Abstract
Halophytes are salt-tolerant plants that can be used to extract dissolved inorganic nutrients from saline aquaculture effluents under a production framework commonly known as Integrated Multi-Trophic Aquaculture (IMTA). Halimione portulacoides (L.) Aellen (common name: sea purslane) is an edible saltmarsh halophyte traditionally consumed by humans living near coastal wetlands and is considered a promising extractive species for IMTA. To better understand its potential for IMTA applications, the present study investigates how artificial lighting and plant density affect its productivity and capacity to extract nitrogen and phosphorous in hydroponic conditions that mimic aquaculture effluents. Plant growth was unaffected by the type of artificial lighting employed—white fluorescent lights vs. blue-white LEDs—but LED systems were more energy-efficient, with a 17% reduction in light energy costs. Considering planting density, high-density units of 220 plants m−2 produced more biomass per unit of area (54.0–56.6 g m−2 day−1) than did low-density units (110 plants m−2; 34.4–37.1 g m−2 day−1) and extracted more dissolved inorganic nitrogen and phosphorus. Overall, H. portulacoides can be easily cultivated hydroponically using nutrient-rich saline effluents, where LEDs can be employed as an alternative to fluorescent lighting and high-density planting can promote higher yields and extraction efficiencies.
1. Introduction
Halophytes are a group of plants characterized by a range of morphological and physiological features that allow them to thrive in brackish and saline environments [1]. Due to these capabilities, they have been increasingly studied for the treatment of eutrophic saline effluents, especially in the context of Integrated Multi-Trophic Aquaculture (IMTA) frameworks [2,3,4]. The major benefit of this integration pertains to the uptake and reuse of wasted nutrients generated within the production system [5,6].
Previous studies, using a variety of halophyte species, demonstrated positive outcomes in growth and extraction efficiency of nitrogen (N) and phosphorous (P) in integrated aquaculture settings [3]. This approach is associated with the principles of the circular economy, where aquaculture waste streams are valorized through the phytoremediation, harvesting and commercialization of plant biomass [7]. Several edible halophyte species can deliver food products with distinct organoleptic and functional properties, including vegetable oils and bioactive compounds [8,9,10,11,12]. Moreover, halophytes’ relatively low-sodium content makes them a suitable alternative that can reduce sodium intake for populations at risk [13,14]. Given their potential socioeconomic and nutritional benefits, continued investigation into the horticultural production of halophytes within integrated aquaculture frameworks is necessary.
This paper focuses on the species Halimione portulacoides (L.) Aellen (a.k.a. sea purslane), an edible halophyte relatively widespread throughout European and Mediterranean saltmarshes with traditional human uses [15] whose aquaculture and nutritional potential has only recently been scientifically explored [16,17,18,19,20,21]. These first studies showed that H. portulacoides has one of the highest productivity rates among studied halophytes in saline hydroponic conditions and can contribute to the substantial removal of dissolved inorganic N and P, up to a rate of approximately 4.0 mg N g−1 and 0.4 mg P g−1 [16].
For an effective introduction of halophytes in integrated aquaculture systems, it is first important to understand the conditions that benefit their production and estimate key variables (e.g., growth rates, nutrient extraction rates) that can help adjust those conditions for cost-efficient solutions. In the present study, the focus lays on two key variables that influence horticultural production: light and planting density [22,23,24,25,26].
Providing optimal and energy-efficient lighting conditions is paramount for commercial plant production [27], and solid-state LED lighting is nowadays considered the most flexible and cost-efficient technology for indoor hydroponics because it provides better control over different light parameters (e.g., spectrum, irradiance, photosynthetically active radiation) [28]. Different light spectra produced by LEDs, particularly red and blue, have been shown to influence the productivity of conventional leafy-green crops, and results suggest a general improvement in performance relative to other types of artificial lighting (e.g., fluorescence) and natural lighting [29,30,31,32,33,34]. Yet, the influence of different light spectra in the development of halophyte crops is still relatively understudied [35,36,37]. In the present study we decided to test a white T5 fluorescent lamp, a common full-spectrum fluorescent light source in hydroponics, against blue-white LED tiles (6 blue and 4 white LEDs) that covered the full-spectrum (white LEDs) but also provided extra blue light (blue LEDs) to inspect potential benefits.
Planting density is known to affect individual plant development, potentially inducing competitive behavior, which can, in turn, affect production outputs [22,25,26]. In the case of halophytes, some species showed reduced individual growth as plant density increased (e.g., Atriplex prostrata Boucher) [38] while others seemed to be relatively unaffected by this variable (e.g., Batis maritima L., Cressa cretica L., Salicornia europaea L., Sesuvium portulacastrum (L.) L.) [39,40,41]. Since different species can have different biomass allocation strategies under crowding conditions, experimental growth trials can provide invaluable information about the appropriate planting densities to maximize whole plant development or promote the growth of target organs, such as fruits, seeds or leaves.
Hydroponic experiments that test halophytes performance for IMTA applications are still limited in the scientific literature and the present study is a contribution to fill this gap. The main goal of this work is to determine the conditions that further benefit the performance of the sea purslane H. portulacoides as an extractive species for IMTA by understanding the influence of lighting conditions and planting density on its vegetative development and nutrient extraction capacity. To achieve this goal, we tested the following null hypotheses: (i) increasing planting density does not affect hydroponic performance, and (ii) shifting white fluorescent lighting to blue-white LED lighting does not affect plant development. The concentration of photosynthetic pigments (chlorophylls and carotenoids) was also quantified to assess potential changes promoted by both types of light, which could have implications for the added value of halophyte products.
2. Materials and Methods
Using a two-way factorial design with two levels, the effects of artificial lighting and planting density on the performance of H. portulacoides extraction units were tested in controlled hydroponic settings rather than real aquaculture settings, to provide better control over the conditions of the aquatic media. Measuring growth performance consisted of recording individual biomass gain every week (one plant per hydroponic unit) and, at the end of the experimental period, recording total biomass yield (whole plant, below- and above-ground biomass), the number of leaves, and stem length at the level of the hydroponic unit. Photosynthetic pigment concentration was also quantified as a proxy measure of the status of the photosynthetic apparatus. Nutrient extraction efficiencies were assessed through measurements and mass–balance calculations of ammonium (NH4–N), oxidized forms of inorganic nitrogen (NOx–N), and orthophosphate (PO4–P) present in the hydroponic solution, at the beginning and the end of each extraction (or remediation) cycle (1 week). The initial concentrations of inorganic nitrogen (N) and phosphorus (P) in each batch of hydroponic solution were approximately 60 mg N L−1 (4.3 mM N) and 3 mg P L−1 (0.1 mM P). Following Custódio et al. [16], these concentrations were not only analogous to those typically found in more intensive aquaculture effluents but also guaranteed non-limiting access to N and P during each extraction cycle. The data generated during this study and presented in the Results section are made available in a spreadsheet as Supplementary Material (File S1).
2.1. Plant Material
Stems of H. portulacoides were collected from wild specimens in Ria de Aveiro (Portugal) coastal lagoon (40°38′04.1″N 8°39′40.0″W) in March 2018. Six hundred cuttings, with 4 nodes each, were obtained from those stems and these were placed in small polyethylene containers filled regularly with Hoagland’s nutrient solution to stimulate root development. The elemental composition of the nutrient solution was: 40 mg Ca L−1, 60 mg K L−1, 16 mg Mg L−1, 56 mg N L−1, 16 mg P L−1, 0.28 mg B L−1, 0.03 mg Cu L−1, 1.12 mg Fe L−1, 0.11 mg Mn L−1, 0.34 mg Mo L−1, 0.13 mg Zn L−1. Cuttings grew under natural light and temperature for 3 months and, in June 2018, fully rooted cuttings were randomly distributed throughout the hydroponic units to initiate the acclimation period. Plants were acclimated to the new hydroponic indoor conditions for 2 weeks and during the second week were progressively adapted to a salinity of 20 ppt (0.5% increments of NaCl every second day).
2.2. Experimental Setup
The growth trial took place over 10 weeks, during which plants were allowed to develop with minimal disturbance on a deep-water culture hydroponics configuration using extruded polystyrene floating-rafts without intermediary harvests during this period. An overview of experimental units under different lighting systems at week 1 and week 9 are displayed in Figure 1.

Figure 1.
Experimental units illuminated with fluorescent lights at week 1 (A) and week 9 (B) & with blue-white LEDs at week 1 (C) and week 9 (D).
The hydroponic units were made of opaque polypropylene boxes (300 × 200 × 170 mm) with an overflow outlet to keep water volume at the 5 L mark. The base for the hydroponic solution was artificial seawater prepared by dissolving commercial Red Sea salt (Red Sea, Cheddar, UK) in freshwater purified by reverse osmosis (V2Pure 360 RO System, TMC, Hertfordshire, UK) until achieving a salinity of 20 ppt. The experimental hydroponic media was a modified version of Hoagland’s solution described previously, where N and P concentrations were modified to resemble realistic values of dissolved inorganic N and P as measured in fish-farming effluents [16]. The detailed nutrient composition of the experimental hydroponic solution is presented in Table S1 (Supplementary Materials).
The experimental design consisted of a two-way factorial design with two levels, resulting in 4 treatments. The levels of artificial lighting were ‘fluorescent lights’ and ‘LEDs’ and the levels of plant density were ‘110 plants m−2’ and ‘220 plants m−2’. Treatment labels were as follows: F110 = fluorescent lights + 110 plants m−2; F220 = fluorescent lights + 220 plants m−2; L110 = LEDs + 110 plants m−2; and L220 = LEDs + 220 plants m−2. Each treatment was assigned to five replicate hydroponic units, resulting in a total of 20 units, each having a surface area of 0.0455 m2. Therefore, the F110 and L110 units were assigned 5 plants each, and the F220 and L220 units were each assigned 10 plants, for a total of 150 plants distributed across all hydroponic units. To do so, 150 fully rooted cuttings of similar weight were selected, identified by a number from 1 to 150 and randomly assigned to the hydroponic units following a randomized sequence of integers (from 1 to 150) generated by a tool provided at https://www.random.org (accessed on 18 June 2018). Each plant was individually photographed and weighed at the beginning of the experiment. The average initial biomass was 6.8 ± 1.8 g plant−1. Afterward, the cuttings were fixed to the floating rafts using natural cotton, to hold each plant in place at the base of the aerial portion. The hydroponic medium was continuously aerated to maintain aerobic conditions, and the hydroponic units were refilled as needed with freshwater purified by reverse osmosis to compensate for evapotranspiration.
The fluorescent light was provided by tubular fluorescent lamps (Philips 54W/830 Min Bipin T5 HO ALTO UNP), while the LED light was provided by solid-state LED lighting tiles (AquaBeam 1500 Ultima NP Ocean Blue Light). Their photometric and colorimetric information is presented in Table 1. Each fluorescent lighting system was composed of two fluorescent lamps and each LED system was composed of three LED tiles. The spectral profiles of artificial lighting sources are presented in Figure S1 as Supplementary Materials. The photoperiod was set at 14 h light: 10 h dark and the photosynthetically active radiation (PAR) was adjusted at the beginning of every week so that all artificial lighting sources delivered identical PAR to the plants. PAR, the fraction of electromagnetic radiation that can be used by plants during photosynthesis, is found within the spectral range of 400–700 nm. It is normally expressed as photosynthetic photon flux density (PPFD, μmol photons m−2 s−1). The PAR values reaching the top of the canopy of stocked plants were measured twice a week with a spherical microquantum sensor (US-SQS/L) connected to a Universal Light Meter ULM-500 (Heinz Walz, Pfullingen, Germany). The average PPFD throughout the experiment was 371.0 ± 12.0 µmol photons m−2 s−1.

Table 1.
Technical information of lighting systems.
Water temperature and pH were measured using a multi-parameter portable meter (ProfiLine pH/Cond 3320, WTW, Weilheim, Germany) and dissolved oxygen was measured by a portable oxygen meter (Oxi 3310, WTW, Weilheim, Germany). Measurements were performed twice a week (day 2 and day 7) and average weekly values are presented in Figure S2 (Supplementary Materials). Overall, hydroponic units displayed an average water temperature of 22.9 ± 0.7 °C, a pH of 7.8 ± 0.2, and a dissolved oxygen concentration of 6.7 ± 0.6 mg L−1 (= 81.1 ± 7.3% saturation).
2.3. Growth Performance
To measure weekly growth, one plant per hydroponic unit (five plants per treatment) was randomly chosen at the beginning of the growth trial, and the total weight was systematically measured at the end of every remediation cycle of 1 week. Only one plant per unit was weighed every week to minimize handling time and avoid stress to the other plants. At the end of the experiment (end of week 10), all plants were individually photographed, divided into roots, stems, and leaves and weighed. The three plant organs (roots, stems, and leaves) were pooled per experimental unit and stored at −80 °C. Image-analysis software (ImageJ 1.51) was used to measure the length of stems and count the leaves.
2.4. Nutrient Extraction Efficiency
The efficiency of H. portulacoides in extracting N and P was determined using a retention time (the amount of time the solution remains in the hydroponic unit) of one week (7 days). The time frame allowed for nutrients to be taken up by the plants was crucial to the performance of a hydroponic unit and the suitable retention time could be highly variable. Previous nutrient extraction studies with halophytes used a wide range of retention times, from 12 h to 5 weeks [20,41,42,43]. Following Custódio et al. [16], a one-week retention time was considered appropriate for the present study.
By the end of each extraction period, water samples from each hydroponic unit were collected and filtered (Whatman GF/C filters) into flasks, and final concentrations of ammonium (NH4–N), oxidized forms of inorganic nitrogen (NOx–N) and orthophosphate (PO4–P) in the hydroponic media were determined using a San++ Continuous Flow Analyzer (Skalar Analytical, Breda, The Netherlands) following Skalar’s standard automated method (NH4–N: modified Berthelot reaction for ammonia determination; NOx-N: Total UV digestible nitrogen/nitrate + nitrite/nitrite; PO4–P: Total UV digestible phosphate/orthophosphate). After sampling, the medium in each unit was renewed with a new batch of its corresponding treatment solution. The real initial concentrations of NH4–N, NOx–N, and PO4–P in every new batch were determined the same way as described above. Nutrient extraction efficiency was estimated (both weekly and in total) for dissolved inorganic nitrogen (DIN–N), calculated as the sum of NH4–N and NOx–N, and dissolved inorganic phosphorus (DIP–P), equivalent to PO4–P.
2.5. Photosynthetic Pigments
Biomass samples were taken from the pool of leaves produced in each hydroponic unit and freeze-dried. Pigments were extracted using 95% cold-buffered methanol (2% ammonium acetate). Before extraction, samples were ground with a mortar and 2–3 mg of the sample were weighed into Eppendorf tubes. Subsequently, 1 mL of extraction solvent was added to each tube, followed by 45 s sonication and 20 min incubation at −20 °C in the dark. The extracts obtained were filtered through 0.2 μm PTFE membrane filters, and 50 μL were injected into an HPLC equipment with an SPD-M20A photodiode array detector (Shimadzu, Kyoto, Japan). The chromatographic separation of pigments was achieved using a Supelcosil C18 column (Sigma-Aldrich, St. Louis, MO, USA) following Cruz et al. [44]. Pigments were identified from absorbance spectra, and retention times and concentrations were calculated using linear regression equations obtained from pure crystalline standards (DHI, Hørsolm, Denmark).
2.6. Statistical Analysis
Statistical analysis was performed using R v3.4.3 (64-bit) with R Studio and statistically significant differences were considered at p < 0.05. A two-way ANOVA was employed to assess the effects of plant density and artificial lighting on outcome variables. Post-hoc Tukey’s HSD test for individual means comparison was used when significance was observed. All data were checked for normality (Shapiro–Wilk test) and homogeneity of variance (Levene’s test). A repeated-measure ANOVA was used to assess treatment differences in cumulative biomass gain and N/P extraction efficiency across time points. The Geenhouse–Geisser correction was applied when the sphericity assumption was violated, and the Bonferroni correction was used when performing multiple pairwise comparisons.
3. Results
3.1. Growth Parameters and Productivity
The initial biomass was 6.80 ± 0.04 g per plant and significant differences in growth parameters were detected after 10 weeks (Table 2). At the individual level, F110 produced significantly (p < 0.05) higher aboveground biomass than L220 and higher stem growth than F220 and L220. At the unit level (pooled individual weights), on the other hand, both F220 and L220 produced significantly higher (p < 0.05) aboveground and belowground biomasses, number of leaves and stem growth than did F110 and L110. The total final biomass in F220 was higher (p < 0.05) than in both L110 and F110; meanwhile, L220 only differed significantly (p < 0.05) from L110. The same trend was observed in total productivity, with F220 expressing the highest productivity rate, at 56.6 ± 14.0 g m−2 day−1. Regarding relative growth rate, the treatments showed similar values even though the low-density treatments had a slightly higher rate on average. Cumulative biomass gains over time (Figure 2) showed a large variance in individual weight across treatments, and, as a result, no statistical main effects of either plant density nor artificial lighting were detected. Nonetheless, a within-subject effect of the variable week was detected [F(76.4,1222.7)= 120.58, p = 2.88 × 10−9, generalized η2 = 0.76].
Table 2.
Growth parameters (mean ± s.d.) of H. portulacoides hydroponic units (n = 5). FW—fresh weight. Different letters (a,b,c) indicate significant differences between treatments (p < 0.05).
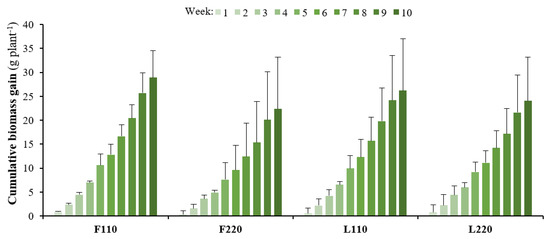
Figure 2.
Individual cumulative weight gain (n = 5). Error bars represent standard deviations.
3.2. Extraction of Dissolved Inorganic N and P
Real initial concentrations of NH4–N were 1.76 ± 0.12 mg N L−1 (0.13 ± 0.01 mM N), of NOx–N were 61.50 ± 4.72 mg N L−1 (4.39 ± 0.34 mM N) and of DIN–N were 63.26 ± 4.82 mg DIN-N L−1 (4.52 ± 0.34 mM N). Orthophosphate concentrations were 3.09 ± 0.15 mg PO4–P L−1 (0.10 ± 0.01 mM P). For each extraction cycle (1 week), the extraction efficiencies were calculated based on the difference between the final and initial concentrations (Figure 3).
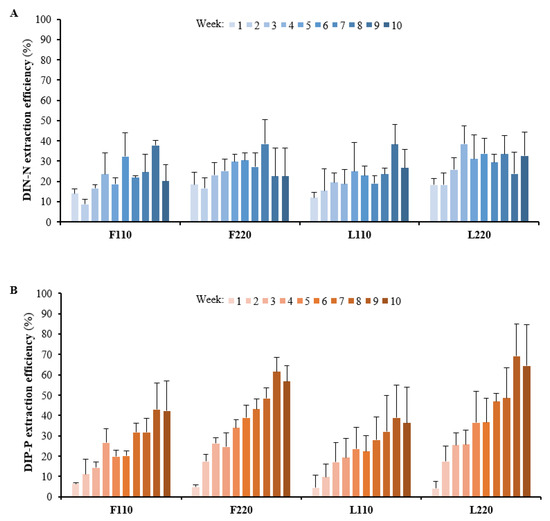
Figure 3.
Extraction efficiency of (A) DIN–N and (B) DIP–P (n = 5). Error bars represent standard deviations.
Using a repeated measures ANOVA, a significant main effect of plant density (F(1,16) = 5.97, p = 0.027, generalized η2 = 0.11), week (F(21.08, 337.24) = 13.86, p = 5.92 × 10−8, generalized η2 = 0.366) and plant density–week interaction (F(21.08, 337.24) = 5.98, p = 4.67 × 10−4, generalized η2 = 0.20) were observed regarding DIN–N extraction efficiencies over time (Figure 3A). Regarding the total quantity of DIN–N extracted during the totality of the experiment (Figure 4A), a main effect of plant density (p < 0.05) was detected: high-density units extracted a total of 875.3 ± 187.7 mg and low-density units extracted in total 686.4 ± 94.2 mg. Overall, high-density units extracted more DIN–N on average than low-density units and extraction efficiencies (%) were: F110 = 21.6 ± 1.8%; F220 = 26.3 ± 6.9%; L110 = 21.8 ± 4.1%; L220 = 29.1 ± 5.1%.
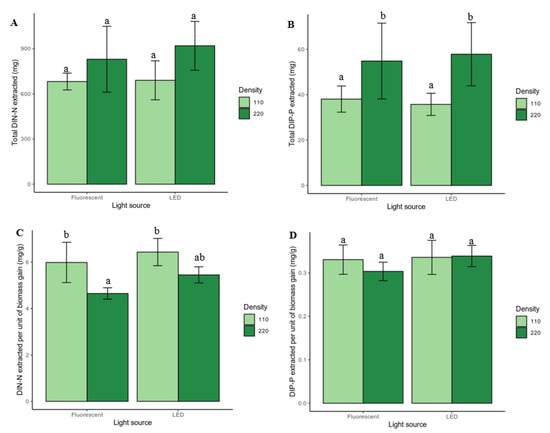
Figure 4.
Total quantity of extracted (A) DIN–N and (B) DIP-P and relative quantity (per biomass gain) of extracted (C) DIN–N and (D) DIP–P (n = 5). Bars represent standard deviations. Different letters (a,b) indicate significant differences between treatments (p < 0.05).
After normalizing the total quantity of DIN–N removed to the total biomass produced (Figure 4C), the main effects of plant density (p < 0.001) and artificial lighting (p < 0.05) were detected: low-density units and LED units resulted in higher DIN–N extraction per gram of biomass produced. Pairwise comparisons show that F220 removed significantly less (p < 0.01) DIN–N per gram of biomass (4.7 ± 0.2 mg g−1) than either F110 (6.0 ± 0.9 mg g−1) or L110 (6.4 ± 0.6 mg g−1), yet the L220 extraction rates (5.5 ± 0.3 mg g−1) were not significantly different from those of low-density units.
Regarding the results of DIP–P extraction efficiency over time (Figure 3B), the significant main effects were also detected fromplant density (F(1,16)= 14.25, p = 0.002, generalized η2 = 0.35), week (F(30.2, 483.2)= 111.60, p = 1.37 × 10−19, generalized η2 = 0.738] and the ‘plant density–week’ interaction (F(30.2, 483.2)= 7.77, p = 4.60 × 10−4, generalized η2 = 0.16]. The total amount of DIP–P extracted (Figure 4B) was also significantly affected by plant density (p < 0.01): high-density units extracted 56.3 ± 14.6 mg and low-density units extracted 36.9 ± 5.2 mg. Pairwise comparisons indicated significantly lower values for F110 and L110 compared with both F220 (p < 0.05) and L220 (p < 0.01). Overall, high-density units extracted more total DIP–P than did the low-density units, and the extraction efficiencies (%) recorded were: F110 = 24.6 ± 3.7%; F220 = 35.5 ± 10.8%; L110 = 23.1 ± 3.2%; L220 = 37.4 ± 9.0%.
Regarding the total DIP–P removed per gram of biomass (Figure 4D), neither main effects nor the treatment effects were detected, and values ranged between 0.30 and 0.34 mg DIP–P extracted per gram of biomass produced.
3.3. Photosynthetic Pigments
The photosynthetic pigments antheraxanthin (Ant), chlorophylls a and b (Chl a, Chl b), 9′cis-neoxanthin (c-Neo), lutein (Lut), violaxanthin (Viola), zeaxanthin (Zea) and β,β-carotene (ββ-Car) were quantified from leaf samples. Pigment concentrations per leaf dry weight (DW) are summarized in Table 3 and statistical analysis suggested that concentrations were not significantly affected by the treatments. However, statistical main effects of plant density on the concentrations of Ant and Zea were detected: concentrations of these two xanthophylls were significantly higher (p < 0.05) in plants growing at low-density (Ant = 38–40 µg g−1 DW; Zea = 42–51 µg g−1 DW) than plants in high-density units (Ant = 32–37 µg g−1 DW; Zea = 36–39 µg g−1 DW).
Table 3.
Photosynthetic pigments concentrations (mean ± sd) in H. portulacoides leaves (n = 5). DW—dry weight.
4. Discussion
4.1. Artificial Lighting
The two tested artificial lighting systems displayed two distinct light spectra that were expected to induce changes in H. portulacoides development. Nonetheless, according to the results, no differences could be associated with the type of lighting system. In contrast, previous studies showed that some types of artificial lighting and their associated light spectra can affect different stages of halophytes’ development. For instance, the seedling germination rates and shoot development of Atriplex halimus L., Atriplex hortensis L., and S. europaea were higher when irradiated with a combined red and blue LED system compared with fluorescent lighting [36], and the vegetative development of A. hortensis also improved under red and blue LEDs [45]. Similarly, the edible Mesembryanthemum crystallinum L. (ice plant) showed improved vegetative development under red and blue LEDs compared to red or blue LEDs alone [35], as well as under red and white LEDs compared with other combinations of white, blue, red, and far-red LEDs [37].
The effect of LED lighting on plant hydroponic production has been more intensively studied using more traditional crops [29,46]. The use of blue and red LEDs (alone or combined) seems to improve the quality and yields of several vegetables and fruit when compared with fluorescent lighting [31]. In aquaponics, for instance, the productivity of kale (B. oleracea) and Swiss chard (Beta vulgaris L.) is higher with LEDs [47] and some studies suggest that blue LEDs seem to stimulate leaf area enlargement and the aboveground development of lettuce (Lactuca sativa L.) and other vegetables (e.g., Chinese cabbage, spinach and coriander) [28,48].
In Table 4, a non-exhaustive summary of hydroponic trials looking at the different effect of LEDs and fluorescent lighting in the vegetative growth of leafy greens is presented, but the results were varied and inconclusive. For instance, L. sativa var. “crispa” grew better under red LEDs compared to either blue or red and blue LEDs [49]; meanwhile, the var. “capitata” seemed to grow better under blue LEDs [50]. The reality is that the effects of light quality in regulating growth processes and physiology in plants is tremendously complex. Even within the same species, outcomes can be different among varieties and time points [51,52].
Table 4.
Hydroponic-based studies of vegetative growth performance and photosynthetic pigment accumulation of leafy greens under different LED spectra and fluorescent lighting. DWC––deep water culture; NFT––nutrient-film technique; FL––fluorescent lighting.
The analysis of pigment concentrations suggested that the photosynthetic pigments in H. portulacoides leaves were unaffected by the type of light spectra after a long-term exposure of 10 weeks. The only indication of an effect came in fact from the planting density, where the concentrations of antheraxanthin and zeaxanthin were higher in low-density units. Being products of the photoprotective xanthophyll cycle [53], one possible explanation for this observation is that leaves from low-density plants were probably exposed to higher irradiance for a longer time-period than high-density plants due to less shading.
Even though the type of light did not seem to affect photosynthetic pigments in H. portulacoides, an increase in the concentration of carotenoids, which include xanthophylls and carotenes, has been observed in leafy vegetables exposed to blue LEDs [29,54]. For instance, several L. sativa varieties, cabbages (Brassica rapa L. and B. oleracea varieties) and water spinach (Ipomoea aquatica Forssk.) displayed an increase in the concentration of chlorophylls and other pigments when growing under blue LEDs [49,55,56,57,58,59]. Nonetheless, some L. sativa varieties displayed similar concentrations regardless of exposure to fluorescent lights or LEDs [60,61], highlighting the complexity of the effects of light quality in plant physiology.
One key aspect to consider when choosing the type of lighting system to cultivate plants in a controlled environment is energy efficiency. In the present study, the wattage of one fluorescent lamp was 54 and one LED unit was 30 W. Therefore, operating on a 14:10 light–dark photoperiod, a fluorescent lamp consumed 0.76 and a LED unit 0.42 kWh day−1. Considering that one fluorescent lighting system was composed of two lamps consuming a total of 1.52 kWh day−1, and one LED lighting system was composed of three LED tiles consuming 1.26 kWh day−1, lighting energy costs were reduced by 17% just by using LEDs.
Operating costs could be driven down even further by employing LEDs in H. portulacoides hydroponic production since their lifespan and maintenance costs are typically lower than fluorescent lights [28]. Despite requiring a higher initial investment, LEDs are likely more cost-efficient given their potentially higher energy efficiency, longer lifespan, and lower maintenance cost [62]. Since H. portulacoides was seemingly unaffected by the type of artificial lighting tested, the LEDs can be seen as a suitable cost-efficient alternative to fluorescent lights.
4.2. Planting Density
Despite promoting lower individual growth, high-density treatments L220 and F220 produced higher yields at the level of the hydroponic unit. Doubling planting density from 110 to 220 plants m−2 increased hydroponic unit productivity 52–57% for an average value of approximately 54–57 g FW m−2 day−1. Higher productivity values were observed by Custódio et al. [16], who reported average yields of 63–73 g FW m−2 day−1 under identical planting densities but by using different combinations of N and P concentrations. On the other hand, Buhmann et al. [42] reported lower productivity values of 33 g FW m−2 day−1 using very different experimental conditions. These first studies with H. portulacoides (Table 5) already suggest contrasting outcomes, most likely influenced by different culture conditions, genetic variability and the use of different ecotypes [64].
Table 5.
Growth and extractive performances of halophyte species under different plant densities. Note: average values and parameters were retrieved, calculated, or estimated from data reported in the referenced publications reported in the methods, tables and/or graphics. A—annual, P—perennial, Aqua—aquaponics, Hydro—hydroponics, CW—constructed wetland.
In other halophyte species, planting density seemed to have had little effect on the hydroponic performance (Table 5). For instance, Webb et al. [41] observed that S. europaea displayed no difference in total productivity between 200 and 10,000 plants m−2. This lack of a plant density effect could be ascribed to the high morphological plasticity of Salicornia plants [41,65]. Still, some species of Salicornia seem to be more productive than others under similar nutrient conditions. For instance, in the same study presented above, S. europaea (200–10,000 plants m−2) produced between 105 and 124 g m−2 day−1 of harvestable biomass, meanwhile Salicornia persica Akhani (100 plants m−2) [66], Salicornia bigelovii Torr. (260 plants m−2) [67] and Salicornia dolichostachya Moss. (38 plants m−2) [42,68] produced on average between 50 and 70 g m−2 day−1.
Salicornia species have annual life cycles, contrary to a perennial plant like H. portulacoides, which indicates major differences in life histories and growth strategies [69]. As such, comparisons between very different species are only meaningful from a horticultural perspective, where some species can be considered better than others in productivity and nutrient extraction rates, for instance. Other edible perennial halophytes investigated for their productivity under different plant densities (Table 5) were S. portulacastrum and B. maritima (92 vs. 184 plants m−2), which displayed no differences between density levels [39]. Sarcocornia ambigua (Michx.) M.A.Alonso & M.B.Crespo, a perennial Salicornioideae, displayed productivity values of 110 g m−2 day−1 when planted at a density of 100 plants m−2 [70]. Yet, much lower values have been reported for the same species (11 g m−2 day−1) at densities of 40 plants m−2 under different aquaponic conditions [71].
Increasing planting density also affected DIN and DIP extraction efficiencies, as high-density units extracted more in total, with average extraction efficiencies of 28% DIN and 36% DIP. Under similar nutrient conditions and using the same fluorescent lighting, high-density and retention time, Custódio et al. [16] reported extraction efficiencies of 35% DIN and 32% DIP, values very close to the present study. However, using radically different hydroponic conditions can produce very different outcomes as shown in previous studies where H. portulacoides displayed extraction efficiencies of 50% DIN and 45% DIP [42] and 65% DIN and 0% DIP [20]. This stresses the fact that culture parameters must be fine-tuned for the specific conditions of a hydroponic or IMTA system, based on the nutrient availability and nutrient extraction rates.
Interestingly, Boxman et al. [39] observed an improvement in DIN extraction efficiency by B. maritima when planted at a low density compared with high-density units; meanwhile, Webb et al. [41] reported no changes in extraction efficiencies after a 50-fold increase in S. europaea planting density, which demonstrated that increasing density did not always equate with an increase in nutrient extraction capacity and that resource competition in plants is complex [72].
The type of substrate used to grow halophytes is also a key factor that influences the performance of a hydroponic unit [73]. Most studies mentioned so far used some type of inert solid substrate (e.g., quarry sand, coconut fiber, expanded clay, perlite) that substantially increased the area available for microbial communities to establish and influence nutrient dynamics in water through complementary processes such as denitrification and adsorption. Confounding factors that affect N and P dynamics are still present in systems using soilless media, as in the present study, such as aeration, water mixing and the presence of microorganisms in the rhizosphere and biofilms [74]. The retention time (exctraction cycle) is another important factor that enables these processes [75] and is a key parameter to consider in these types of studies.
The optimization of hydroponic conditions for halophyte production is indeed very complex due to the interplay of many different environmental variables. Therefore, carefully deliberated experimental designs are key to controlling the many confounding factors present in these systems, which become even more complex in real, rather than simulated, integrated aquaculture settings.
5. Conclusions
As edible halophytes continue to reveal their potential as crops with a role to play in the future of sustainable food production, the conditions for their commercial cultivation must continue to be explored. This is especially true in the context of integrated saline aquaculture where their role could be i) as phytoremediation units that can recover wasted dissolved nutrients and ii) as cash-crops that are nutritional, material and energy sources. By shedding light on the most suitable hydroponic conditions for growing H. portulacoides, future halophyte producers can make informed decisions that translate into sustainable and profitable cultures. Halimione portulacoides displayed productivity values at the higher end of those exhibited by other edible halophytes, given that appropriate planting density and nutritional conditions were present. Potential biomass allocation trade-offs should nonetheless be taken into consideration since certain densities can promote undesirable phenotypes. Regarding artificial lighting. H. portulacoides grew similarly under white fluorescent lighting and blue-white LED lighting, suggesting that the LEDs can be more cost-efficient solution given their similar performance and have potentially lower operating costs. Future trials should continue to explore how different LED spectral profiles can improve development and stimulate the accumulation of bioactive compounds to add functional value to halophyte products. It is also crucial to promote the design and engineering of integrated experimental and commercial setups and the standardized reporting of results to allow a more reliable comparison between hydroponics/IMTA studies and provide more robust data to help determine which halophytes are the most suitable for integrated aquaculture frameworks.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/app11114995/s1, Figure S1: Experimental growth systems with A) fluorescent lights and B) LEDs, Figure S2: Average water A) pH, B) temperature and C) dissolved oxygen, Table S1: Chemical composition of the hydroponic medium. The data generated and presented in this study is made available as supplementary dataset(File S1) in the form of an Excel file, namely growth data, nutrient extraction data and photosynthetic pigments data
Author Contributions
M.C., R.C., and A.I.L. designed the experiment. M.C. conducted the growth trials. M.C., P.C., and A.I.L. conducted the laboratory analysis. M.C. and P.C. analyzed the data. M.C. performed the statistical analysis. M.C. wrote the manuscript. P.C., S.V., R.C., and A.I.L. reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The authors thank the Portuguese Foundation for Science and Technology (FCT) for the financial support to Marco Custódio (PD/BD/127990/2016), Paulo Cartaxana (CEECIND/01434/2018) and CESAM (UIDB/50017/2020+UIDP/50017/2020). This work was also supported by the Integrated Program of SR&TD “Smart Valorization of Endogenous Marine Biological Resources Under a Changing Climate” (reference Centro-01-0145-FEDER-000018), co-funded by Centro 2020 program, Portugal 2020 and European Union, through the European Regional Development Fund, and by the project “AquaMMIn—Development and validation of a modular integrated multitrophic aquaculture system for marine and brackish water species” (MAR-02.01.01-FEAMP-0038) co-funded by Portugal 2020 and the European Union through Mar 2020, the Operational Programme (OP) for the European Maritime and Fisheries Fund (EMFF) in Portugal.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are made available as supplementary material.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Chopin, T.; Robinson, S.M.C.; Troell, M.; Neori, A.; Buschmann, A.H.; Fang, J. Multitrophic Integration for Sustainable Marine Aquaculture. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 2463–2475. ISBN 978-0-08-045405-4. [Google Scholar]
- Custódio, M.; Villasante, S.; Cremades, J.; Calado, R.; Lillebø, A.I. Unravelling the Potential of Halophytes for Marine Integrated Multi-Trophic Aquaculture (IMTA)—A Perspective on Performance, Opportunities and Challenges. Aquac. Environ. Interact. 2017, 9, 445–460. [Google Scholar] [CrossRef]
- Gunning, D.; Maguire, J.; Burnell, G. The Development of Sustainable Saltwater-Based Food Production Systems: A Review of Established and Novel Concepts. Water 2016, 8, 598. [Google Scholar] [CrossRef]
- Goddek, S.; Joyce, A.; Kotzen, B.; Burnell, G.M. Aquaponics Food Production Systems: Combined Aquaculture and Hydroponic Production Technologies for the Future; Springer: Cham, Switzerland, 2019; ISBN 978-3-030-15942-9. [Google Scholar]
- Martins, C.I.M.; Eding, E.H.; Verdegem, M.C.J.; Heinsbroek, L.T.N.; Schneider, O.; Blancheton, J.P.; d’Orbcastel, E.R.; Verreth, J.A.J. New Developments in Recirculating Aquaculture Systems in Europe: A Perspective on Environmental Sustainability. Aquac. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef]
- Ruiz-Salmón, I.; Margallo, M.; Laso, J.; Villanueva-Rey, P.; Mariño, D.; Quinteiro, P.; Dias, A.C.; Nunes, M.L.; Marques, A.; Feijoo, G.; et al. Addressing Challenges and Opportunities of the European Seafood Sector under a Circular Economy Framework. Curr. Opin. Environ. Sci. Health 2020, 13, 101–106. [Google Scholar] [CrossRef]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; da Silva, M.M.; Varela, J.; Custódio, L. Halophytes: Gourmet Food with Nutritional Health Benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal Halophytes: Potent Source of Health Promoting Biomolecules with Medical, Nutraceutical and Food Applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef]
- Loconsole, D.; Cristiano, G.; De Lucia, B. Glassworts: From Wild Salt Marsh Species to Sustainable Edible Crops. Agriculture 2019, 9, 14. [Google Scholar] [CrossRef]
- Maciel, E.; Costa Leal, M.; Lillebø, A.I.; Domingues, P.; Domingues, M.R.; Calado, R. Bioprospecting of Marine Macrophytes Using MS-Based Lipidomics as a New Approach. Mar. Drugs 2016, 14, 49. [Google Scholar] [CrossRef]
- Sharma, R.; Wungrampha, S.; Singh, V.; Pareek, A.; Sharma, M.K. Halophytes As Bioenergy Crops. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- WHO. Guideline: Sodium Intake for Adults and Children; World Health Organization: Geneva, Switzerland, 2012; ISBN 978-92-4-150483-6. [Google Scholar]
- WHO; Regional Office for Europe. European Food and Nutrition Action Plan 2015–2020; World Health Organization: Geneva, Switzerland; Regional Office for Europe: Copenhagen, Denmark, 2015; ISBN 978-92-890-5123-1. [Google Scholar]
- Zanella, L.; Vianello, F. Functional Food from Endangered Ecosystems: Atriplex portulacoides as a Case Study. Foods 2020, 9, 1533. [Google Scholar] [CrossRef]
- Custódio, M.; Villasante, S.; Calado, R.; Lillebø, A.I. Testing the Hydroponic Performance of the Edible Halophyte Halimione portulacoides, a Potential Extractive Species for Coastal Integrated Multi-Trophic Aquaculture. Sci. Total Environ. 2020, 144378. [Google Scholar] [CrossRef]
- Custódio, M.; Maciel, E.; Domingues, M.R.; Lillebø, A.I.; Calado, R. Nutrient Availability Affects the Polar Lipidome of Halimione portulacoides Leaves Cultured in Hydroponics. Sci. Rep. 2020, 10, 6583. [Google Scholar] [CrossRef]
- Maciel, E.; Domingues, P.; Domingues, M.R.M.; Calado, R.; Lillebø, A. Halophyte Plants from Sustainable Marine Aquaponics Are a Valuable Source of Omega-3 Polar Lipids. Food Chem. 2020, 320, 126560. [Google Scholar] [CrossRef] [PubMed]
- Maciel, E.; Lillebø, A.; Domingues, P.; da Costa, E.; Calado, R.; Domingues, M.R.M. Polar Lipidome Profiling of Salicornia ramosissima and Halimione portulacoides and the Relevance of Lipidomics for the Valorization of Halophytes. Phytochemistry 2018, 153, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Marques, B.; Calado, R.; Lillebø, A.I. New Species for the Biomitigation of a Super-Intensive Marine Fish Farm Effluent: Combined Use of Polychaete-Assisted Sand Filters and Halophyte Aquaponics. Sci. Total Environ. 2017, 599–600, 1922–1928. [Google Scholar] [CrossRef]
- Martins-Noguerol, R.; Cambrollé, J.; Mancilla-Leytón, J.M.; Puerto-Marchena, A.; Muñoz-Vallés, S.; Millán-Linares, M.C.; Millán, F.; Martínez-Force, E.; Figueroa, M.E.; Pedroche, J.; et al. Influence of Soil Salinity on the Protein and Fatty Acid Composition of the Edible Halophyte Halimione portulacoides. Food Chem. 2021, 352, 129370. [Google Scholar] [CrossRef]
- Cha, M.-K.; Jeon, Y.A.; Son, J.E.; Cho, Y.-Y. Development of Planting-Density Growth Harvest (PGH) Charts for Quinoa (Chenopodium quinoa Willd.) and Sowthistle (Ixeris dentata Nakai) Grown Hydroponically in Closed-Type Plant Production Systems. Hortic. Environ. Biotechnol. 2016, 57, 213–218. [Google Scholar] [CrossRef]
- van Gelderen, K.; Kang, C.; Pierik, R. Light Signaling, Root Development, and Plasticity. Plant Physiol. 2018, 176, 1049–1060. [Google Scholar] [CrossRef]
- Ouzounis, T.; Rosenqvist, E.; Ottosen, C.-O. Spectral Effects of Artificial Light on Plant Physiology and Secondary Metabolism: A Review. HortScience 2015, 50, 1128–1135. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass Allocation to Leaves, Stems and Roots: Meta-Analyses of Interspecific Variation and Environmental Control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Planting Density and Site Effects on Stem Dimensions, Stand Productivity, Biomass Partitioning, Carbon Stocks and Soil Nutrient Supply in Hybrid Poplar Plantations. Forests 2018, 9, 293. [Google Scholar] [CrossRef]
- Jones, M.A. Using Light to Improve Commercial Value. Hortic. Res. 2018, 5, 47. [Google Scholar] [CrossRef]
- Viršilė, A.; Olle, M.; Duchovskis, P. LED Lighting in Horticulture. In Light Emitting Diodes for Agriculture: Smart Lighting; Dutta Gupta, S., Ed.; Springer: Singapore, 2017; pp. 113–147. ISBN 978-981-10-5807-3. [Google Scholar]
- Alrifai, O.; Hao, X.; Marcone, M.F.; Tsao, R. Current Review of the Modulatory Effects of LED Lights on Photosynthesis of Secondary Metabolites and Future Perspectives of Microgreen Vegetables. J. Agric. Food Chem. 2019, 67, 6075–6090. [Google Scholar] [CrossRef]
- Cammarisano, L.; Donnison, I.S.; Robson, P.R.H. The Effect of Red & Blue Rich LEDs vs. Fluorescent Light on Lollo Rosso Lettuce Morphology and Physiology. Front. Plant Sci. 2021, 12, 603411. [Google Scholar] [CrossRef]
- Hasan, M.M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef]
- Kook, H.-S.; Park, S.-H.; Jang, Y.-J.; Lee, G.-W.; Kim, J.S.; Kim, H.M.; Oh, B.-T.; Chae, J.-C.; Lee, K.-J. Blue LED (Light-Emitting Diodes)-Mediated Growth Promotion and Control of Botrytis Disease in Lettuce. Acta Agric. Scand. B Soil Plant Sci. 2013, 63, 271–277. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-Scale Vegetable Production and the Rise of Microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Heydarizadeh, P.; Zahedi, M.; Boroomand, A.; Agharokh, M.; Sahba, M.R.; Schoefs, B. High Performance of Vegetables, Flowers, and Medicinal Plants in a Red-Blue LED Incubator for Indoor Plant Production. Agron. Sustain. Dev. 2014, 34, 879–886. [Google Scholar] [CrossRef]
- He, J. Integrated Vertical Aeroponic Farming Systems for Vegetable Production in Space Limited Environments. Acta Hortic. 2017, 25–36. [Google Scholar] [CrossRef]
- Sanoubar, R.; Calone, R.; Noli, E.; Barbanti, L. Data on Seed Germination Using LED versus Fluorescent Light under Growth Chamber Conditions. Data Brief 2018, 19, 594–600. [Google Scholar] [CrossRef]
- Weeplian, T.; Yen, T.-B.; Ho, Y.-S. Growth, Development, and Chemical Constituents of Edible Ice Plant (Mesembryanthemum crystallinum L.) Produced under Combinations of Light-Emitting Diode Lights. HortScience 2018, 53, 865–874. [Google Scholar] [CrossRef]
- Wang, L.-W.; Showalter, A.M.; Ungar, I.A. Effects of Intraspecific Competition on Growth and Photosynthesis of Atriplex prostrata. Aquat. Bot. 2005, 83, 187–192. [Google Scholar] [CrossRef]
- Boxman, S.E.; Nystrom, M.; Capodice, J.C.; Ergas, S.J.; Main, K.L.; Trotz, M.A. Effect of Support Medium, Hydraulic Loading Rate and Plant Density on Water Quality and Growth of Halophytes in Marine Aquaponic Systems. Aquac. Res. 2017, 48, 2463–2477. [Google Scholar] [CrossRef]
- Khan, M.A.; Aziz, S. Some Aspects of Salinity, Plant Density, and Nutrient Effects on Cressa cretica L. J. Plant Nutr. 1998, 21, 769–784. [Google Scholar] [CrossRef][Green Version]
- Webb, J.M.; Quintã, R.; Papadimitriou, S.; Norman, L.; Rigby, M.; Thomas, D.N.; Le Vay, L. The Effect of Halophyte Planting Density on the Efficiency of Constructed Wetlands for the Treatment of Wastewater from Marine Aquaculture. Ecol. Eng. 2013, 61, 145–153. [Google Scholar] [CrossRef]
- Buhmann, A.K.; Waller, U.; Wecker, B.; Papenbrock, J. Optimization of Culturing Conditions and Selection of Species for the Use of Halophytes as Biofilter for Nutrient-Rich Saline Water. Agric. Water Manag. 2015, 149, 102–114. [Google Scholar] [CrossRef]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-Goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of Seawater Concentration on the Productivity and Nutritional Value of Annual Salicornia and Perennial Sarcocornia Halophytes as Leafy Vegetable Crops. Sci. Hortic. 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Cruz, S.; Calado, R.; Serôdio, J.; Jesus, B.; Cartaxana, P. Pigment Profile in the Photosynthetic Sea Slug Elysia viridis (Montagu, 1804). J. Mollus. Stud. 2014, 80, 475–481. [Google Scholar] [CrossRef][Green Version]
- Izzo, L.G.; Arena, C.; De Micco, V.; Capozzi, F.; Aronne, G. Light Quality Shapes Morpho-Functional Traits and Pigment Content of Green and Red Leaf Cultivars of Atriplex hortensis. Sci. Hortic. 2019, 246, 942–950. [Google Scholar] [CrossRef]
- Mitchell, C.A.; Dzakovich, M.P.; Gomez, C.; Lopez, R.; Burr, J.F.; Hernández, R.; Kubota, C.; Currey, C.J.; Meng, Q.; Runkle, E.S.; et al. Light-Emitting Diodes in Horticulture. In Horticultural Reviews; Janick, J., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; Volume 43, pp. 1–88. ISBN 978-1-119-10778-1. [Google Scholar]
- Oliver, L.P.; Coyle, S.D.; Bright, L.A.; Shultz, R.C.; Hager, J.V.; Tidwell, J.H. Comparison of Four Artificial Light Technologies for Indoor Aquaponic Production of Swiss Chard, Beta vulgaris, and Kale, Brassica oleracea. J. World Aquacult. Soc. 2018, 49, 837–844. [Google Scholar] [CrossRef]
- Amoozgar, A.; Mohammadi, A.; Sabzalian, M.R. Impact of Light-Emitting Diode Irradiation on Photosynthesis, Phytochemical Composition and Mineral Element Content of Lettuce Cv. Grizzly. Photosynthetica 2017, 55, 85–95. [Google Scholar] [CrossRef]
- Chen, X.; Guo, W.; Xue, X.; Wang, L.; Qiao, X. Growth and Quality Responses of ‘Green Oak Leaf’ Lettuce as Affected by Monochromic or Mixed Radiation Provided by Fluorescent Lamp (FL) and Light-Emitting Diode (LED). Sci. Hortic. 2014, 172, 168–175. [Google Scholar] [CrossRef]
- Namgyel, T.; Khunarak, C.; Siyang, S.; Pobkrut, T.; Norbu, J.; Kerdcharoen, T. Effects of Supplementary LED Light on the Growth of Lettuce in a Smart Hydroponic System. In Proceedings of the 10th International Conference on Knowledge and Smart Technology (KST 2018), Chiang Mai, Thailand, 31 January–3 February 2018; pp. 216–220. [Google Scholar]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of Light Quality on the Accumulation of Phytochemicals in Vegetables Produced in Controlled Environments: A Review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Bugbee, B. Toward an Optimal Spectral Quality for Plant Growth and Development: The Importance of Radiation Capture. Acta Hortic. 2016, 1–12. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W. The Role of Xanthophyll Cycle Carotenoids in the Protection of Photosynthesis. Trends Plant Sci. 1996, 1, 21–26. [Google Scholar] [CrossRef]
- Metallo, R.M.; Kopsell, D.A.; Sams, C.E.; Bumgarner, N.R. Influence of Blue/Red vs. White LED Light Treatments on Biomass, Shoot Morphology, and Quality Parameters of Hydroponically Grown Kale. Sci. Hortic. 2018, 235, 189–197. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hashida, S.; Yoshihara, T. Blue Light-Emitting Diode Light Irradiation of Seedlings Improves Seedling Quality and Growth after Transplanting in Red Leaf Lettuce. HortScience 2010, 45, 1809–1814. [Google Scholar] [CrossRef]
- Kitayama, M.; Nguyen, D.T.P.; Lu, N.; Takagaki, M. Effect of Light Quality on Physiological Disorder, Growth, and Secondary Metabolite Content of Water Spinach (Ipomoea aquatica Forsk) Cultivated in a Closed-Type Plant Production System. Korean J. Hortic. Sci. Technol. 2019, 37, 206–218. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Barickman, T.C.; Morrow, R.C. Sprouting Broccoli Accumulate Higher Concentrations of Nutritionally Important Metabolites under Narrow-Band Light-Emitting Diode Lighting. J. Am. Soc. Hortic. Sci. 2014, 139, 469–477. [Google Scholar] [CrossRef]
- Meng, Q.; Kelly, N.; Runkle, E.S. Substituting Green or Far-Red Radiation for Blue Radiation Induces Shade Avoidance and Promotes Growth in Lettuce and Kale. Environ. Exp. Bot. 2019, 162, 383–391. [Google Scholar] [CrossRef]
- Wang, J.; Lu, W.; Tong, Y.; Yang, Q. Leaf Morphology, Photosynthetic Performance, Chlorophyll Fluorescence, Stomatal Development of Lettuce (Lactuca sativa L.) Exposed to Different Ratios of Red Light to Blue Light. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef]
- Lin, K.; Huang, Z.; Xu, Y. Influence of Light Quality and Intensity on Biomass and Biochemical Contents of Hydroponically Grown Lettuce. HortScience 2018, 53, 1157–1163. [Google Scholar] [CrossRef]
- Lin, K.-H.; Huang, M.-Y.; Huang, W.-D.; Hsu, M.-H.; Yang, Z.-W.; Yang, C.-M. The Effects of Red, Blue, and White Light-Emitting Diodes on the Growth, Development, and Edible Quality of Hydroponically Grown Lettuce (Lactuca sativa L. Var. Capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for Energy Efficient Greenhouse Lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef]
- Zhang, X.; He, D.; Niu, G.; Yan, Z.; Song, J. Effects of Environment Lighting on the Growth, Photosynthesis, and Quality of Hydroponic Lettuce in a Plant Factory. Int. J. Agric. Biol. Eng. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Duarte, B.; Silva, H.; Dias, J.M.; Sleimi, N.; Marques, J.C.; Caçador, I. Functional and Ecophysiological Traits of Halimione portulacoides and Sarcocornia perennis Ecotypes in Mediterranean Salt Marshes under Different Tidal Exposures. Ecol. Res. 2018, 33, 1145–1156. [Google Scholar] [CrossRef]
- Davy, A.J.; Bishop, G.F.; Costa, C.S.B. Salicornia L. (Salicornia pusilla J. Woods, S. ramosissima J. Woods, S. europaea L., S. obscura P.W. Ball & Tutin, S. nitens P.W. Ball & Tutin, S. fragilis P.W. Ball & Tutin and S. dolichostachya Moss). J. Ecol. 2001, 89, 681–707. [Google Scholar] [CrossRef]
- Shpigel, M.; Ben-Ezra, D.; Shauli, L.; Sagi, M.; Ventura, Y.; Samocha, T.; Lee, J.J. Constructed Wetland with Salicornia as a Biofilter for Mariculture Effluents. Aquaculture 2013, 412–413, 52–63. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Potential of Producing Salicornia bigelovii Hydroponically as a Vegetable at Moderate NaCl Salinity. HortScience 2014, 49, 1154–1157. [Google Scholar] [CrossRef]
- Waller, U.; Buhmann, A.K.; Ernst, A.; Hanke, V.; Kulakowski, A.; Wecker, B.; Orellana, J.; Papenbrock, J. Integrated Multi-Trophic Aquaculture in a Zero-Exchange Recirculation Aquaculture System for Marine Fish and Hydroponic Halophyte Production. Aquacult. Int. 2015, 23, 1473–1489. [Google Scholar] [CrossRef]
- Smaje, C. The Strong Perennial Vision: A Critical Review. Agroecol. Sustain. Food Syst. 2015, 39, 471–499. [Google Scholar] [CrossRef]
- Pinheiro, I.; Arantes, R.; do Espírito Santo, C.M.; do Nascimento Vieira, F.; Lapa, K.R.; Gonzaga, L.V.; Fett, R.; Barcelos-Oliveira, J.L.; Seiffert, W.Q. Production of the Halophyte Sarcocornia ambigua and Pacific White Shrimp in an Aquaponic System with Biofloc Technology. Ecol. Eng. 2017, 100, 261–267. [Google Scholar] [CrossRef]
- Pinheiro, I.; Carneiro, R.F.S.; Vieira, F. do N.; Gonzaga, L.V.; Fett, R.; Costa, A.C. de O.; Magallón-Barajas, F.J.; Seiffert, W.Q. Aquaponic Production of Sarcocornia ambigua and Pacific White Shrimp in Biofloc System at Different Salinities. Aquaculture 2020, 519, 734918. [Google Scholar] [CrossRef]
- Craine, J.M.; Dybzinski, R. Mechanisms of Plant Competition for Nutrients, Water and Light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Lüderitz, V.; Gerlach, F. Phosphorus Removal in Different Constructed Wetlands. Acta Biotechnol. 2002, 22, 91–99. [Google Scholar] [CrossRef]
- Wongkiew, S.; Hu, Z.; Chandran, K.; Lee, J.W.; Khanal, S.K. Nitrogen Transformations in Aquaponic Systems: A Review. Aquac. Eng. 2017, 76, 9–19. [Google Scholar] [CrossRef]
- Endut, A.; Jusoh, A.; Ali, N.; Wan Nik, W.B.; Hassan, A. A Study on the Optimal Hydraulic Loading Rate and Plant Ratios in Recirculation Aquaponic System. Bioresour. Technol. 2010, 101, 1511–1517. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).