects behavior, welfare, gut microbiome composition, and diversity in broiler chickens. Front. Vet. Sci. 2022, 9, 977359. [Google Scholar] [CrossRef]
- David, S.A.; Vitorino Carvalho, A.; Gimonnet, C.; Brionne, A.; Hennequet-Antier, C.; Piegu, B.; Crochet, S.; Courousse, N.; Bordeau, T.; Bigot, Y.; et al. Thermal Manipulation During Embryogenesis Impacts H3K4me3 and H3K27me3 Histone Marks in Chicken Hypothalamus. Front. Genet. 2019, 10, 1207. [Google Scholar] [CrossRef]
- Dunislawska, A.; Pietrzak, E.; Wishna Kadawarage, R.; Beldowska, A.; Siwek, M. Pre-hatching and post-hatching environmental factors related to epigenetic mechanisms in poultry. J. Anim. Sci. 2021, 100, skab370. [Google Scholar] [CrossRef]
- Sikorska, M.; Siwek, M.; Slawinska, A.; Dunislawska, A. miRNA Profiling in the Chicken Liver under the Influence of Early Microbiota Stimulation with Probiotic, Prebiotic, and Synbiotic. Genes 2021, 12, 685. [Google Scholar] [CrossRef]
- Dunislawska, A.; Slawinska, A.; Stadnicka, K.; Bednarczyk, M.; Gulewicz, P.; Jozefiak, D.; Siwek, M. Synbiotics for broiler chickens—In Vitro design and evaluation of the influence on host and selected microbiota populations following in ovo delivery. PLoS ONE 2017, 12, e0168587. [Google Scholar] [CrossRef]
- Trevisi, P.; Luise, D.; Correa, F.; Bosi, P. Timely Control of Gastrointestinal Eubiosis: A Strategic Pillar of Pig Health. Microorganisms 2021, 9, 313. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef]
- Timmerman, H.M.; Koning, C.J.; Mulder, L.; Rombouts, F.M.; Beynen, A.C. Monostrain, multistrain and multispecies probiotics—A comparison of functionality and efficacy. Int. J. Food Microbiol. 2004, 96, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Luo, L.; Wang, X.; Wen, Q.; Zhou, L.; Wu, K. Characterization of the cecal microbiome composition of Wenchang chickens before and after fattening. PLoS ONE 2019, 14, e0225692. [Google Scholar] [CrossRef]
- Mancabelli, L.; Ferrario, C.; Milani, C.; Mangifesta, M.; Turroni, F.; Duranti, S.; Lugli, G.A.; Viappiani, A.; Ossiprandi, M.C.; van Sinderen, D.; et al. Insights into the biodiversity of the gut microbiota of broiler chickens. Environ. Microbiol. 2016, 18, 4727–4738. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Ge, Y.; Xu, Z.; Zhang, D.; Li, D. The digestive and reproductive tract microbiotas and their association with body weight in laying hens. Poult. Sci. 2021, 100, 101422. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Ali, Q.; Ma, S.; Farooq, U.; Liu, B.; Wang, Z.; Sun, H.; Cui, Y.; Li, D.; Shi, Y. Chronological dynamics of the gut microbiome in response to the pasture grazing system in geese. Microbiol. Spectr. 2024, 12, e0418823. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Zhang, D.; Jian, Y.P.; Zhang, Y.N.; Li, Y.; Gu, L.T.; Sun, H.H.; Liu, M.D.; Zhou, H.L.; Wang, Y.S.; Xu, Z.X. Short-chain fatty acids in diseases. Cell Commun. Signal. 2023, 21, 212. [Google Scholar] [CrossRef]
- Wlazło, Ł.; Kowalska, D.; Bielański, P.; Ossowski, M.; Czech, A.; Łukaszewicz, M.; Nowakowicz-Dębek, B. Study of the Possibility of Modulating the Composition of the Gastrointestinal Microbiome of Rabbits Fed Fermented Rapeseed Meal. Ann. Anim. Sci. 2025, 25, 271–279. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Umar, M.; Hassan, F.-u.; Alagawany, M.; Arif, M.; Taha, A.E.; Elnesr, S.S.; El-Tarabily, K.A.; Abd El-Hack, M.E. Applications of butyric acid in poultry production: The dynamics of gut health, performance, nutrient utilization, egg quality, and osteoporosis. Anim. Health Res. Rev. 2022, 23, 136–146. [Google Scholar] [CrossRef]
- Zhang, W.H.; Jiang, Y.; Zhu, Q.; Gao, F.; Dai, S.; Chen, J.; Zhou, G. Sodium butyrate maintains growth performance by regulating the immune response in broiler chickens. Br. Poult. Sci. 2011, 52, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Kruh, J. Effects of sodium butyrate, a new pharmacological agent, on cells in culture. Mol. Cell. Biochem. 1981, 42, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Khaleel, K.E.; Al-Zghoul, M.B.; Saleh, K.M.M. Molecular and morphometric changes in the small intestine during hot and cold exposure in thermally manipulated broiler chickens. Vet. World 2021, 14, 1511. [Google Scholar] [CrossRef] [PubMed]
- Gałęcka, I.; Rychlik, A.; Całka, J. Influence of selected dosages of plastic microparticles on the porcine fecal microbiome. Sci. Rep. 2025, 15, 1269. [Google Scholar] [CrossRef]
- De Boever, S.; Croubels, S.; Meyer, E.; Sys, S.; Beyaert, R.; Ducatelle, R.; De Backer, P. Characterization of an intravenous lipopolysaccharide inflammation model in broiler chickens. Avian Pathol. 2009, 38, 403–411. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, J.; Pan, L.; Zhang, Y. Roles and applications of probiotic Lactobacillus strains. Appl. Microbiol. Biotechnol. 2018, 102, 8135–8143. [Google Scholar] [CrossRef]
- Bartley, A.; Yang, T.; Arocha, R.; Malphurs, W.L.; Larkin, R.; Magee, K.L.; Vickroy, T.W.; Zubcevic, J. Increased Abundance of Lactobacillales in the Colon of Beta-Adrenergic Receptor Knock Out Mouse Is Associated With Increased Gut Bacterial Production of Short Chain Fatty Acids and Reduced IL17 Expression in Circulating CD4+ Immune Cells. Front. Physiol. 2018, 9, 1593. [Google Scholar] [CrossRef]
- Pérez-Santiago, J.; Gianella, S.; Massanella, M.; Spina, C.A.; Karris, M.Y.; Var, S.R.; Patel, D.; Jordan, P.S.; Young, J.A.; Little, S.J.; et al. Gut Lactobacillales are associated with higher CD4 and less microbial translocation during HIV infection. AIDS 2013, 27, 1921–1931. [Google Scholar] [CrossRef]
- Ludwig, W.; Schleifer, K.-H.; Whitman, W.B. Lactobacillales ord. nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar]
- Alakomi, H.-L.; Skytta, E.; Saarela, M.; Mattila-Sandholm, T.; Latva-Kala, K.; Helander, I. Lactic acid permeabilizes gram-negative bacteria by disrupting the outer membrane. Appl. Environ. Microbiol. 2000, 66, 2001–2005. [Google Scholar] [CrossRef]
- Alaqil, A. The Effect of Lactobacillus Acidophilus on Alleviating Stress Response and Production Impairment Induced by Escherichia Coli Lipopolysaccharide in Laying Hens. Adv. Anim. Vet. Sci. 2023, 11, 1183–1192. [Google Scholar] [CrossRef]
- Dai, X.; Hackmann, T.J.; Lobo, R.R.; Faciola, A.P. Lipopolysaccharide Stimulates the Growth of Bacteria That Contribute to Ruminal Acidosis. Appl. Environ. Microbiol. 2020, 86, e02193-19. [Google Scholar] [CrossRef]
- Dinan, T.G.; Cryan, J.F. Gut instincts: Microbiota as a key regulator of brain development, ageing and neurodegeneration. J. Physiol. 2017, 595, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Iraqi, E.; Hady, A.A.; Elsayed, N.; Khalil, H.; El-Saadany, A.; El-Sabrout, K. Effect of thermal manipulation on embryonic development, hatching process, and chick quality under heat-stress conditions. Poult. Sci. 2024, 103, 103257. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203. [Google Scholar] [PubMed]
- Tetel, M.J.; de Vries, G.J.; Melcangi, R.C.; Panzica, G.; O’Mahony, S.M. Steroids, stress and the gut microbiome-brain axis. J. Neuroendocrinol. 2018, 30, e12548. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L. Influence of hormones on glycogen and glucose metabolism in embryonic chick intestine. Am. J. Physiol. 1988, 254, G65–G73. [Google Scholar] [CrossRef]
- Chotinsky, D.; Mihaylov, R. Effect of probiotics and avotan on the level of thyroid hormones in the blood plasma of broiler chickens. Bulgarian J. Agric. Sci. 2013, 19, 817–821. [Google Scholar]
- Li, N.; Ansari, A.R.; Sun, Z.; Huang, H.; Cui, L.; Hu, Y.; Zhao, X.; Zhong, J.; Abdel-Kafy, E.M.; Liu, H. Toll like receptor 4 signaling pathway participated in Salmonella lipopolysaccharide-induced spleen injury in young chicks. Microb. Pathog. 2017, 112, 288–294. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Al-Natour, M.; Dalab, A.; Alturki, O.; Althnaian, T.; Al-Ramadan, S.; Hannon, K.; El-Bahr, S. Thermal manipulation mid-term broiler chicken embryogenesis: Effect on muscle growth factors and muscle marker genes. Braz. J. Poult. Sci. 2016, 18, 607–618. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Wang, K.; Liu, L.; Chen, K.; Shan, W.; Liu, L.; Kahiel, M.; Li, C. Embryo thermal manipulation enhances mitochondrial function in the skeletal muscle of heat-stressed broilers by regulating transient receptor potential V2 expression. Poult. Sci. 2024, 103, 104034. [Google Scholar] [CrossRef] [PubMed]
- Dalab, A.S.; Ali, A.M.; Althnaian, T.A.; Alkhodair, K.M.; Al-Ramadan, S.Y. Molecular and ultrastructural investigations of the effect of thermal manipulation during embryogenesis on pectoral and thigh muscles growth factors in broilers. J. Appl. Poult. Res. 2022, 31, 100188. [Google Scholar] [CrossRef]
- Piestun, Y.; Harel, M.; Barak, M.; Yahav, S.; Halevy, O. Thermal manipulations in late-term chick embryos have immediate and longer term effects on myoblast proliferation and skeletal muscle hypertrophy. J. Appl. Physiol. 2009, 106, 233–240. [Google Scholar] [CrossRef]
- Piestun, Y.; Yahav, S.; Halevy, O. Thermal manipulation during embryogenesis affects myoblast proliferation and skeletal muscle growth in meat-type chickens. Poult. Sci. 2015, 94, 2528–2536. [Google Scholar] [CrossRef] [PubMed]
- Tzschentke, B.; Halle, I. Influence of temperature stimulation during the last 4 days of incubation on secondary sex ratio and later performance in male and female broiler chicks. Br. Poult. Sci. 2009, 50, 634–640. [Google Scholar] [CrossRef]
- Collin, A.; Berri, C.; Tesseraud, S.; Rodon, F.E.; Skiba-Cassy, S.; Crochet, S.; Duclos, M.J.; Rideau, N.; Tona, K.; Buyse, J.; et al. Effects of thermal manipulation during early and late embryogenesis on thermotolerance and breast muscle characteristics in broiler chickens. Poult. Sci. 2007, 86, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Walstra, I.; Ten Napel, J.; Kemp, B.; van den Brand, H. Temperature manipulation during layer chick embryogenesis. Poult. Sci. 2010, 89, 1502–1508. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Waste not, want not: Why rarefying microbiome data is inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef]
- Schloss, P.D. Waste not, want not: Revisiting the analysis that called into question the practice of rarefaction. Msphere 2024, 9, e00355-23. [Google Scholar] [CrossRef]
- Team RS. RStudio: Integrated Development Environment for R; RStudio Inc.: Boston, MA, USA, 2021. [Google Scholar]
- R Core Team. RA Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018. [CrossRef]
- ter Braak, C.J.F. Canonical Correspondence Analysis: A New Eigenvector Technique for Multivariate Direct Gradient Analysis. Ecology 1986, 67, 1167–1179. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Ordination; Cambridge University Press: Cambridge, UK, 1995; Volume 5. [Google Scholar]
- Gauch, H.G., Jr.; Whittaker, R.H.; Singer, S.B. A comparative study of nonmetric ordinations. J. Ecol. 1981, 69, 135–152. [Google Scholar] [CrossRef]
- Gauch, H.G., Jr. Noise reduction by eigenvector ordinations. Ecology 1982, 63, 1643–1649. [Google Scholar] [CrossRef]
- Wilmes, P.; Bond, P.L. The application of two-dimensional polyacrylamide gel electrophoresis and downstream analyses to a mixed community of prokaryotic microorganisms. Environ. Microbiol. 2004, 6, 911–920. [Google Scholar] [CrossRef]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern Wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Anderson, M.J.; Walsh, D.C. PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: What null hypothesis are you testing? Ecol. Monogr. 2013, 83, 557–574. [Google Scholar] [CrossRef]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).