Abstract
The use of seawater in horticulture is underestimated. Although pure seawater is harmful to most living plants, diluted seawater could represent a promising integration to meet the crop’s nutrient and water requirements. In the current trial, we compared the effects of moderate and high concentrations of seawater and a comparable NaCl solution on a salt-tolerant (Tetragonia tetragonioides) and a salt-sensitive (Lactuca sativa) crop grown in hydroponics. We tested the hypothesis that, due to its mineral composition, diluted seawater would result in a less stressful growing medium than NaCl. We observed that diluted seawater resulted in a less detrimental growing medium compared to an EC-comparable NaCl solution, with remarkable differences between the salt-tolerant and the salt-sensitive species. While the growth rates in Tetragonia did not vary between the two types of stress, diluted seawater led to a higher FW and DW biomass yield in the salt-sensitive lettuce compared to the NaCl treatment. Moreover, NaCl reduced the water consumption and water productivity in Tetragonia. In lettuce, NaCl-treated plants demonstrated lower water use efficiency and water productivity compared to the EC-comparable seawater treatment. Physiological parameters and the concentration of mineral elements, phenolics and proline also demonstrated that, due to different mineral composition, seawater is a less stressful growing medium compared to a NaCl solution at comparable EC.
1. Introduction
Currently, the main agricultural challenge is to meet the rapidly growing and changing demand for food in an environmentally and socially sustainable way [1]. Whilst the increase in food production will have a decisive role in meeting this target, the Earth’s finite resources will negatively affect the process, with food producers already experiencing greater competition for natural resources [2]. Under this scenario, the production of food using alternative natural resources, including salt-affected soils and saline waters, might represent a necessary tool to explore [3].
Plants can be arranged in two main groups with respect to their response to salinity: salt-sensitive (i.e., glycophyte) and salt-tolerant (i.e., halophyte) species [4]. Other classifications, such as for instance Maas and Hoffman’s [5], correlate the crop yield to the electroconductivity in the soil identifying four groups of responses: sensitive, moderately sensitive, moderately tolerant and tolerant. Currently, the majority of agricultural crops are salt-sensitive. A possible explanation for this is that the domestication and selection of the requested productive traits have been made under nonsaline conditions, where a possible loss of salt-tolerance traits did not represent an actual constraint [6]. Despite these detailed classifications, most data on crop salt-tolerance, for instance those presented by the FAO, are actually based on a single variety of a specific crop species [5,7]. However, it has been shown that different varieties are characterized by different tolerance to salinity [8]. Moreover, data on crop salinity tolerance are often obtained through screening made with NaCl solutions, which do not reflect the more complex salinity conditions of salt-affected soils. In fact, only a small number of studies on salinity consist of enough critical information to meet the requirements for a good salt-tolerance dataset [9]. In response to the increasing saline conditions in several world areas, a number of halophyte species have been recently investigated as potential new crops for these harsh environments [3,10,11,12,13,14,15,16]. Anyhow, to the best of our knowledge, the role of different salt types on determining specific salinity stress conditions for the plant has rarely been investigated. A screening that would consider the more salt-tolerant varieties of commonly grown glycophyte crops along with potential new edible halophyte crops, under different types of salinity conditions, would provide a better understanding of potential crops suitable for saline agriculture.
Salt-affected soils occur in more than 100 countries, and their worldwide extent is estimated at about 1 billion ha [17] more than 34 Mha arable lands are salt affected [18]. These surfaces are currently not suitable for most agricultural crops either because they are coastal or because inappropriate irrigation practices have degraded the soil and depleted or salinized the groundwater [19]. The causes of soil salinization are numerous, including the use of saline irrigation water, the deposition of salts on the soil surface due to capillary rise from a shallow water table, drought, poor drainage, and seawater intrusion in coastal areas [20]. Although seawater is often detrimental to a plant’s growth, it has been proven that its specific mineral composition may represent a balanced ionic environment for plants [21]. In fact, despite a very high sodium and chloride content (i.e., about 75% NaCl and 10% of MgCl2), seawater is rich in nutritive elements needed by plants [12], also including the necessary trace elements and living or dead micro-organisms [22,23]. In addition, the detrimental effects of salinity on crop yield depend on co-occurring factors. For instance, a different response in crop yield to increasing soil salinity has been observed for sandy and clay soils, with the former being favorably tested in saline conditions [8,13]. Moreover, despite a very limited number of studies comparing seawater salt stress to other salinity stress [24], the tolerance to seawater seems to be remarkably higher compared to that for NaCl solutions at comparable electrical conductivity (EC) [22]. Despite such fundamental differences, most available scientific literature still considers seawater to have the same detrimental effects of a salt solution, thereby underestimating the potential above-mentioned benefits. In fact, nonconventional water sources (i.e., saline water) have been successfully used in agriculture, especially in the MENA region, showing a potential in rehabilitating the marginal and degraded lands for meeting the food fodder demands [25].
Since, as stated above, a screening considering the role of different salt types on determining specific salinity stress conditions for plants has rarely been conducted, the current study was carried out with the aim of evaluating the differences in salt tolerance in one halophyte and one glycophyte species grown with two different types of saline media (i.e., NaCl solution and diluted seawater). The New Zealand spinach (Tetragonia tetragonioides (Pallas) Kuntz.) was chosen among the edible halophytes because it is already consumed in several regions, although cultivated on a very small scale [26], and, thus, is able to become a new staple food for saline environments [11]. Lettuce (Lactuca sativa L.) was chosen because is one of the most cultivated horticultural crops, focusing in particular on the largely diffused variety ‘Canasta’. In the current trial, we tested the hypothesis that due to different mineral composition, seawater would actually result in a less harmful growing medium for both crops than NaCl.
2. Materials and Methods
2.1. Experimental Design, Plant Material and Growth Conditions
The trial was carried out in 2019 at the greenhouse facilities of the Department of Agriculture, Food, Environment and Forestry—University of Florence (Italy). A hydroponic system was set up, with 20 plastic containers (4 L volume) constantly aerated, under the same growing conditions as reported in [11]. The first trial took place from 26 March to 7 May testing Tetragonia tetragonioides (Pallas) Kuntz. at different salinity levels and conditions. Tetragonia seeds were obtained by the Tuttosemi company (www.tuttosemi.com (accessed on 23 June 2021)) and germinated in a dark chamber at 18.5 °C starting from 8 January. About two months later, young plantlets were transplanted into 5 cm mesh pots filled with expanded clay and transferred in polystyrene layer (one plant per container) used as support in the hydroponic floating system. Half-strength Hoagland solution [27] was used as a growing medium for an additional 10 days, and plants were then gradually acclimated to the salinity conditions tested in the experiment. Salt acclimation occurred through the gradual (i.e., twice a week for a two-week period) addition of seawater or NaCl in the respective treatments, until the achievement of the final concentrations. Five treatments were set up: C (control, half-strength Hoagland solution, EC = 1.5 dS m−1); M-SW (moderate seawater, 25% seawater and 75% half-strength Hoagland solution, EC = 14.5 dS m−1); M-SA (moderate salt, 150 mM NaCl half-strength Hoagland solution, EC = 13.5 dS m−1); H-SW (high seawater, 50% seawater and 50% half-strength Hoagland solution, EC = 24.5 dS m−1); H-SA (high salt, 280 mM NaCl half-strength Hoagland solution, EC = 23.5 dS m−1), for a total of four plants per treatment, randomly distributed.
The second trial was carried out from 20 May to 20 June with Lactuca sativa L. Plantlets obtained from a nursery were transplanted into 5 cm mesh pots filled with expanded clay. They were adapted to hydroponic conditions and then gradually acclimated to salinity conditions, up to the beginning of the trial. As for Tetragonia, salt acclimation occurred through the gradual addition of seawater or NaCl reaching in 10 days the final concentrations. Afterwards, plants were grown under the following salinity conditions: C (control, half-strength Hoagland solution, EC = 1.5 dS m−1); M-SW (moderate seawater, 5% seawater and 95% half-strength Hoagland solution, EC = 4.5 dS m−1); M-SA (moderate salt, 50 mM NaCl half-strength Hoagland solution, EC = 5.5 dS m−1); H-SW (high seawater, 15% seawater and 85% half-strength Hoagland solution, EC = 12.5 dS m−1); H-SA (high salt, 100 mM NaCl half-strength Hoagland solution, EC = 12.5 dS m−1), again for a total of four plants per treatment, randomly distributed. For both experiments, samples from the nutrient solution were collected twice a week to measure pH and EC by a laboratory pH meter (pHmeter PHM 210 Meter Lab, Radiometer Analytical, Lyon, France) and the nutrient solutions (treatments) were replaced every two weeks. Seawater used in the experiment was collected at Marina di Pisa (Italy) one week before the beginning of each experiment and stored in 20 L sterile tanks at 4 °C. The chemical and physical characteristics of seawater are reported in Table 1.

Table 1.
Seawater chemical and physical characteristics.
Throughout both trials, plants were maintained at relative humidity ranging from 40% to 55%, natural light, (light intensity reaching 700 µmol m−2 s−1 during sunny days) and 28 °C/18 °C day/night air temperature.
2.2. Plant Growth and Biomass Yield
The increment of plants’ growth for the two crops was determined by weighting all plants along with the pot on a weekly basis. After the final sampling, empty pots and expanded clay weight was detracted from the previous weights, thus obtaining the entire plant fresh weight. At harvest, samples of fresh leaves were collected from each replicate per treatment (n = 4), frozen in liquid nitrogen and stored at −80 °C for further analyses. Subsequently, plants were divided into leaves and roots and weighted separately, with the aim of assessing the yield production through the plants’ fresh weight.
2.3. Water Consumption and Water Use Efficiency
For both plants species, the crop evapotranspiration (ET) rate was measured on a two-week basis by recording the volume before replacing the nutritive solution. Since the two species are characterized by different cycle lengths (i.e., 43 days for Tetragonia and 32 days for lettuce), ET was reported both cumulatively for the entire crop cycle and as average daily consumption. Water use efficiency (WUE) was calculated as the ratio between the whole oven-dry biomass and total ET throughout the crop cycle, as follows:
where DWwhole plant = whole plant dry weight (g), ET = crop evapotranspiration (L).
WUE = DWwhole plant/ET
Water productivity (WP) was used to better correlate the biomass production and ET, as the fresh shoot is the edible part of both the considered species. This parameter was calculated as the ratio between the fresh marketable biomass and total ET throughout the crop cycle, as follows:
where FWleaves = fresh weight of the edible and marketable leaves (g), ET = crop evapotranspiration (L), according to [28].
WP = FWleaves/ET
2.4. Leaf Gas-Exchange Parameters
Leaf gas-exchange parameters were determined using the open gas-exchange system Li-6400 XT (Li-COR, Lincoln, NE, USA) once every two weeks throughout the crop cycle on four plants per treatment. Net photosynthetic rate and stomata conductance were measured on the youngest fully expanded leaves from the apex at ambient relative humidity, reference CO2 of 400 µmol mol−1, flow rate of 400 µmol s−1, chamber temperature of 25 °C and photosynthetically active radiation (PAR) of 700 µmol m−2 s−1.
2.5. Concentration of Mineral Elements, Phenolics and Proline in Edible Leaves
The sodium, potassium and calcium concentration in young fully expanded leaves (n = 4) was obtained after digesting 0.1 g of ground dried tissues in 10 mL of 0.5 M HNO3 by shaking vials in the dark at 25 °C for 48 h, as reported in [13]. Diluted extracts were analyzed using a Flame Photometer Digiflame2000 (Lab Services SAS, Rome, Italy). The values of the calibration curve ranged from 0 to 0.1 mg mL−1 for Na and K (R2 = 0.998) and from 0 to 0.05 mg mL−1 for Ca (R2 = 0.999) determination.
Total phenolic compounds concentration in young fully expanded leaves (n = 4) was calculated following the protocol reported in [29], through a colorimetric total phenolics assay that utilizes Folin–Ciocalteu reagent (FCR): fresh dried samples (0.1 g) were grounded and left incubating in the dark with ice-cold 95% (vol/vol) methanol for 48 h. After that, samples were left reacting with 10% (vol/vol) FCR and 700 mM Na2CO3 for 2 h, and then the absorbance was read at 765 nm. Total phenolics are calculated as gallic acid equivalents using the regression equation between gallic acid standards and A765. The values of the calibration curve ranged from 0 to 150 µL mL−1 (R2 = 0.999). The proline concentration in young fully expanded leaves (n = 4) was calculated according to [30]: fresh dried samples (0.2 g) were grounded and left reacting with ninhydrin in glacial acetic acid at 100 °C for 30 min followed by spectrophotometric measurement of the proline–ninhydrin condensation product in the reaction mixture itself. The values of the calibration curve ranged from 0 to 100 µg mL−1 (R2 = 0.995 for New Zealand spinach samples and R2 = 0.993 for lettuce samples).
2.6. Statistical Analysis
Both experiments followed a complete randomized design to take into account the different treatments conditions. All collected data were analyzed, within each experiment, through one-way ANOVA using GraphPad Prism 6 for Windows (GraphPad software Inc., San Diego, CA, USA). Post hoc comparisons (Tukey’s HSD) were made to contrast the levels of the independent variables, and differences were considered significant when p ≤ 0.05.
3. Results
3.1. Plant Growth and Biomass Yield
The temporal patterns of fresh weight for each species are shown in Figure 1. Tetragonia showed similar growth rates in moderate-salinity conditions, with an increase in plant final biomass of 27.4% and of 47.8% in M-SW and M-SA, respectively, compared to the control. However, high-salinity treatment determined a significant growth reduction compared to both moderate-salinity conditions and control. No significant differences were observed in growth rate in Tetragonia due to the type of salinity stress (i.e., M-SW vs. M-SA and H-SW vs. H-SA). On the other hand, the response of lettuce followed a decreasing trend in accordance with increasing salinity: in particular, all salinity treatments significantly decreased the plants’ growth compared to that of the control (i.e., 27% and 32% in M-SW and M-SA, respectively), with no significant differences found between the stress types under moderate stress conditions. However, under high salinity, a significantly stronger growth reduction was observed in H-SA (83%) compared to that in H-SW (62%) treatment.
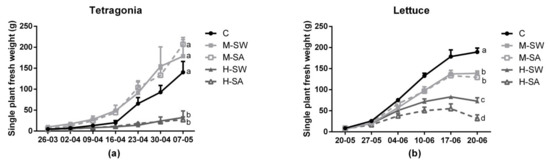
Figure 1.
The fresh weight of whole Tetragonia (a) and lettuce (b) plants. Values are single plants’ weight means (n = 4) ± SEM expressed in grams. Different letters at the final sampling indicate significant differences assessed at p < 0.05 (Tukey’s test).
The dry biomass weight of the plants at the end of the cycle is shown in Table 2.
Table 2.
The dry weight of whole plants at harvest.
In the case of Tetragonia, we found the same differences among treatments as observed for fresh weight. Regarding lettuce, unlike fresh weight, the dry biomass of plants grown under moderate salinity conditions was not significantly different from that of the control. However, under high-salinity treatments we observed the same response as in fresh weight: a significant reduction in dry weight determined by H-SA treatment compared to H-SW treatment was observed.
The commercial yield of the two species (i.e., shoot fresh weight) is reported in Figure 2. The shoot production of Tetragonia was enhanced, although not significantly, by both moderate-salinity treatments: an increase of 59.7% and of 55.2% was observed for M-SW and M-SA, respectively. The crop yield showed a significant and severe decrease under both high-salinity treatments. In lettuce, all treatments resulted in a significant reduction in commercial yield compared to that of control. Both medium-salinity stress conditions resulted in 32% (M-SW) and 38% (M-SA) yield reduction compared to control. Under higher-stress conditions a significant yield reduction due to NaCl treatment compared to the seawater was also observed, with H-SA treatment showing 86% and H-SW showing 68.5% yield reductions.
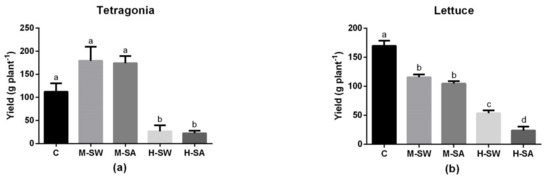
Figure 2.
The Tetragonia (a) and lettuce (b) plants’ yield. Values are single plants’ shoot weight means (n = 4) ± SEM expressed in grams. Different letters indicate significant differences assessed at p < 0.05 (Tukey’s test).
3.2. Water Consumption, Water Use Efficiency and Water Productivity
Plants’ cumulative and daily average ETs are reported in Table 3.
Table 3.
Cumulative and daily average ETs.
In Tetragonia, only H-SA-treated plants consumed significantly less water compared to the control; in lettuce, the same significant reduction was observed for both high-salinity treatments, whereas both moderate-stress treatments did not show any significant difference compared to the control.
The WUE and the WP of the tested species are reported in Figure 3.
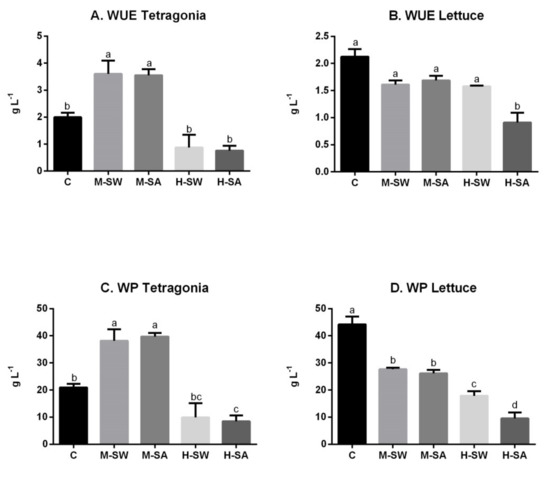
Figure 3.
Plants WUE (A,B) and WP (C,D). Values are means (n = 4) ± SEM expressed in grams per liter. Different letters indicate significant differences assessed at p < 0.05 (Tukey’s test).
Tetragonia and lettuce responded differently, with the halophyte significantly enhancing its WUE and WP in both M-SW and M-SA compared to the control. In fact, the WUE increased by 80.4% and of 77.8% and, similarly, an increase in WP of 82% and of 89.4% was recorded under M-SW and M-SA, respectively. By contrast, lettuce showed for both seawater treatments a WUE comparable to the control, whereas WP decreased in all salt-treated plants compared to the control. Moreover, whereas no significant differences were assessed in lettuce WUE and WP between the two moderate stresses (i.e., M-SW vs. M-SA), the H-SA treatment significantly reduced both WUE and WP compared to the EC-comparable H-SW.
3.3. Leaf Gas-Exchange Parameters
The net assimilation rates in Tetragonia were significantly affected by the treatments (Figure 4).
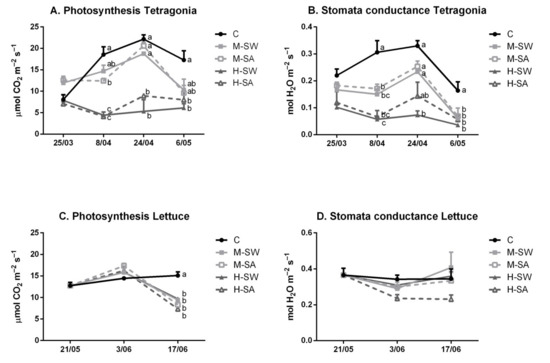
Figure 4.
Photosynthetic rate (A,C) and stomata conductance (B,D) of the tested species. Values are means (n = 4) ± SEM measured during the crop cycles. Different letters indicate significant differences at the same recording day assessed at p < 0.05 (Tukey’s test).
In particular, starting two weeks after the beginning of the trial, only plants submitted to M-SW treatment showed comparable photosynthetic rate to that of the control. M-SA was significantly lower than the control. Both high-concentration salt treatments showed significant lower net assimilation rates compared to both control and moderate-salinity stress. However, no significant differences were observed between the two types of salt stress at comparable EC (i.e., M-SW vs. M-SA; H-SW vs. H-SA). By the end of the trial, the photosynthetic rate in both moderate-stress treatments did not statistically differ from the control, while in high-stress treatments photosynthesis was statistically lower than control. Stronger response was observed for stomatal conductance for which both salinity treatments resulted in a significant decrease in comparison to the control. Lettuce was not affected by salinity during its growth peak, whereas at the end of the crop cycle all salinity treatments resulted in a lower photosynthetic rate compared to control conditions. Moreover, although a decreasing pattern was observed for the H-SA treatment compared to the control, lettuce stomata conductance was not significantly reduced by any treatment.
3.4. Element, Phenolic Compounds and Proline Concentration in Edible Leaves
Sodium (Na) concentration in edible leaves did significantly increase with increasing salinity in both species (Table 4), with both SA treatments leading to a higher accumulation of this element compared to the respective SW treatment. Salinity also led to a significant reduction in leaf potassium (K) concentration in both species, whereas calcium (Ca) accumulation followed different trends in the two species. In lettuce, this element significantly dropped in all treatments compared to the control whereas in Tetragonia the H-SW treatment showed the same Ca concentration as the control, whereas in H-SA its concentration was significantly lower than that of the control (Table 4).
Table 4.
The Na, K and Ca concentration in edible leaves for different treatments.
In Tetragonia, the concentration in phenolic compounds in edible leaves significantly decreased only for H-SA treatment, while it was comparable to the control in all the other salinity treatments (Figure 5). In lettuce, a significant increase compared to the control was observed in both high-salinity treatments ranging from 81% in H-SW to 130% H-SA.
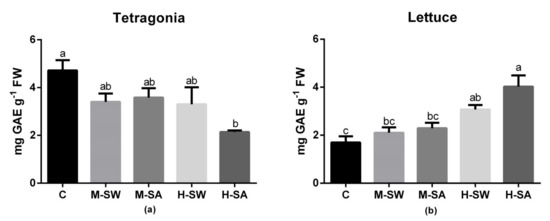
Figure 5.
Phenolic concentration in the edible leaves of Tetragonia (a) and lettuce (b). Values are means (n = 4) ± SEM measured at the end of the crop cycles. Different letters indicate significant differences at p < 0.05 (Tukey’s test).
As shown in Table 5, the proline concentration in the edible leaves increased significantly at all salinity levels in Tetragonia, whereas this occurred only for the higher NaCl treatment in lettuce.
Table 5.
Proline concentration in the edible leaves.
In Tetragonia, the proline concentration in leaves was significantly higher under either salinity treatment in comparison to the control, but no significant differences among moderate and high salinity or between seawater and NaCl treatments were found. In lettuce, H-SA treatment led to a higher (and only significant) increase in proline concentration, whereas M-SA led to an increase of about four times and H-SW of almost seven times.
4. Discussion
4.1. Salinity Type Affected Biomass Growth Only in Salt-Sensitive Species
The comparison between NaCl and seawater salinity stress did not lead to any significant difference in the growth of the salt tolerant Tetragonia. In fact, the two stresses at comparable EC did not lead to significant differences either in the FW or in the DW and the yield. In addition, for these growth parameters, both M-SW and M-SA led to a biomass production comparable to that of the control, whereas H-SW and H-SA to significantly lower values, thereby suggesting a salinity threshold among the two tested groups of EC: 14 and 24 dS m−1. This is in agreement with results obtained by Atzori et al. [11], assessing for Tetragonia a salinity tolerance threshold up to EC of 18 dS m−1. Lettuce, on the other hand, showed a different response to the stress. Firstly, both M-SW and M-SA led to significant reductions in both FW and yield compared to control conditions (even if the two types of salt stress tested led to comparable results), as expected with lettuce being a salt-sensitive crop [9]. Even more important for the goal of the current trial, in the case of high-salinity treatments, the two stress types at comparable EC resulted in a significantly different FW, DW and yield, with H-SW showing higher biomass than its NaCl corresponding H-SA. These findings would support the findings of early research in which seawater was proved a less stressful growing medium than other media characterized by the same salinity but lacking most macro- and microelements dissolved in seawater [22]. In addition, similar results have been observed in Lactuca sativa L. cv. Mother-red, where a significant higher salt resistance was found when grown with seawater (EC of the culture was 10.6 dS m−1) compared to NaCl (EC 12.6 dS m−1) [24], even if the reason is probably also connected to the difference in EC between the two treatments.
4.2. Water Relations Are Differently Affected by Seawater and NaCl in Both Species
The type of stress affected the crop water relations in different ways in the two species. In the salt-resistant halophyte, the only stress significantly decreasing water consumption was the H-SA, whereas H-SW and both moderate salinities did not result in any significant difference compared to the control. Such differences between H-SW and H-SA suggest a higher stress determined by NaCl compared to seawater, even if the biomass produced with the two treatments was comparable. In fact, the decrease of water use is possibly connected to the limited water uptake and translocation due to a salt-stress-induced reduced stomata conductance [31,32], as shown in Section 3. Lettuce’s water consumption, on the other hand, was the same for both stresses, with both M-SW- and M-SA-treated plants in line with control and both H-SW and H-SA plants significantly lower than control. The results on H-SW reduction of lettuce water use are in agreement with those obtained when lettuce was grown with a share of 15% seawater and EC 9.2 dS m−1 [28,33]. Here again, the observed response was likely due to the reduced water uptake and translocation caused by salinity. This is also corroborated by the fact that the response of lettuce in terms of fresh and dry weight at the end of the cycle was different, with the fresh weight of moderately stressed plants significantly reduced compared to the control, whereas the dry weight showed an opposite response. This would suggest that even moderate salinity induces in this species an alteration in water uptake and translocation mechanisms. Regarding the WUE, lettuce was the only crop showing a different response to different type of stress, with the WUE being significantly reduced only in H-SA-treated plants compared to the control. By contrast, no differences were observed among EC-comparable stresses in Tetragonia, with both M-SW and M-SA significantly increasing the WUE compared to the control and both H-SW and H-SA showing values comparable to the control. Such results support data available in the literature, indicating for salt-tolerant crops a relatively constant WUE at increasing levels of salinity, whereas for sensitive crops, they indicate a decrease of WUE [34]. The WP of lettuce decreased compared to that of the control already at moderate-salinity conditions, but the most reducing treatment was H-SA, decreasing significantly not only compared to the control but also to the EC-equivalent H-SW treatment. In Tetragonia, instead, again both moderate salinities significantly increased the water productivity compared to control conditions, whereas while the H-SW-treated plants’ WP was comparable to the control, the H-SA-treated plants’ WP was significantly reduced, confirming at high salinity the major severity of the NaCl stress compared to that of seawater.
4.3. Physiological Insights
The investigation of the net photosynthetic rate and of the stomata conductance did not highlight significant differences in the comparison of seawater and NaCl stresses in both species. The observed effects on the two species are in line with results found in literature on the two species treated with seawater. For instance, Tetragonia showed a comparable trend of photosynthesis and stomata conductance when treated with 15% and 30% seawater (corresponding to EC 9.8 and 18.0 dS m−1, respectively) [11]. On the other hand, lettuce treated with 5% and 15% seawater (EC 3.4 and 6.1 dS m−1, respectively) was found to be less damaged in its photosynthetic rate compared to the current experiment, although the results on stomata conductance were similar to ours [28]. Anyhow, the overall lack of differences among the stress types suggests that both the photosynthetic apparatus and the stomata response of the tested species in moderate- and high-salinity conditions were actively able to deal with a general salinity stress, and deeper investigation on this topic should be carried out in order to evaluate possible differences between the two types of stress on the functionality of photosystems.
4.4. Effects of the Type of Salt Stress on the Nutritional Characteristics
In the current experiment, Na, Ca and K accumulation varied between EC-comparable stress types. Sodium, as expected, did increase in all salt-treated plants compared to the control, with plants growing with NaCl accumulating higher amounts of Na compared to seawater at comparable EC. Moreover, the accumulation of sodium in M-SA was so much higher than in M-SW as to be comparable to H-SW. Sodium accumulation in leaves represents one of most common responses to salinity in halophytes, especially for sodium-includer species such as Tetragonia [26,35]. In these cases, plants accumulate sodium in their vacuoles to be used for osmotic adjustments [36]. The decrease in Na and K accumulation observed in lettuce is also in line with previous published studies [37]. Interestingly, in moderate-salinity conditions, the accumulation of K decreased in M-SA-treated plants only, whereas the effect of M-SW was comparable to the control. Similar results were obtained on different glycophytes too, i.e., on the common spinach (Spinacia oleracea) [38]. Furthermore, in Tetragonia, the reduction in K concentration was stronger in NaCl-treated plants compared to that for seawater, and the accumulation of Ca in H-SW was the only one comparable to the control condition, whereas other treatments led to significant reductions. Such differences in plants treated with EC-comparable salt types might play a role in osmotic adjustment if efficiently compartmentalized at the cell level [39]. Since sodium could have a negative effect on human health, whereas K and Ca are among the main mineral elements lacking in the diet of over two-thirds of the world’s population [40], seawater in the growing medium seems to provide better results than NaCl from a food quality standpoint. The concentration of phenolic compounds followed different patterns in salt-tolerant and salt-sensitive species, although no significant differences were observed between EC-comparable stresses. In Tetragonia leaves, the concentration of phenolic compounds was always comparable to the control except for H-SA treatment, where the concentration significantly decreased overtime. In a previous experiment, we found that the phenolics concentration did significantly decrease with seawater irrigation at EC of 9.8 and 18 dS m−1 [11]. This suggests that Tetragonia may produce other specific chemical compounds (i.e., proline) to act as compatible solutes for osmotic adjustment, as shown in a previous study [26]. The decrease in phenolic compounds in response to salinity has been observed in other halophyte plants. For instance, a severe decrease in polyphenol concentration was observed in Mesembryanthemum edule L. in response to increasing salinity [41]. However, it has been also shown that this response may vary also according to the species and within each species with the varieties. An interesting trial comparing two different Cakile maritima Scop. accessions showed that the high salt-stress tolerance of one accession was concomitant with leaf enrichment in phenolic compounds, whereas the more-sensitive accession showed a decrease in these compounds, thereby suggesting that a difference in ability to accumulate polyphenols contributes to the difference in salt-tolerance of the two varieties [42]. This would also suggest that actually Tetragonia should be considered less salt-tolerant than other halophytes. In lettuce, salinity stress led to an increase in phenolic concentration accordingly with increasing salinity: both moderate treatments resulted comparable to the control, whereas the high-salinity treatments assessed significantly higher concentration compared to control conditions. Similar results have been reported for several plants [43], including lettuce [44,45], since phenolics are among the secondary metabolites whose concentration is modulated by plants as a function of osmotic regulation [46]. However, a review on this topic pointed out that the direction of phenolic changes upon plant treatment with high doses of salt still provides inconclusive results and needs further investigation [47]. The proline concentration showed an increase in response to salt stress mostly in the halophyte species compared to that in the glycophyte. Other studies reported the same tendency. For instance, a trial comparing the physiological response of Plantago major L. (glycophyte) and Thellungiella halophila Mey. (halophyte) to salt stress reported a significant increase in this amino acid concentration in the halophyte compared to that of the glycophyte [48]. The increase in proline concentration under saline conditions in salt-tolerant species is associated with the improvement of enzyme and membrane integrity and is involved in osmotic adjustments in response to the stress [49]. It is also likely that high constitutive and stress-induced levels of proline in salt-tolerant plants reduce ROS generation, thereby being involved in detoxification of the superoxide radical [50].
5. Conclusions
The hypothesis that, due to its different mineral composition, seawater would actually result in a less stressful growing medium compared to a NaCl solution at comparable EC was supported by our results. However, remarkable differences between the salt-tolerant and the salt-sensitive investigated species were observed. The most promising results were those related to growth: while for Tetragonia no differences were observed between the two stresses at comparable EC, in the salt-sensitive lettuce H-SW treatment led to a higher biomass compared to its NaCl corresponding H-SA for FW, DW and yield. Additionally, water relations highlighted differences between the tested stress, with Tetragonia decreasing water consumption in H-SA-treated plants compared to H-SW-treated plants, instead in line with control conditions. Such differences between H-SW and H-SA collaborate to indicate a major stress administered through the NaCl solution compared to seawater, even if the biomass produced with the two treatments was in the end comparable. Moreover, in lettuce, the WUE was significantly reduced in plants treated with H-SA only compared to the control, with H-SA also being the treatment associated with the lower WP, decreasing significantly not only compared to the control but also to the EC-equivalent H-SW treatment. Similarly, in Tetragonia, the H-SW-treated plants’ WP was comparable to that of the control, whilst the H-SA-treated plants’ WP was significantly reduced, confirming at high salinity the major severity of the NaCl stress compared to seawater. The use of seawater on lettuce increased the concentration of phenolic compounds, which are associated with an increase of biofortification effect in the edible parts. The results presented here confirm the need of a more complete salt-tolerance dataset for cultivated horticultural crops, since data on salinity thresholds are often obtained through screenings that do not reflect the complex salinity conditions of salt-affected soils: the highlighted differences in the salt response to salt stresses types in fact suggest a need for further studies investigating the specific salt conditions actually present in salt-affected soils.
Author Contributions
Conceptualization, W.G.N. and G.A.; data curation, W.G.N. and G.A.; formal analysis, W.G.N. and G.A.; funding acquisition, E.M., C.P. and S.M.; methodology, G.A.; project administration, E.M., C.P. and S.M.; resources, E.M.; supervision, E.M., C.P. and S.M.; visualization, W.G.N. and G.A.; writing—original draft, G.A.; writing—review and editing, W.G.N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the POR FSE 2014–2020 Program of the Regione Toscana with the Jellyfish Barge: una serra galleggiante autosufficiente per coltivare il mare Project (UNIFI_FSE2017) and supported by Fondazione CR Firenze (project MEDUSA—Jellyfish Barge) and by PNAT S.R.L.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors gratefully acknowledge the LINV and PNAT S.R.L. for inspiring and supporting this research.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Béné, C.; Oosterveer, P.; Lamotte, L.; Brouwer, I.D.; de Haan, S.; Prager, S.D.; Talsma, E.F.; Khoury, C.K. When food systems meet sustainability—Current narratives and implications for actions. World Dev. 2019, 113, 116–130. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of Feeding 9 Billion People. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Atzori, G. The potential of edible halophytes as new crops in saline agriculture—The ice plant (Mesembryanthemum crtstallinum L.) case study. In Future of Sustainable Agriculture in Saline Environments; Negacz, K., Vellinga, P., Barrett-Lennard, E., Choukr-Allah, R., Elzenga, T., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 443–460. ISBN 9781003112327. [Google Scholar]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Maas, E.V.; Hoffman, G.J. Crop Salt Tolerance—Current Assessment. J. Irrig. Drain. Div. Am. Soc. Civ. Eng. 1977, 103, 115–134. [Google Scholar] [CrossRef]
- Cheeseman, J.M. The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol. 2015, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Tanji, K.K.; Kielen, N.C. Agricultural Drainage Water Management in Arid and Semi-Arid Areas; FAO Irrigation and Drainage Paper 61; FAO: Rome, Italy, 2002. [Google Scholar]
- De Vos, A.; Bruning, B.; van Straten, G.; Oosterbaan, R.; Rozema, J.; van Bodegom, P. Crop Salt Tolerance under Controlled Field Conditions in The Netherlands, Based on Trials Conducted at Salt Farm Texel; Salt Farm Texel: Den Burg, The Netherlands, 2016. [Google Scholar]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hortic. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Y.; Han, G.; Guo, J.; Meng, Z.; Chen, M. Progress in the Study and Use of Seawater Vegetables. J. Agric. Food Chem. 2020, 68, 5998–6006. [Google Scholar] [CrossRef] [PubMed]
- Atzori, G.; Nissim, W.G.; Macchiavelli, T.; Vita, F.; Azzarello, E.; Pandolfi, C.; Masi, E.; Mancuso, S. Tetragonia tetragonioides (Pallas) Kuntz. as promising salt-tolerant crop in a saline agriculture context. Agric. Water Manag. 2020, 240, 106261. [Google Scholar] [CrossRef]
- Atzori, G.; Mancuso, S.; Masi, E. Seawater potential use in soilless culture: A review. Sci. Hortic. 2019, 249, 199–207. [Google Scholar] [CrossRef]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of increased seawater salinity irrigation on growth and quality of the edible halophyte Mesembryanthemum crystallinum L. under field conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- De Vos, A.C.; Broekman, R.; de Almeida Guerra, C.C.; van Rijsselberghe, M.; Rozema, J. Developing and testing new halophyte crops: A case study of salt tolerance of two species of the Brassicacea, Diplotaxis tenuifoli and Cochlearia officinalis. Environ. Exp. Bot. 2013, 92, 154–164. [Google Scholar] [CrossRef]
- De Vos, A.C.; Broekman, R.; Groot, M.P.; Rozema, J. Ecophysiological response of Crambe maritima to airborne and soil-borne salinity. Ann. Bot. 2010, 105, 925–937. [Google Scholar] [CrossRef] [PubMed]
- Rozema, J.; Cornelisse, D.; Zhang, Y.; Li, H.; Bruning, B.; Katschnig, D.; Broekman, R.; Ji, B.; van Bodegom, P. Comparing salt tolerance of beet cultivars and their halophytic ancestor: Consequences of domestication and breeding programmes. AoB Plants 2015, 7, plu083. [Google Scholar] [CrossRef] [PubMed]
- FAO. Status of the World’s Soil Resources Main Report; FAO: Rome, Italy, 2015. [Google Scholar]
- FAO. The State of the World’s Land and Water Resources for Food and Agriculture (SOLAW). Managing Systems at Risk; The Food and Agriculture Organization of the United Nations: Rome, Italy; Earthscan: London, UK, 2011. [Google Scholar]
- Cheeseman, J.M. Food Security in the Face of Salinity, Drought, Climate Change, and Population Growth. In Halophytes for Food Security in Dry Lands; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 111–124. ISBN 9780128018545. [Google Scholar]
- Singh, A. Soil salinization and waterlogging: A threat to environment and agricultural sustainability. Ecol. Indic. 2015, 57, 128–130. [Google Scholar] [CrossRef]
- Boyko, H. (Ed.) Salinity and Aridity New Approaches to Old Problems; Dr. W. Junk Publishers: The Hague, The Netherlands, 1966. [Google Scholar]
- Boyko, H.; Boyko, E. Experiments of Plant Growing under Irrigation with Saline Waters from 2000 mg/L T.D.S. (Total Diluted Solids) up to Sea-water of Oceanic Concentration, without Desalination. In Salinity and Aridity New Approaches to Old Problems; Boyko, H., Ed.; Dr. W. Junk Publishers: The Hague, The Netherlands, 1966; pp. 214–282. [Google Scholar]
- Eyster, C. Seawater as a Source of Plant Nutrients. Nature 1968, 220, 260–261. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Kogi, M.; Yanagisawa, T. Effects of Salinity and Nutrients in Seawater on Hydroponic Culture of Red Leaf Lettuce. Environ. Control Biol. 2014, 52, 189–195. [Google Scholar] [CrossRef][Green Version]
- Hussain, M.I.; Muscolo, A.; Farooq, M.; Ahmad, W. Sustainable use and management of non-conventional water resources for rehabilitation of marginal lands in arid and semiarid environments. Agric. Water Manag. 2019, 221, 462–476. [Google Scholar] [CrossRef]
- Yousif, B.S.; Liu, L.Y.; Nguyen, N.T.; Masaoka, Y.; Saneoka, H. Comparative Studies in Salinity Tolerance between New Zealand Spinach (Tetragonia tetragonioides) and chard (Beta vulgaris) to salt stress. Agric. J. 2010, 5, 19–24. [Google Scholar] [CrossRef][Green Version]
- Hoagland, D.R.; Arnon, D.I. Nutrient Solutions for Hydroponic Culture; University of California Agricultural Experimental Station Circular: Berkeley, CA, USA, 1938; Volume 347. [Google Scholar]
- Atzori, G.; Guidi Nissim, W.; Caparrotta, S.; Masi, E.; Azzarello, E.; Pandolfi, C.; Vignolini, P.; Gonnelli, C.; Mancuso, S. Potential and constraints of different seawater and freshwater blends as growing media for three vegetable crops. Agric. Water Manag. 2016, 176, 255–262. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Shabnam, N.; Tripathi, I.; Sharmila, P.; Pardha-Saradhi, P. A rapid, ideal, and eco-friendlier protocol for quantifying proline. Protoplasma 2016, 253, 1577–1582. [Google Scholar] [CrossRef] [PubMed]
- Downton, W.J.S.; Grant, W.J.R.; Robinson, S.P. Photosynthetic and Stomatal Responses of Spinach Leaves to Salt Stress. Plant Physiol. 1985, 77, 85–88. [Google Scholar] [CrossRef]
- Maggio, A.; Raimondi, G.; Martino, A.; De Pascale, S. Salt stress response in tomato beyond the salinity tolerance threshold. Environ. Exp. Bot. 2007, 59, 276–282. [Google Scholar] [CrossRef]
- Atzori, G.; Guidi Nissim, W.; Caparrotta, S.; Santantoni, F.; Masi, E. Seawater and water footprint in different cropping systems: A chicory (Cichorium intybus L.) case study. Agric. Water Manag. 2019, 211, 172–177. [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Salinity effect on crop development and yield, analysis of salt tolerance according to several classification methods. Agric. Water Manag. 2003, 62, 37–66. [Google Scholar] [CrossRef]
- Neves, M.A.; Miguel, M.G.; Marques, C.; Panagopoulos, T.; Beltrao, J. The Combined Effects of Salts and Calcium on Growth and Mineral Accumulation of Tetragonia tetragonioides—A Salt Removing Species. WSEAS Trans. Environ. Dev. 2008, 4, 1–5. [Google Scholar]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Bartha, C.; Fodorpataki, L.; Martinez-Ballesta, M.d.C.; Popescu, O.; Carvajal, M. Sodium accumulation contributes to salt stress tolerance in lettuce cultivars. J. Appl. Bot. Food Qual. 2015, 88, 42–48. [Google Scholar]
- Caparrotta, S.; Masi, E.; Atzori, G.; Diamanti, I.; Azzarello, E.; Mancuso, S.; Pandolfi, C. Growing spinach (Spinacia oleracea) with different seawater concentrations: Effects on fresh, boiled and steamed leaves. Sci. Hortic. 2019, 256, 1–7. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Falleh, H.; Jalleli, I.; Ksouri, R.; Boulaaba, M.; Guyot, S.; Magné, C.; Abdelly, C. Effect of salt treatment on phenolic compounds and antioxidant activity of two Mesembryanthemum edule provenances. Plant Physiol. Biochem. 2012, 52, 1–8. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Debez, A.; Falleh, H.; Grignon, C.; Abdelly, C. Salinity effects on polyphenol content and antioxidant activities in leaves of the halophyte Cakile maritima. Plant Physiol. Biochem. 2007, 45, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Huang, J.; Gruber, M.Y.; Kaddour, R.; Lachaâl, M.; Ouerghi, Z.; Hannoufa, A. The impact of genotype and salinity on physiological function, secondary metabolite accumulation, and antioxidative responses in lettuce. J. Agric. Food Chem. 2010, 58, 5122–5130. [Google Scholar] [CrossRef]
- Khani, A.; Barzegar, T.; Nikbakht, J.; Ghahremani, Z. Effect of foliar spray of calcium lactate on the growth, yield and biochemical attribute of lettuce (Lactuca sativa L.) under water deficit stress. Adv. Hortic. Sci. 2020, 34, 11–24. [Google Scholar]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Waskiewicz, A.; Muzolf-Panek, M.; Golinski, P. Phenolic content changes in plants under salt stress. In Ecophysiology and Responses of Plants under Salt Stress; Springer: Berlin/Heidelberg, Germany, 2013; pp. 283–314. ISBN 9781461447474. [Google Scholar]
- Smirnoff, N.; Cumbes, J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemisrty 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Radyukina, N.L.; Kartashov, A.V.; Ivanov, Y.V.; Shevyakova, N.I.; Kuznetsov, V.V. Functioning of defense systems in halophytes and glycophytes under progressing salinity. Russ. J. Plant Physiol. 2007, 54, 806–815. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).