NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours


Abstract
:1. Introduction
2. Materials and Methods
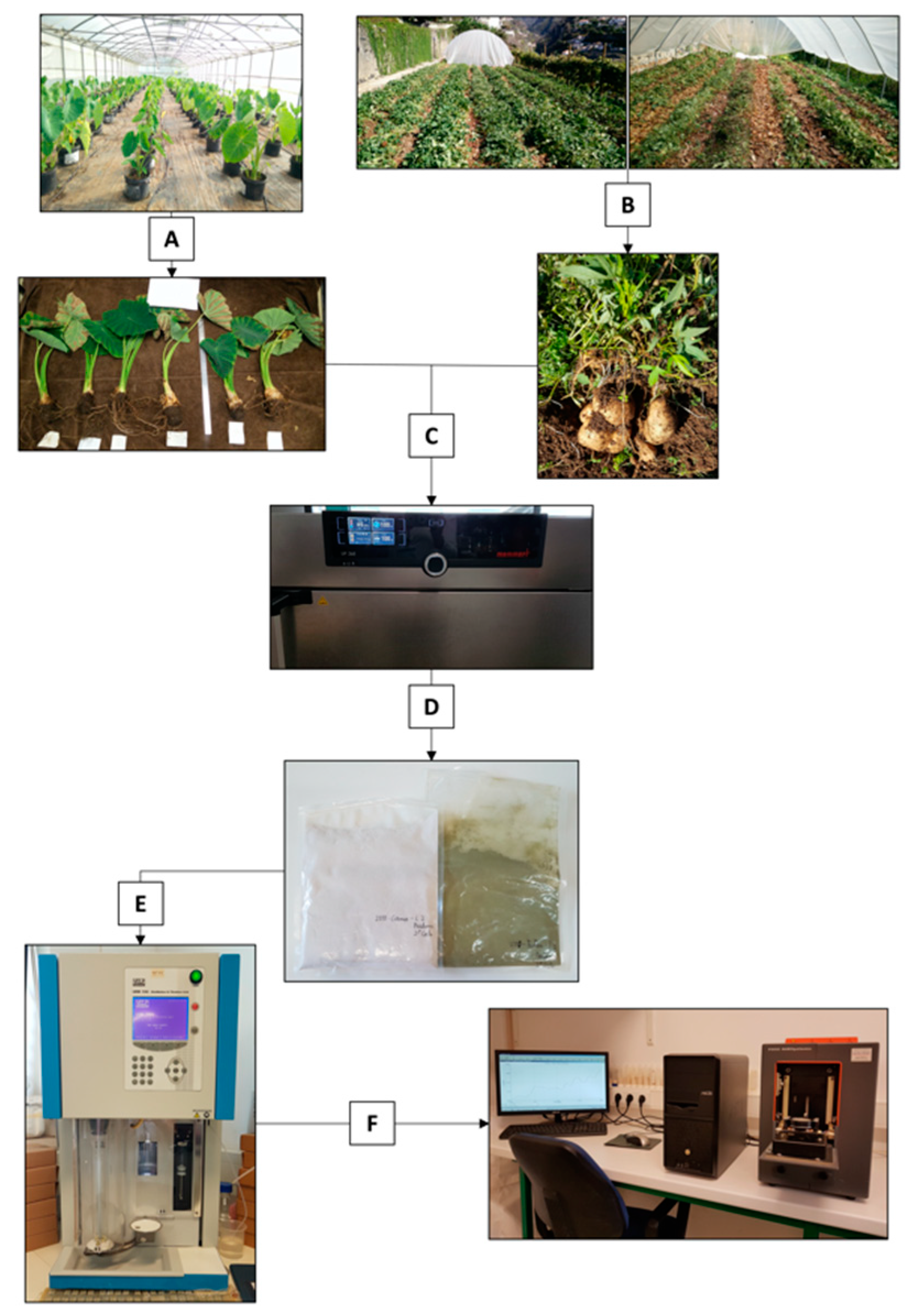
2.1. Sample Preparation and Chemical Analysis
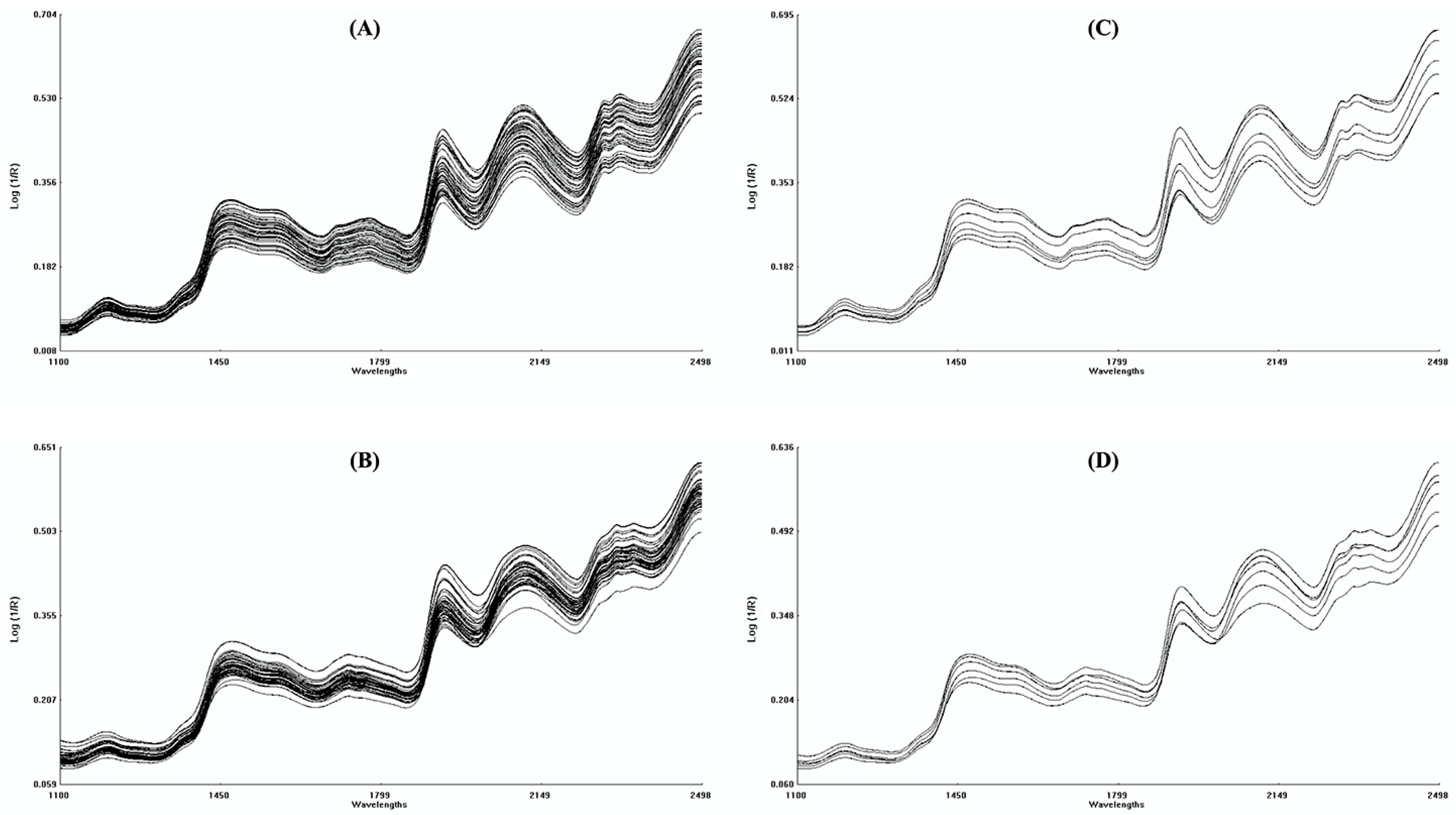
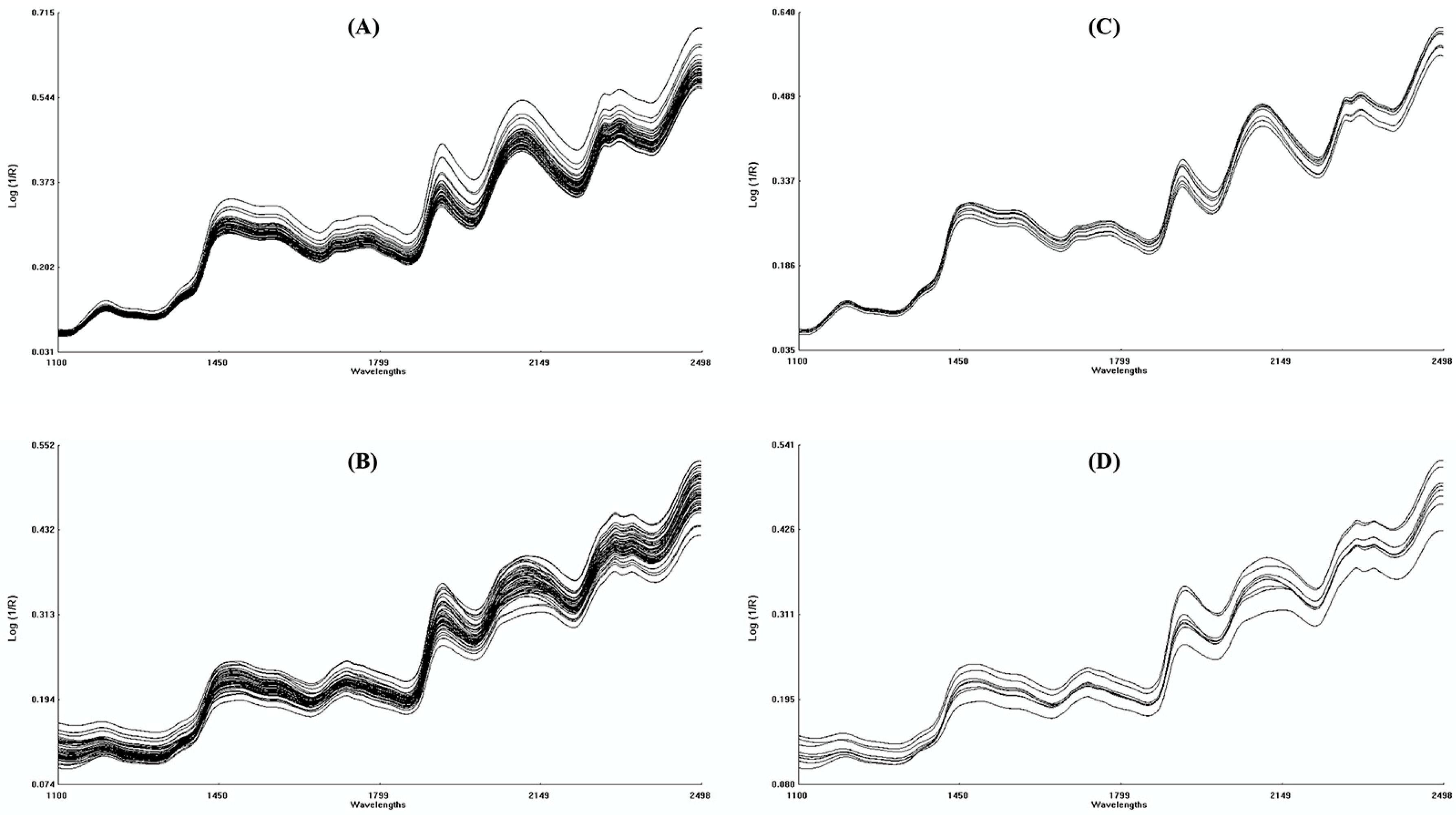
2.2. NIRS Measurements, Data Pretreatment and Analysis
2.3. Statistical Analysis
3. Results and Discussion
3.1. NIRS Calibration and Validation of PLS Models
3.1.1. Taro
3.1.2. Sweet Potato
3.2. Variability of Chemical Constituents
3.2.1. Taro
3.2.2. Sweet Potato
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAOSTAT Database. 2019. Available online: www.fao.org (accessed on 21 September 2019).
- Laxminarayana, K.; Mishra, S.; Soumya, S. Good Agricultural Practices in Tropical Root and Tuber Crops. In Tropical Roots and Tubers–Production, Processing and Technology; Sharma, H.K., Njintang, N.Y., Singhal, R.S., Kaushal, P., Eds.; John Wiley & Sons: Oxford, UK, 2016; pp. 192–204. [Google Scholar]
- Sharma, H.K.; Kaushal, P.; Singh, B. Taro Flour, Achu and Starch. In Tropical Roots and Tubers–Production, Processing and Technology; Sharma, H.K., Njintang, N.Y., Singhal, R.S., Kaushal, P., Eds.; John Wiley & Sons: Oxford, UK, 2016; pp. 326–327. [Google Scholar]
- Bradbury, J.H.; Holloway, W.D. Chemistry of tropical root crops: Significance for nutrition and agriculture in the Pacific. In Australian Centre for International Agricultural Research (ACIAR); (Monograph No. 6); ACIAR Monograph Series: Canberra, Australia, 1988; pp. 18, 160. [Google Scholar]
- Lebot, V. Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yams and Aroids; Atherton, J., Rees, A., Eds.; CAB International: Cambridge, UK, 2009; Volume xvi, pp. 134–135, 167–168, 325–329, 351. [Google Scholar]
- Woolfe, J.A. Sweet Potato: An Untapped Food Resource; Woolfe, J.A., Ed.; Cambridge University Press: Cambridge, UK, 1992; pp. 61–62. [Google Scholar]
- Oscarsson, K.V.; Savage, G.P. Composition and availability of soluble and insoluble oxalates in raw and cooked taro (Colocasia esculenta var. Schott) leaves. Food Chem. 2007, 101, 559–562. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Horner, H.T. Calcium oxalate crystals in plants. Bot. Rev. 1980, 46, 361–427. [Google Scholar] [CrossRef]
- Mulugeta, M.; Tebeka, T. Proximate and some minerals analysis of Colocasia esculenta (taro) tuber in Southern Ethiopia. Int. J. Pharm. Pharm. Res. Hum. J. 2017, 10, 1–12. [Google Scholar]
- Magwaza, L.S.; Naidoo, S.I.M.; Laurie, S.M.; Laing, M.D.; Shimelis, H. Development of NIRS models for rapid quantification of protein content in sweet potato [Ipomoea batatas (L.) Lam.]. LWT Food Sci. Technol. 2016, 72, 63–70. [Google Scholar] [CrossRef]
- Temesgen, M.; Retta, N.; Tesfaye, E. Effect of pre-curding on nutritional and anti-nutritional composition of taro (Colocasia esculenta L.) leaf. Int. J. Food Sci. Nutr. 2016, 1, 5–11. [Google Scholar]
- Kumoro, A.C.; Putri, R.D.A.; Budiyati, C.S.; Retnowati, D.S. Kinetics of calcium oxalate reduction in taro (Colocasia esculenta) corm chips during treatments using baking soda solution. Procedia Chem. 2014, 9, 102–112. [Google Scholar] [CrossRef] [Green Version]
- Alcantara, R.M.; Hurtada, W.A.; Dizon, E.I. The nutritional value and phytochemical components of taro [Colocasia esculenta (L.) Schott] powder and its selected processed foods. J. Nutr. Food Sci 2013, 3, 207. [Google Scholar] [CrossRef] [Green Version]
- Fai, F.Y.; Danbature, W.L.; Yahaya, A.; Usman, Y.M. Proximate and some minerals analysis of Colocasia esculenta (taro) leaves. J. Med. Biol. Sci. 2013, 3, 8–14. [Google Scholar]
- Amon, A.S.; Soro, R.Y.; Koffi, P.K.B.; Dué, E.A.; Kouamé, L.P. Biochemical characteristics of flours from ivorian taro (Colocasia esculenta, Cv Yatan) corm as affected by boiling time. Adv. J. Food Sci. Technol. 2011, 3, 424–435. [Google Scholar]
- Lebot, V.; Ndiaye, A.; Malapa, R. Phenotypic characterization of sweet potato [Ipomoea batatas (L.) Lam.] genotypes in relation to prediction of chemical quality constituents by NIRS equations. Plant Breed. 2011, 130, 457–463. [Google Scholar] [CrossRef]
- Lebot, V.; Malapa, R.; Bourrieau, M. Rapid estimation of taro (Colocasia esculenta) quality by near-infrared reflectance spectroscopy. J. Agric. Food Chem. 2011, 59, 9327–9334. [Google Scholar] [CrossRef] [PubMed]
- Lebot, V.; Champagne, A.; Malapa, R.; Shiley, D. NIR determination of major constituents in tropical root and tuber crop flours. J. Agric. Food Chem. 2009, 57, 10539–10547. [Google Scholar] [CrossRef] [PubMed]
- Motsa, N.M.; Modi, A.T.; Mabhaudhi, T. Sweet potato response to low-input agriculture and varying environments of KwaZulu-Natal, South Africa: Implications for food security strategies. Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 329–340. [Google Scholar] [CrossRef]
- Ekanayake, I.J.; Collins, W. Effect of irrigation on sweet potato root carbohydrates and nitrogenous compounds. Food Agric. Environ. 2004, 2, 243–248. [Google Scholar]
- Ishida, H.; Suzuno, H.; Sugiyama, N.; Innami, S.; Tadokoro, T.; Maekawa, A. Nutritive evaluation on chemical components of leaves, stalks and stems of sweet potatoes (Ipomoea batatas poir). Food Chem. 2000, 68, 359–367. [Google Scholar] [CrossRef]
- Ravindran, V.; Ravindran, G.; Sivakanesan, R.; Rajaguru, S.B. Biochemical and nutritional assessment of tubers from 16 cultivars of sweet potato (Ipomoea batatas L.). J. Agric. Food Chem. 1995, 43, 2646–2651. [Google Scholar] [CrossRef]
- Holloway, W.D.; Argall, M.E.; Jealous, W.T.; Lee, J.A.; Bradbury, J.H. Organic acids and calcium oxalate in tropical root crops. J. Agric. Food Chem. 1989, 37, 337–341. [Google Scholar] [CrossRef]
- Tattiyakul, J.; Pradipasena, P.; Asavasaksakul, S. Taro Colocasia esculenta (L.) Schott amylopectin structure and its effect on starch functional properties. Starch/Staerke 2007, 59, 342–347. [Google Scholar] [CrossRef]
- Tattiyakul, J.; Asavasaksakul, S.; Pradipasena, P. Chemical and physical properties of flour extracted from taro Colocasia esculenta (L.) Schott grown in different regions of Thailand. SenseAsia 2006, 32, 279–284. [Google Scholar]
- Ejoh, A.R.; Mbiapo, F.T.; Fokou, E. Nutrient composition of the leaves and flowers of Colocasia esculenta and the fruits of Solanum melongena. Plant Foods Hum. Nutr. 1996, 49, 107–112. [Google Scholar] [CrossRef]
- Ganança, J.F.T.; Freitas, J.G.R.; Nóbrega, H.G.M.; Rodrigues, V.; Antunes, G.; Gouveia, C.S.S.; Rodrigues, M.; Chaϊr, H.; de Carvalho, M.Â.A.P.; Lebot, V. Screening for drought tolerance in thirty three taro cultivars. Not. Bot. Horti Agrobot. Cluj Napoca 2018, 46, 65–74. [Google Scholar] [CrossRef]
- Ganança, J.F.T.; Freitas, J.G.F.; Nóbrega, H.G.M.; Rodrigues, V.; Antunes, G.; Rodrigues, M.; de Carvalho, M.Â.A.P.; Lebot, V. Screening of elite and local taro (Colocasia esculenta) cultivars for drought tolerance. Procedia Environ. Sci. 2015, 29, 41–42. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhu, L.; Zhang, C.; Ning, Y.; Zhang, Y. Effect of water stress on dry mass accumulation and allocation in sweet potato based on stable isotope analysis. Can. J. Plant Sci. 2015, 95, 263–271. [Google Scholar] [CrossRef]
- Williams, P. Grains and Seeds. In Near-Infrared Spectroscopy in Food Science and Technology; Ozaki, Y., McClure, W.F., Christy, A.A., Eds.; Wiley-Interscience: Hoboken, NJ, USA, 2007; pp. 165–217. [Google Scholar]
- Siesler, H.W.; Ozaki, Y.; Kawata, S.; Heise, H.M. Near-Infrared Spectroscopy–Principles, Instruments, Applications; WILEY-VCH Press: Weinheim, Germany, 2002. [Google Scholar]
- Pasquini, C. Near infrared spectroscopy: A mature analytical technique with new perspectives—A review. Anal. Chim. Acta 2018, 1026, 8–36. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Bosquet, L.; Crossa, J.; von Zitewitz, J.; Serret, M.D.; Araus, J.L. High-throughput phenotyping and genomic selection: The frontiers of crop Breeding converge. J. Integr. Plant Biol. 2012, 54, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Champagne, A.; Legendre, L.; Lebot, V. Chemotype profiling to guide breeders and explore traditional selection of root crops in Vanuatu, South Pacific. J. Agric. Food Chem. 2009, 57, 10363–10370. [Google Scholar] [CrossRef]
- Desjardins, A.Z. Natural product chemistry meets genetics: When is a genotype a chemotype? J. Agric. Food Chem. 2008, 56, 7587–7592. [Google Scholar] [CrossRef]
- Hatfield, J.L. Climate change: Challenges for future crop adjustments. In Climate Change and Plant Abiotic Stress Tolerance; Tuteja, N., Gill, S.S., Eds.; Wiley-Blackwell: Weinheim, Germany, 2014; p. 4. [Google Scholar]
- Wang, Y.; Frei, M. Stressed food—The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 2011, 141, 271–286. [Google Scholar] [CrossRef]
- Ferrio, J.P.; Bertran, E.; Nachit, M.; Royo, C.; Araus, J.L. Near infrared reflectance spectroscopy as a potential surrogate method for the analysis of Δ13C in mature kernels of durum wheat. Aust. J. Agric. Res. 2001, 52, 809–816. [Google Scholar] [CrossRef]
- Bruno-Soares, A.M.; Murray, I.; Paterson, R.M.; Abreu, J.M.F.F. Use of near infrared reflectance spectroscopy (NIRS) for the prediction of the chemical composition and nutritional attributes of green crop cereals. Anim. Feed Sci. Technol. 1998, 75, 15–25. [Google Scholar] [CrossRef]
- de Boever, J.L.; Cottyn, B.G.; Vanacker, J.M.; Boucqué, C.V. The use of NIRS to predict the chemical composition and the energy value of compound feeds for cattle. Anim. Feed Sci. Technol. 1995, 51, 243–253. [Google Scholar] [CrossRef]
- Kleinebecker, T.; Schmidt, S.R.; Fritz, C.; Smolders, A.J.P.; Holzel, N. Prediction of δ13C and δ15N in plant tissues with near-infrared reflectance spectroscopy. New Phytol. 2009, 184, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, C.S.S.; Ganança, J.F.T.; de Nóbrega, H.G.M.; de Freitas, J.G.R.; Lebot, V.; de Carvalho, M.Â.A.P. Phenotypic flexibility and drought avoidance in taro (Colocasia esculenta (L.) Schott). Emir. J. Food Agric. 2020, 32, 150–159. [Google Scholar] [CrossRef]
- Gouveia, C.S.S.; Ganança, J.F.T.; de Nóbrega, H.G.M.; de Freitas, J.G.R.; Lebot, V.; de Carvalho, M.Â.A.P. Drought avoidance and phenotypic flexibility of sweet potato (Ipomoea batatas (L.) Lam.) under water scarcity conditions. Not. Bot. Horti Agrobot. Cluj Napoca 2019, 47, 1037–1046. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis; AOAC International Publisher: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Hodge, J.E.; Hofreiter, B.T. Analysis and preparation of sugars. In Methods in Carbohydrate Chemistry, 6th ed.; Whistler, R.L., Miller, J.N.B., Eds.; Academic Press: New York, NY, USA, 1962; pp. 356–378. [Google Scholar]
- ISO. Agricultural Food Products. Determination of Crude Fibre Content-Modified Scharrer, Method 6541; International Organization of Standardization: Geneva, Switzerland, 1981. [Google Scholar]
- Gouveia, C.S.S.; Ganança, J.F.T.; Lebot, V.; de Carvalho, M.Â.A.P. Quantitation of oxalates in corms and shoots of Colocasia esculenta (L.) Schott under drought conditions. Acta Physiol. Plant. 2018, 40, 124. [Google Scholar] [CrossRef]
- Dye, W.B. Chemical studies on Halogeton Glomeratus. Weeds 1956, 1, 55–60. [Google Scholar] [CrossRef]
- AOAC. Micro-chemical Determination of Carbon, Hydrogen, and Nitrogen, Automated Method, Method 972.43. In Official Methods of Analysis; AOAC International Publisher: Arlington, VA, USA, 2000. [Google Scholar]
- Ceballos, H.; Sánchez, T.; Chávez, A.L.; Iglesias, C.; Debouck, D.; Mafla, G.; Tohme, J. Variation in crude protein content in cassava (Manihot esculenta Crantz) roots. J. Food Compos. Anal. 2006, 19, 589–593. [Google Scholar] [CrossRef]
- Ding, X.; Ni, Y.; Kokot, S. NIR spectroscopy and chemometrics for the discrimination of pure, powdered, purple sweet potatoes and their samples adulterated with the white sweet potato flour. Chemom. Intell. Lab. Syst. 2015, 144, 17–23. [Google Scholar] [CrossRef]
- Katoch, R. Analytical Techniques in Biochemistry and Molecular Biology; Springer: New York, NY, USA, 2011; pp. 264, 303, 326. [Google Scholar]
- Van den Boogaard, R.; Kostadinova, S.; Veneklaas, E.; Lambers, H. Association of water use efficiency and nitrogen use efficiency with photosynthetic characteristics of two wheat cultivars. J. Exp. Bot. 1995, 46, 1429–1438. [Google Scholar] [CrossRef]
- Gouveia, C.S.S.; Ganança, J.F.T.; Slaski, J.; Lebot, V.; Pinheiro de Carvalho, M.Â.A. Stable isotope natural abundances (δ13C and δ15N) and carbon-water relations as drought stress mechanism response of taro (Colocasia esculenta L. Schott). J. Plant Physiol. 2019, 232, 100–106. [Google Scholar] [CrossRef]
- Ivlev, A.A. Global redox cycle of biospheric carbon: Interaction of photosynthesis and earth crust processes. BioSystems 2015, 137, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Serret, M.D.; Yousfi, S.; Vicente, R.; Piñero, M.C.; Otálora-Alcón, G.; del Amor, F.M.; Araus, J.L. Interactive effects of CO2 concentration and water regime on stable isotope signatures, nitrogen assimilation and growth in sweet pepper. Front. Plant Sci. 2018, 8, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Duman, F. Uptake of mineral elements during abiotic stress. In Abiotic Stress Responses in Plants: Metabolism, Productivity and Sustainability; Ahmad, P., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 270–271. [Google Scholar]
- Gouveia, C.S.S.; Ganança, J.F.T.; Slaski, J.; Lebot, V.; de Carvalho, M.Â.A.P. Variation of carbon and isotope natural abundances (δ15N and δ13C) of whole-plant sweet potato (Ipomoea batatas L.) subjected to prolonged water stress. J. Plant Physiol. 2019, 243, 153052. [Google Scholar] [CrossRef] [PubMed]
- Zeeman, S.C.; Kossmann, J.; Smith, A.M. Starch: Its metabolism, evolution, and biotechnological modification in plants. Annu. Rev. Plant Biol. 2010, 61, 209–234. [Google Scholar] [CrossRef] [Green Version]
- Gouveia, C.S.S.; Ganança, J.F.T.; Lebot, V.; de Carvalho, M.Â.A.P. Changes in oxalate composition and other nutritive traits in root tubers and shoots of sweet potato (Ipomoea batatas L. [Lam.]) under water stress. J. Sci. Food Agric. 2020, 100, 1702–1710. [Google Scholar] [CrossRef]
- Schadel, W.E.; Walter, W.M., Jr. Calcium oxalate crystals in the roots of sweet potato. J. Am. Soc. Hortic. Sci. 1980, 105, 851–854. [Google Scholar]
- Nakata, P.A. Advances in our understanding of calcium oxalate crystal formation and function in plants. Plant Sci. 2003, 164, 901–909. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Accession ID 1 | Name | Locality | Origin | Category |
|---|---|---|---|---|
| Taro [Colocasia esculenta (L.) Schott] | ||||
| 2056 | Listado | La Palma | Canary Islands—Spain (CAN) | Local |
| 2061 | Blanco Saucero | La Palma | Canary Islands—Spain (CAN) | Local |
| 2210 | Roxo | Porto Moniz | Madeira Island—Portugal (MAD) | Local |
| 2216 | Branco | Santa Cruz | Madeira Island—Portugal (MAD) | Local |
| 2232 | PExPH 15-6 BL/HW/08 | Hawaii | Fiji—Pacific Community (SPC) | Breeding lines |
| 2234 | C3-22 BL/PNG/11 | Papua New Guinea | Fiji—Pacific Community (SPC) | Breeding lines |
| 2239 | Karang CE/MAL/10 | Malaysia | Fiji—Pacific Community (SPC) | Breeding lines |
| Sweet potato [Ipomoea batatas (L.) Lam.] | ||||
| 1036 | Brasileira | Câmara de Lobos | Madeira Island—Portugal (MAD) | Local |
| 1038 | 5 Bicos | Santana | Madeira Island—Portugal (MAD) | Local |
| 2927 | de Flor | Porto Moniz | Madeira Island—Portugal (MAD) | Local |
| 3126 | Inglesa | Porto Moniz | Madeira Island—Portugal (MAD) | Local |
| 2937 | Roja | Tenerife | Canary Islands—Spain (CAN) | Local |
| 2938 | Cubana | Tenerife | Canary Islands—Spain (CAN) | Local |
| 3124 | Vermelha | Bafatá | Guinea-Bissau (GUI) | Local |
| 3125 | Branca | Bafatá | Guinea-Bissau (GUI) | Local |
| Calibration Set | Validation Independent Set | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Taro Models | n | Outliers | Mean | SD | PLS Terms | SEC | SECV | r2cv | RPD | n | r2pred | SEP |
| Moisture, g/100g DW | ||||||||||||
| Corm | 105 | 6 | 7.584 | 1.352 | 9 | 0.592 | 0.633 | 0.783 | 2.14 | 21 | 0.894 | 0.433 |
| Shoot | 102 | 11 | 7.894 | 2.352 | 9 | 1.011 | 1.063 | 0.795 | 2.21 | 18 | 0.838 | 0.837 |
| Protein, g/100g DW | ||||||||||||
| Corm | 105 | 5 | 4.705 | 1.460 | 9 | 0.119 | 0.126 | 0.993 | 11.56 | 21 | 0.995 | 0.191 |
| Shoot | 102 | 4 | 11.542 | 2.145 | 9 | 0.363 | 0.359 | 0.972 | 5.97 | 18 | 0.883 | 0.693 |
| Nitrogen, g/100g DW | ||||||||||||
| Corm | 105 | 3 | 0.754 | 0.231 | 9 | 0.020 | 0.021 | 0.992 | 11.07 | 21 | 0.995 | 0.031 |
| Shoot | 102 | 5 | 1.856 | 0.340 | 9 | 0.059 | 0.059 | 0.970 | 5.79 | 18 | 0.953 | 0.058 |
| Starch, g/100g DW | ||||||||||||
| Corm | 105 | 3 | 47.021 | 5.469 | 9 | 0.982 | 1.048 | 0.964 | 5.22 | 21 | 0.977 | 0.767 |
| Shoot | 102 | 3 | 10.955 | 2.999 | 9 | 0.483 | 0.524 | 0.969 | 5.72 | 18 | 0.984 | 0.805 |
| Crude Fiber, g/100g DW | ||||||||||||
| Corm | 105 | 6 | 0.435 | 0.108 | 9 | 0.019 | 0.019 | 0.968 | 5.59 | 21 | 0.985 | 0.018 |
| Shoot | 102 | 14 | 1.646 | 0.227 | 9 | 0.053 | 0.057 | 0.939 | 4.01 | 18 | 0.980 | 0.031 |
| Minerals, g/100g DW | ||||||||||||
| Corm | 105 | 4 | 4.483 | 1.026 | 9 | 0.170 | 0.176 | 0.970 | 5.83 | 21 | 0.986 | 0.122 |
| Shoot | 102 | 5 | 9.707 | 1.049 | 9 | 0.285 | 0.334 | 0.898 | 3.14 | 18 | 0.950 | 0.245 |
| CaOx, g/100g DW | ||||||||||||
| Corm | 105 | 9 | 0.249 | 0.101 | 9 | 0.027 | 0.028 | 0.922 | 3.58 | 21 | 0.721 | 0.046 |
| Shoot | 102 | 4 | 0.241 | 0.107 | 9 | 0.038 | 0.040 | 0.863 | 2.70 | 18 | 0.498 | 0.056 |
| δ13C, ‰ DW | ||||||||||||
| Corm | 105 | 3 | −26.183 | 0.788 | 9 | 0.250 | 0.285 | 0.868 | 2.76 | 21 | 0.977 | 0.177 |
| Shoot | 102 | 0 | −26.304 | 1.077 | 9 | 0.642 | 0.704 | 0.574 | 1.53 | 18 | 0.873 | 0.420 |
| Δ13C, ‰ DW | ||||||||||||
| Corm | 105 | 3 | 18.662 | 0.830 | 9 | 0.263 | 0.293 | 0.874 | 2.83 | 21 | 0.977 | 0.156 |
| Shoot | 102 | 7 | 18.804 | 1.074 | 9 | 0.503 | 0.552 | 0.740 | 1.94 | 18 | 0.910 | 0.411 |
| δ15N, % DW | ||||||||||||
| Corm | 105 | 0 | 4.634 | 0.864 | 9 | 0.473 | 0.564 | 0.575 | 1.53 | 21 | 0.717 | 0.488 |
| Shoot | 102 | 4 | 0.241 | 0.107 | 8 | 0.038 | 0.040 | 0.863 | 2.70 | 18 | 0.711 | 0.549 |
| Calibration Set | Validation Independent Set | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sweet Potato Models | n | Outliers | Mean | SD | PLS Terms | SEC | SECV | r2cv | RPD | n | r2pred | SEP |
| Moisture, g/100g DW | ||||||||||||
| Tuber | 105 | 7 | 5.066 | 1.539 | 9 | 0.670 | 0.761 | 0.755 | 2.02 | 24 | 0.532 | 0.822 |
| Shoot | 111 | 0 | 6.114 | 1.083 | 9 | 0.616 | 0.701 | 0.584 | 1.54 | 24 | 0.495 | 0.945 |
| Protein, g/100g DW | ||||||||||||
| Tuber | 105 | 9 | 6.038 | 1.262 | 9 | 0.112 | 0.114 | 0.992 | 11.10 | 24 | 0.994 | 0.099 |
| Shoot | 111 | 9 | 14.480 | 3.160 | 9 | 0.218 | 0.244 | 0.994 | 12.96 | 24 | 0.995 | 0.274 |
| Nitrogen, g/100g DW | ||||||||||||
| Tuber | 105 | 11 | 0.973 | 0.203 | 9 | 0.022 | 0.024 | 0.986 | 8.47 | 24 | 0.994 | 0.016 |
| Shoot | 111 | 9 | 2.318 | 0.506 | 9 | 0.035 | 0.039 | 0.994 | 12.96 | 24 | 0.995 | 0.044 |
| Starch, g/100g DW | ||||||||||||
| Tuber | 105 | 3 | 42.509 | 4.122 | 9 | 1.430 | 1.608 | 0.847 | 2.56 | 24 | 0.856 | 0.944 |
| Shoot | 111 | 4 | 4.944 | 2.599 | 9 | 0.487 | 0.517 | 0.961 | 5.03 | 24 | 0.981 | 0.323 |
| Minerals, g/100g DW | ||||||||||||
| Tuber | 105 | 8 | 4.336 | 0.482 | 9 | 0.082 | 0.089 | 0.966 | 5.40 | 24 | 0.868 | 0.158 |
| Shoot | 111 | 8 | 11.266 | 1.800 | 9 | 0.215 | 0.221 | 0.985 | 8.16 | 24 | 0.992 | 0.171 |
| CaOx, g/100g DW | ||||||||||||
| Tuber | 105 | 8 | 0.058 | 0.034 | 9 | 0.012 | 0.014 | 0.826 | 2.41 | 24 | 0.865 | 0.020 |
| Shoot | 111 | 3 | 0.099 | 0.079 | 9 | 0.027 | 0.029 | 0.866 | 2.74 | 24 | 0.960 | 0.014 |
| Total carbon, g/100g DW | ||||||||||||
| Tuber | 105 | 6 | 40.341 | 0.333 | 9 | 0.173 | 0.185 | 0.687 | 1.79 | 24 | 0.705 | 0.176 |
| Shoot | 111 | 6 | 41.510 | 0.572 | 9 | 0.220 | 0.238 | 0.826 | 2.40 | 24 | 0.914 | 0.232 |
| δ13C, ‰ DW | ||||||||||||
| Tuber | 105 | 6 | −25.065 | 1.161 | 9 | 0.306 | 0.364 | 0.901 | 3.19 | 24 | 0.178 | 0.602 |
| Shoot | 111 | 6 | −26.611 | 1.257 | 9 | 0.257 | 0.265 | 0.956 | 4.75 | 24 | 0.579 | 0.311 |
| Δ13C, ‰ DW | ||||||||||||
| Tuber | 105 | 6 | 17.505 | 1.211 | 9 | 0.319 | 0.380 | 0.901 | 3.18 | 24 | 0.629 | 0.174 |
| Shoot | 111 | 6 | 19.121 | 1.314 | 9 | 0.269 | 0.277 | 0.956 | 4.75 | 24 | 0.580 | 0.325 |
| δ15N, % DW | ||||||||||||
| Tuber | 105 | 0 | 4.586 | 1.120 | 9 | 0.533 | 0.584 | 0.727 | 1.92 | 24 | 0.769 | 0.499 |
| Shoot | 111 | 5 | 4.185 | 1.097 | 9 | 0.260 | 0.274 | 0.938 | 4.01 | 24 | 0.788 | 0.345 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouveia, C.S.S.; Lebot, V.; Pinheiro de Carvalho, M. NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours. Appl. Sci. 2020, 10, 8724. https://doi.org/10.3390/app10238724
Gouveia CSS, Lebot V, Pinheiro de Carvalho M. NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours. Applied Sciences. 2020; 10(23):8724. https://doi.org/10.3390/app10238724
Chicago/Turabian StyleGouveia, Carla S. S., Vincent Lebot, and Miguel Pinheiro de Carvalho. 2020. "NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours" Applied Sciences 10, no. 23: 8724. https://doi.org/10.3390/app10238724
APA StyleGouveia, C. S. S., Lebot, V., & Pinheiro de Carvalho, M. (2020). NIRS Estimation of Drought Stress on Chemical Quality Constituents of Taro (Colocasia esculenta L.) and Sweet Potato (Ipomoea batatas L.) Flours. Applied Sciences, 10(23), 8724. https://doi.org/10.3390/app10238724