Evaluation of the Antioxidant and Antimicrobial Activities of Porcine Liver Protein Hydrolysates Obtained Using Alcalase, Bromelain, and Papain

 ,
,  ,
,  ,
,  ,
, 
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Chemical Composition of Porcine Liver
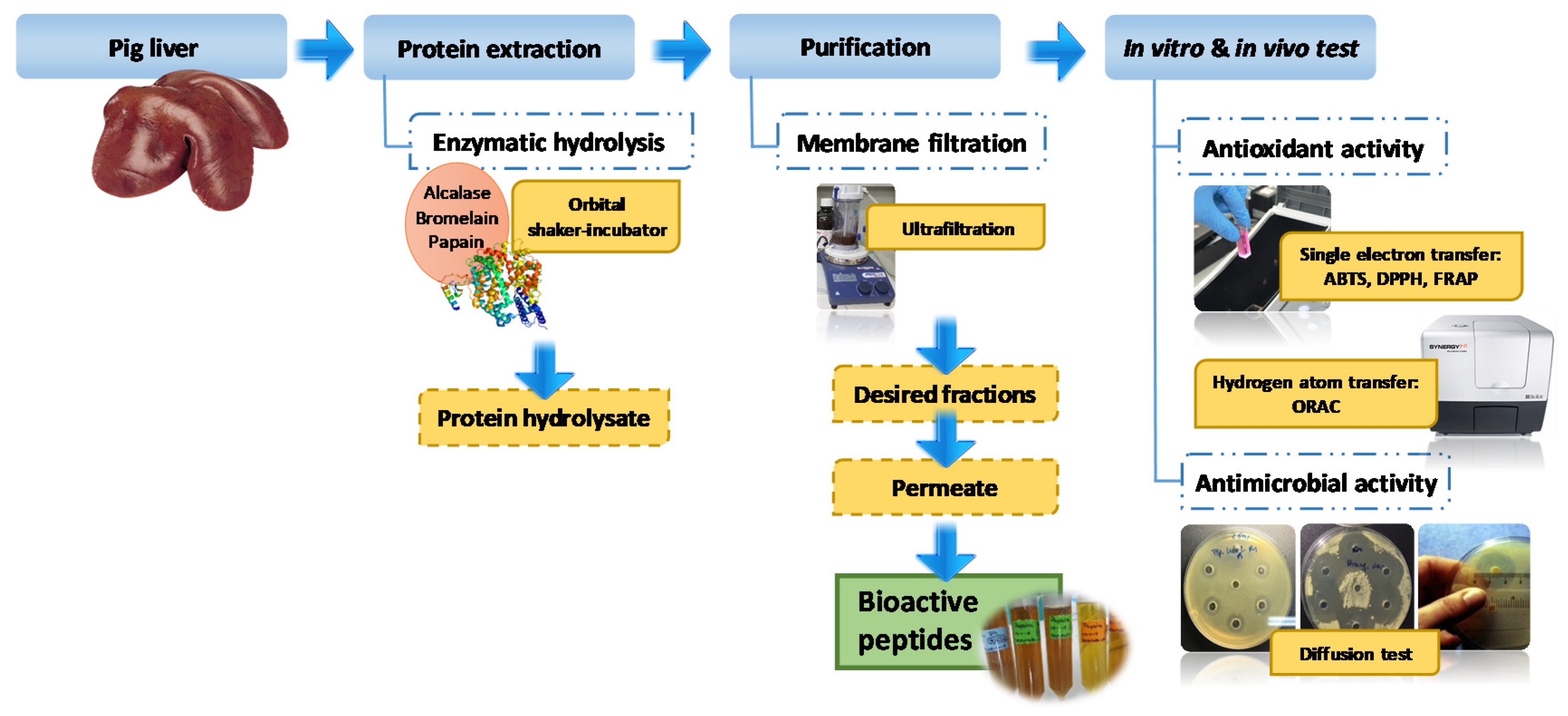
2.3. Peptic Hydrolysis of Porcine Liver
2.4. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Activity
2.5. 2-2′-Azino-di-[3-ethylbenzthiazoline sulfonate] (ABTS) Radical Scavenging Activity
2.6. Ferric Reducing Antioxidant Power Assay (FRAP)
2.7. Oxygen Radical Absorbance Capacity Assay (ORAC)
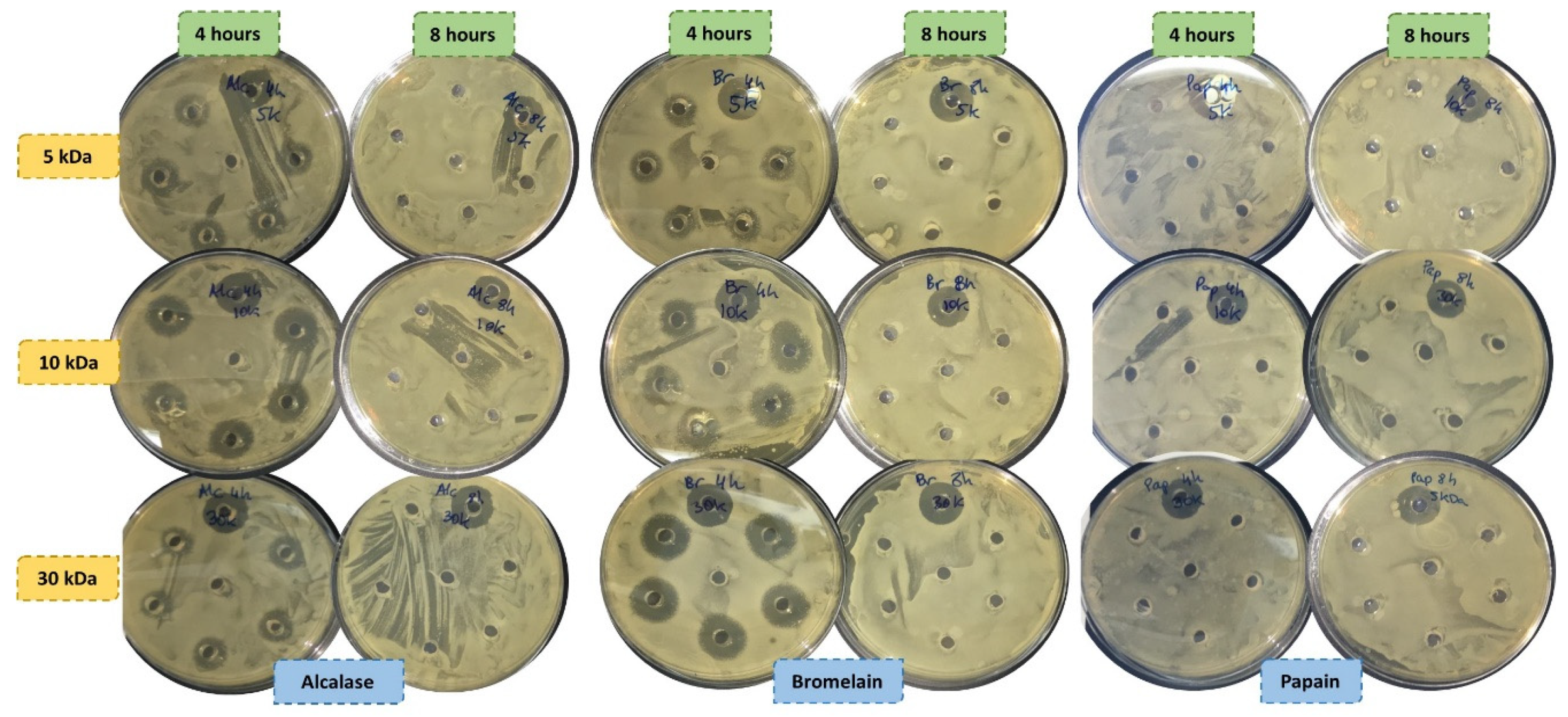
2.8. Antimicrobial Activity
2.9. Statistical Analysis
3. Results
3.1. Chemical Composition
3.2. DPPH Radical Scavenging Activity
3.3. ABTS Radical Scavenging Activity
3.4. Ferric Reducing Antioxidant Power Assay (FRAP)
3.5. Oxygen Radical Absorbance Capacity Assay (ORAC)
3.6. Antimicrobial Activity
4. Discussion
4.1. Chemical Composition
4.2. DPPH Radical Scavenging Activity
4.3. ABTS Radical Scavenging Activity
4.4. Ferric Reducing Antioxidant Power Assay (FRAP)
4.5. Oxygen Radical Absorbance Capacity Assay (ORAC)
4.6. Antimicrobial Activity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lorenzo, J.M.; Sarriés, M.V.; Tateo, A.; Polidori, P.; Franco, D.; Lanza, M. Carcass characteristics, meat quality and nutritional value of horsemeat: A review. Meat Sci. 2014, 96, 1478–1488. [Google Scholar] [CrossRef]
- Siti, R.A.M.; Zainal, S.; Noriham, A. Determination of optimum condition of leucine content in beef protein hydrolysate using response surface methodology. Malays. J. Anal. Sci. 2016, 20, 829–837. [Google Scholar]
- Lorenzo, J.M.; Pateiro, M. Influence of type of muscles on nutritional value of foal meat. Meat Sci. 2013, 93, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Borrajo, P.; Pateiro, M.; Barba, F.J.; Mora, L.; Franco, D.; Toldrá, F.; Lorenzo, J.M. Antioxidant and Antimicrobial Activity of Peptides Extracted from Meat By-products: A Review. Food Anal. Methods 2019, 12, 2401–2415. [Google Scholar] [CrossRef]
- Mullen, A.M.; Álvarez, C.; Zeugolis, D.I.; Henchion, M.; O’Neill, E.; Drummond, L. Alternative uses for co-products: Harnessing the potential of valuable compounds from meat processing chains. Meat Sci. 2017, 132, 90–98. [Google Scholar] [CrossRef]
- Pateiro, M.; Borrajo, P.; Campagnol, P.C.B.; Domínguez, R.; Tomasevic, I.; Munekata, P.E.S.; Barba, F.J.; Lorenzo, J.M. Extraction of valuable compounds from meat by-products. In Green Extraction and Valorization of By-Products from Food Processing; Barba, F.J., Roselló-Soto, E., Brncic, M., Lorenzo, J.M., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 55–90. [Google Scholar]
- Yu, H.C.; Hsu, J.L.; Chang, C.I.; Tan, F.J. Antioxidant properties of porcine liver proteins hydrolyzed using Monascus purpureus. Food Sci. Biotechnol. 2017, 26, 1217–1225. [Google Scholar] [CrossRef]
- Jayathilakan, K.; Sultana, K.; Radhakrishna, K.; Bawa, A.S. Utilization of byproducts and waste materials from meat, poultry and fish processing industries: A review. J. Food Sci. Technol. 2012, 49, 278–293. [Google Scholar] [CrossRef]
- MAPA Porcino. Sectores Ganaderos. Producción y Mercados Ganaderos. Ganadería. Ministerio de Agricultura, Pesca y Alimentación. Gobierno de España. Available online: https://www.mapa.gob.es/es/ganaderia/temas/produccion-y-mercados-ganaderos/sectores-ganaderos/porcino/ (accessed on 24 February 2020).
- Verma, A.K.; Chatli, M.K.; Kumar, P.; Mehta, N. Antioxidant and antimicrobial activity of protein hydrolysate extracted from porcine liver. Indian J. Anim. Sci. 2017, 87, 711–717. [Google Scholar]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-derived bioactive peptides in human health: Challenges and opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef]
- Rai, M.; Pandit, R.; Gaikwad, S.; Kövics, G. Antimicrobial peptides as natural bio-preservative to enhance the shelf-life of food. J. Food Sci. Technol. 2016, 53, 3381–3394. [Google Scholar] [CrossRef]
- Najafian, L.; Babji, A.S. Production of bioactive peptides using enzymatic hydrolysis and identification antioxidative peptides from patin (Pangasius sutchi) sarcoplasmic protein hydolysate. J. Funct. Foods 2014, 9, 280–289. [Google Scholar] [CrossRef]
- Mokni Ghribi, A.; Maklouf Gafsi, I.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, M.; Zou, Y.; Sun, Z.; Xu, W. Optimization of flavourzyme hydrolysis condition for the preparation of antioxidant peptides from duck meat using response surface methodology. J. Poult. Sci. 2018, 55, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Banan-Mwine Daliri, E.; Oh, D.H.; Lee, B.H. Bioactive Peptides—Review. Foods 2017, 6, 1–21. [Google Scholar]
- Bhat, Z.F.; Kumar, S.; Bhat, H.F. Bioactive peptides of animal origin: A review. J. Food Sci. Technol. 2015, 52, 5377–5392. [Google Scholar] [CrossRef]
- Verma, A.K.; Chatli, M.K.; Kumar, P.; Mehta, N. In-vitro assessment of antioxidant and antimicrobial activity of whole porcine-liver hydrolysates and its fractions. Anim. Prod. Sci. 2019, 59, 641–646. [Google Scholar] [CrossRef]
- Ee, K.-Y.; Khoo, L.-Y.; Ng, W.-J.; Wong, F.-C.; Chai, T.-T. Effects of bromelain and trypsin hydrolysis on the phytochemical content, antioxidant activity, and antibacterial activity of roasted butterfly pea seeds. Processes 2019, 7, 534. [Google Scholar] [CrossRef]
- International Standards Meat and Meat Products—Determination of Ash Content; ISO 936; International Organization for Standarization: Geneva, Switzerland, 1998.
- International Standards Meat and Meat Products—Determination of Moisture Content; ISO 1442; International Organization for Standarization: Geneva, Switzerland, 1997.
- International Standards Meat and Meat Products—Determination of Nitrogen Content; ISO 937; International Organization for Standarization: Geneva, Switzerland, 1978.
- AOCS. AOCS Official Procedure Am5-04. Rapid Determination of Oil/Fat Utilizing High Temperature Solvent Extraction; American Oil Chemists Society: Urbana, IL, USA, 2005. [Google Scholar]
- Mishra, K.; Ojha, H.; Chaudhury, N.K. Estimation of antiradical properties of antioxidants using DPPH- assay: A critical review and results. Food Chem. 2012, 130, 1036–1043. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “Antioxidant Power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Huan, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Ramírez Aristizabal, L.S.; Marín, D. Evaluación de la actividad antibacteriana de aceites esenciales y extractos etanólicos utilizando métodos de difusión en agar y dilución en pozo. Sci. Tech. 2012, 17, 152–157. [Google Scholar]
- Alao, B.O.; Falowo, A.B.; Chulayo, A.; Muchenje, V. The potential of animal by-products in food systems: Production, prospects and challenges. Sustainability 2017, 9, 1089. [Google Scholar] [CrossRef]
- Honikel, K.O. Composition and calories. In Handbook of Analysis of Edible Animal By-Products; Nollet, L.M.L., Toldrá, F., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 105–123. [Google Scholar]
- Kakimov, A.; Suychinov, A.; Tsoy, A.; Mustambayev, N.; Ibragimov, N.; Kuderinova, N.; Mirasheva, G.; Yessimbekov, Z. Nutritive and biological value of liver and blood of various slaughtered animals. J. Pharm. Res. Int. 2018, 22, 1–5. [Google Scholar] [CrossRef]
- Seong, P.N.; Cho, S.H.; Park, K.M.; Kang, G.H.; Park, B.Y.; Moon, S.S.; Ba, H. Van Characterization of chicken by-products by mean of proximate and nutritional compositions. Korean J. Food Sci. Anim. Resour. 2015, 35, 179–188. [Google Scholar] [CrossRef]
- Seong, P.N.; Kang, G.H.; Park, K.M.; Cho, S.H.; Kang, S.M.; Park, B.Y.; Moon, S.S.; Ba, H. Van Characterization of Hanwoo bovine by-products by means of yield, physicochemical and nutritional compositions. Korean J. Food Sci. Anim. Resour. 2014, 34, 434–447. [Google Scholar] [CrossRef]
- Seong, P.N.; Park, K.M.; Cho, S.H.; Kang, S.M.; Kang, G.H.; Park, B.Y.; Moon, S.S.; Ba, H. Van Characterization of edible pork by-products by means of yield and nutritional composition. Korean J. Food Sci. Anim. Resour. 2014, 34, 297–306. [Google Scholar] [CrossRef]
- Di Bernardini, R.; Rai, D.K.; Bolton, D.; Kerry, J.; O’Neill, E.; Mullen, A.M.; Harnedy, P.; Hayes, M. Isolation, purification and characterization of antioxidant peptidic fractions from a bovine liver sarcoplasmic protein thermolysin hydrolyzate. Peptides 2011, 32, 388–400. [Google Scholar] [CrossRef]
- Onuh, J.O.; Girgih, A.T.; Aluko, R.E.; Aliani, M. In vitro antioxidant properties of chicken skin enzymatic protein hydrolysates and membrane fractions. Food Chem. 2014, 150, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-K.; Lee, S.-J.; Jeon, B.-T.; Moon, S.-H.; Kim, B.; Park, T.-K.; Han, J.-S.; Park, P.-J. Purification and characterisation of antioxidative peptides from enzymatic hydrolysates of venison protein. Food Chem. 2009, 114, 1365–1370. [Google Scholar] [CrossRef]
- Hidalgo, M.E.; Daroit, D.J.; Folmer Corrêa, A.P.; Pieniz, S.; Brandelli, A.; Risso, P.H. Physicochemical and antioxidant properties of bovine caseinate hydrolysates obtained through microbial protease treatment. Int. J. Dairy Technol. 2012, 65, 342–352. [Google Scholar] [CrossRef]
- Castro-Muñoz, R.; Ruby-Figueroa, R. Membrane technology for the recovery of high-added value compounds from meat processing coproducts. In Sustainable Meat Production and Processing; Galanakis, C.M., Ed.; Academic Press: London, UK, 2019; pp. 127–143. [Google Scholar]
- Liu, R.; Xing, L.; Fu, Q.; Zhou, G.H.; Zhang, W.G. A review of antioxidant peptides derived from meat muscle and by-products. Antioxidants 2016, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Hayes, K.D.; Shahidi, F. Comparative study on antioxidative activity of yellow stripe trevally protein hydrolysate produced from Alcalase and Flavourzyme. Int. J. Food Sci. Technol. 2008, 43, 1019–1026. [Google Scholar] [CrossRef]
- Jamdar, S.N.; Rajalakshmi, V.; Sharma, A. Antioxidant and ace inhibitory properties of poultry viscera protein hydrolysate and its peptide fractions. J. Food Biochem. 2012, 36, 494–501. [Google Scholar] [CrossRef]
- Phanturat, P.; Benjakul, S.; Visessanguan, W.; Roytrakul, S. Use of pyloric caeca extract from bigeye snapper (Priacanthus macracanthus) for the production of gelatin hydrolysate with antioxidative activity. LWT Food Sci. Technol. 2010, 43, 86–97. [Google Scholar] [CrossRef]
- Damgaard, T.D.; Otte, J.A.; Meinert, L.; Jensen, K.; Lametsch, R. Antioxidant capacity of hydrolyzed porcine tissues. Food Sci. Nutr. 2014, 2, 282–288. [Google Scholar] [CrossRef]
- O’Sullivan, S.M.; Lafarga, T.; Hayes, M.; O’Brien, N.M. Bioactivity of bovine lung hydrolysates prepared using papain, pepsin, and Alcalase. J. Food Biochem. 2017, 41, e12406. [Google Scholar] [CrossRef]
- Bah, C.S.F.; Carne, A.; McConnell, M.A.; Mros, S.; Bekhit, A.E.D.A. Production of bioactive peptide hydrolysates from deer, sheep, pig and cattle red blood cell fractions using plant and fungal protease preparations. Food Chem. 2016, 202, 458–466. [Google Scholar] [CrossRef]
- Liu, Q.; Kong, B.; Xiong, Y.L.; Xia, X. Antioxidant activity and functional properties of porcine plasma protein hydrolysate as influenced by the degree of hydrolysis. Food Chem. 2010, 118, 403–410. [Google Scholar] [CrossRef]
- Chang, C.Y.; Wu, K.C.; Chiang, S.H. Antioxidant properties and protein compositions of porcine haemoglobin hydrolysates. Food Chem. 2007, 100, 1537–1543. [Google Scholar] [CrossRef]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K.; Chatli, M.K.; Mehta, N.; Kumar, P. Efficacy of antioxidant and antimicrobial activity of whole porcine blood hydrolysates and its fractions under in-vitro conditions. Anim. Prod. Sci. 2018, 58, 2084–2090. [Google Scholar] [CrossRef]
- Bah, C.S.F.; Bekhit, A.E.D.A.; Carne, A.; McConnell, M.A. Production of bioactive peptide hydrolysates from deer, sheep and pig plasma using plant and fungal protease preparations. Food Chem. 2015, 176, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, N.W.; Wong, G.C.L. Antimicrobial peptides and induced membrane curvature: Geometry, coordination chemistry, and molecular engineering. Curr. Opin. Solid State Mater. Sci. 2013, 17, 151–163. [Google Scholar] [CrossRef]
- Beaussart, A.; El-Kirat-Chatel, S. Microbial adhesion and ultrastructure from the single-molecule to the single-cell levels by atomic force microscopy. Cell Surf. 2019, 5, 100031. [Google Scholar] [CrossRef]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? GMS Hyg. Infect. Control 2017, 12, 1–24. [Google Scholar]



{kind=link}
{kind=link}
| Parameter | g/100 g * |
|---|---|
| Moisture | 74.1 ± 0.12 |
| Protein | 19.0 ± 0.25 |
| Fat | 3.56 ± 0.100 |
| Ash | 1.40 ± 0.010 |
| Carbohydrate | 1.97 ± 0.270 |
| Enzyme | Papain | Bromelain | Alcalase | Enzyme Sig. | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Hydrolysis Time | 4 h | 8 h | Sig. | 4 h | 8 h | Sig. | 4 h | 8 h | Sig. | 4 h | 8 h |
| MWCO | DPPH (µg trolox/g) | ||||||||||
| 5 kDa | 249 ± 2.2 aγ | 304 ± 4.6 Aα | *** | 341 ± 6.1 bα | 322 ± 3.9 Aα | ns | 372 ± 3.7 cα | 443 ± 7.1 Bα | *** | *** | *** |
| 10 kDa | 228 ± 4.1 aβ | 325 ± 3.2 Aβ | *** | 427 ± 3.4 cβ | 379 ± 5.2 Bβ | ** | 408 ± 2.0 bβ | 542 ± 2.7 Cβ | *** | *** | *** |
| 30 kDa | 145 ± 5.4 aα | 309 ± 3.9 Aα | *** | 344 ± 2.6 bα | 376 ± 4.1 Bβ | ** | 477 ± 3.9 cγ | 562 ± 4.5 Cγ | *** | *** | *** |
| Sig. | *** | * | *** | *** | *** | *** | |||||
| ABTS (mg Ascorbic Acid/100 g) | |||||||||||
| 5 kDa | 277 ± 1.9 aβ | 352 ± 4.9 Bα | *** | 346 ± 10.3 bβ | 284 ± 5.7 Aα | ** | 754 ± 8.2 cα | 815 ± 6.7 Cβ | ** | *** | *** |
| 10 kDa | 264 ± 5.1 aα | 368 ± 5.1 Bβ | *** | 392 ± 6.7 bγ | 335 ± 5.4 Aβ | ** | 791 ± 7.2 cβ | 1068 ± 6.1 Cγ | *** | *** | *** |
| 30 kDa | 335 ± 1.9 bγ | 416 ± 1.9 Bγ | *** | 281 ± 1.5 aα | 280 ± 2.9 Aα | ns | 936 ± 4.6 cγ | 761 ± 8.7 Cα | *** | *** | *** |
| Sig. | *** | *** | *** | *** | *** | *** | |||||
| FRAP (µmol Fe+2/100 g) | |||||||||||
| 5 kDa | 24.3 ± 0.72 aα | 46.2 ± 2.47 Aαβ | ** | 37.4 ± 0.54 cα | 57.2 ± 3.88 Bα | ** | 31.9 ± 0.66 bα | 43.3 ± 1.21 Aα | ** | *** | * |
| 10 kDa | 42.8 ± 2.95 aβ | 57.0 ± 3.98 Aβ | * | 50.0 ± 0.15 bβ | 64.4 ± 2.51 Aαβ | ** | 38.6 ± 0.40 aβ | 62.4 ± 0.44 Aαβ | *** | ** | ns |
| 30 kDa | 49.0 ± 5.79 aβ | 36.9 ± 4.29 Aα | ns | 58.3 ± 0.46 aγ | 69.8 ± 1.06 Bβ | *** | 51.2 ± 0.92 aγ | 82.9 ± 1.24 Bβ | ns | ns | * |
| Sig. | ** | * | *** | * | *** | * | |||||
| ORAC (mg trolox/g) | |||||||||||
| 5 kDa | 21.3 ± 0.76 aα | 35.5 ± 3.28 Aα | * | 34.5 ± 0.51 bβ | 40.3 ± 0.13 Aβ | *** | 38.2 ± 1.11 cα | 35.8 ± 1.10 Aα | ns | *** | ns |
| 10 kDa | 34.1 ± 0.25 aβ | 31.7 ± 0.64 Aα | * | 36.4 ± 0.56 aγ | 45.2 ± 0.09 Bγ | *** | 35.7 ± 1.69 aα | 49.6 ± 1.09 Cβ | ** | ns | *** |
| 30 kDa | 37.4 ± 1.90 bβ | 44.3 ± 2.74 Bβ | ns | 31.5 ± 0.48 aα | 28.7 ± 0.90 Aα | ns | 40.1 ± 2.19 bα | 53.2 ± 3.94 Bβ | * | * | ** |
| Sig. | *** | * | ** | *** | ns | ** | |||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borrajo, P.; Pateiro, M.; Gagaoua, M.; Franco, D.; Zhang, W.; Lorenzo, J.M. Evaluation of the Antioxidant and Antimicrobial Activities of Porcine Liver Protein Hydrolysates Obtained Using Alcalase, Bromelain, and Papain. Appl. Sci. 2020, 10, 2290. https://doi.org/10.3390/app10072290
Borrajo P, Pateiro M, Gagaoua M, Franco D, Zhang W, Lorenzo JM. Evaluation of the Antioxidant and Antimicrobial Activities of Porcine Liver Protein Hydrolysates Obtained Using Alcalase, Bromelain, and Papain. Applied Sciences. 2020; 10(7):2290. https://doi.org/10.3390/app10072290
Chicago/Turabian StyleBorrajo, Paula, Mirian Pateiro, Mohammed Gagaoua, Daniel Franco, Wangang Zhang, and José Manuel Lorenzo. 2020. "Evaluation of the Antioxidant and Antimicrobial Activities of Porcine Liver Protein Hydrolysates Obtained Using Alcalase, Bromelain, and Papain" Applied Sciences 10, no. 7: 2290. https://doi.org/10.3390/app10072290
APA StyleBorrajo, P., Pateiro, M., Gagaoua, M., Franco, D., Zhang, W., & Lorenzo, J. M. (2020). Evaluation of the Antioxidant and Antimicrobial Activities of Porcine Liver Protein Hydrolysates Obtained Using Alcalase, Bromelain, and Papain. Applied Sciences, 10(7), 2290. https://doi.org/10.3390/app10072290