
Immediate Insulin Treatment Prevents Diabetes-Induced Gut Region-Specific Increase in the Number of Myenteric Serotonergic Neurons


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Tissue Handling
2.3. Fluorescent Immunohistochemistry
2.4. Statistical Analysis
3. Results
3.1. Disease Characteristics in Diabetic and Insulin-Treated Diabetic Rats
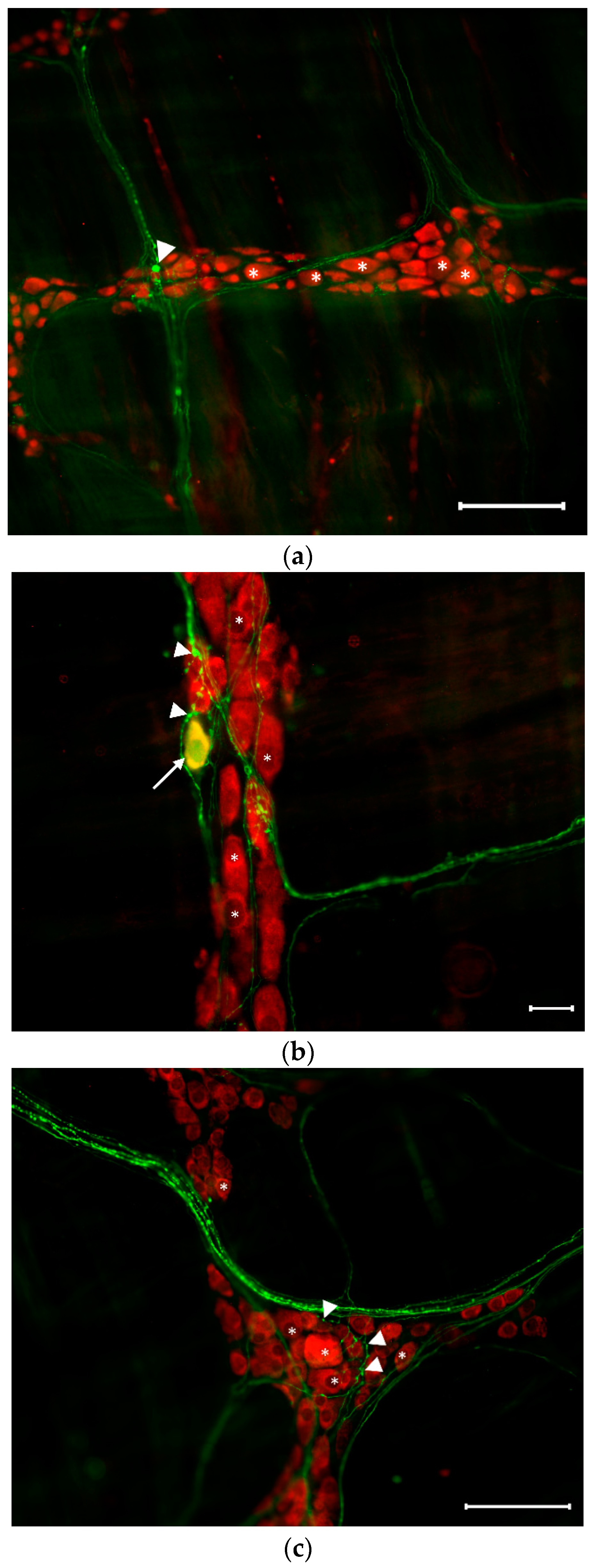
3.2. Distribution of Serotonergic Neurons in the Myenteric Ganglia of Controls
3.3. Proportion of Serotonergic Neurons in the Myenteric Ganglia of Diabetics and Insulin-Treated Diabetics
3.4. Proportion of Myenteric Ganglia Containing 5-HT-IR Neurons and the Amount of the Serotonergic Neurons in These Ganglia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azpiroz, F.; Malagelada, C. Diabetic neuropathy in the gut: Pathogenesis and diagnosis. Diabetologia 2016, 59, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B. The enteric nervous system: Normal functions and enteric neuropathies. Neurogastroenterol. Motil. 2008, 20, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Andres, H.; Rock, R.; Bridges, R.J.; Rummel, W.; Schreiner, J. Submucosal plexus and electrolyte transport across rat colonic mucosa. J. Physiol. 1985, 364, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Timmermans, J.P.; Hens, J.; Adriaensen, D. Outer submucous plexus: An intrinsic nerve network involved in both secretory and motility processes in the intestine of large mammals and humans. Anat. Rec. Off. Publ. Am. Assoc. Anat. 2001, 262, 71–78. [Google Scholar] [CrossRef]
- Krueger, D.; Michel, K.; Zeller, F.; Demir, I.E.; Ceyhan, G.O.; Slotta-Huspenina, J.; Schemann, M. Neural influences on human intestinal epithelium in vitro. J. Physiol. 2016, 594, 357–372. [Google Scholar] [CrossRef] [PubMed]
- Aouacheri, O.; Saka, S.; Krim, M.; Messaadia, A.; Maidi, I. The investigation of the oxidative stress-related parameters in type 2 diabetes mellitus. Can. J. Diabetes 2015, 39, 44–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrasekharan, B.; Srinivasan, S. Diabetes and the enteric nervous system. Neurogastroenterol. Motil. 2007, 19, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Jancso, Z.; Bodi, N.; Borsos, B.; Fekete, E.; Hermesz, E. Gut region-specific accumulation of reactive oxygen species leads to regionally distinct activation of antioxidant and apoptotic marker molecules in rats with STZ-induced diabetes. Int. J. Biochem. Cell Biol. 2015, 62, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Bodi, N.; Talapka, P.; Poles, M.Z.; Hermesz, E.; Jancso, Z.; Katarova, Z.; Izbeki, F.; Wittmann, T.; Fekete, E.; Bagyanszki, M. Gut region-specific diabetic damage to the capillary endothelium adjacent to the myenteric plexus. Microcirculation 2012, 19, 316–326. [Google Scholar] [CrossRef]
- Meldgaard, T.; Olesen, S.S.; Farmer, A.D.; Krogh, K.; Wendel, A.A.; Brock, B.; Drewes, A.M.; Brock, C. Diabetic Enteropathy: From Molecule to Mechanism-Based Treatment. J. Diabetes Res. 2018, 2018, 3827301. [Google Scholar] [CrossRef] [PubMed]
- Gotfried, J.; Priest, S.; Schey, R. Diabetes and the Small Intestine. Curr. Treat. Options Gastroenterol. 2017, 15, 490–507. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Young, R.L.; Leong, L.; Rogers, G.B.; Spencer, N.J.; Jessup, C.F.; Keating, D.J. The Diverse Metabolic Roles of Peripheral Serotonin. Endocrinology 2017, 158, 1049–1063. [Google Scholar] [CrossRef] [PubMed]
- Young, R.L.; Lumsden, A.L.; Martin, A.M.; Schober, G.; Pezos, N.; Thazhath, S.S.; Isaacs, N.J.; Cvijanovic, N.; Sun, E.W.L.; Wu, T.; et al. Augmented capacity for peripheral serotonin release in human obesity. Int. J. Obes. 2018, 42, 1880–1889. [Google Scholar] [CrossRef] [PubMed]
- Qu, Z.D.; Thacker, M.; Castelucci, P.; Bagyanszki, M.; Epstein, M.L.; Furness, J.B. Immunohistochemical analysis of neuron types in the mouse small intestine. Cell Tissue Res. 2008, 334, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Hempfling, C.; Neuhuber, W.L.; Worl, J. Serotonin-immunoreactive neurons and mast cells in the mouse esophagus suggest involvement of serotonin in both motility control and neuroimmune interactions. Neurogastroenterol. Motil. 2012, 24, e67–e78. [Google Scholar] [CrossRef]
- Costa, M.; Brookes, S.J.; Steele, P.A.; Gibbins, I.; Burcher, E.; Kandiah, C.J. Neurochemical classification of myenteric neurons in the guinea-pig ileum. Neuroscience 1996, 75, 949–967. [Google Scholar] [CrossRef]
- Spencer, N.J.; Sia, T.C.; Brookes, S.J.; Costa, M.; Keating, D.J. CrossTalk opposing view: 5-HT is not necessary for peristalsis. J. Physiol. 2015, 593, 3229–3231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, T.K.; Gershon, M.D. CrossTalk proposal: 5-HT is necessary for peristalsis. J. Physiol. 2015, 593, 3225–3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mawe, G.M.; Hoffman, J.M. Serotonin signalling in the gut--functions; dysfunctions and therapeutic targets. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 473–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, T.; Barton, M.J.; Hennig, G.W.; Birch, G.C.; Grainger, N.; Corrigan, R.D.; Koh, S.D.; Sanders, K.M.; Smith, T.K. Extensive projections of myenteric serotonergic neurons suggest they comprise the central processing unit in the colon. Neurogastroenterol. Motil. 2014, 26, 556–570. [Google Scholar] [CrossRef] [PubMed]
- Furness, J.B.; Costa, M. Neurons with 5-hydroxytryptamine-like immunoreactivity in the enteric nervous system: Their projections in the guinea-pig small intestine. Neuroscience 1982, 7, 341–349. [Google Scholar] [CrossRef]
- Banerjee, S.; Akbar, N.; Moorhead, J.; Rennie, J.A.; Leather, A.J.; Cooper, D.; Papagrigoriadis, S. Increased presence of serotonin-producing cells in colons with diverticular disease may indicate involvement in the pathophysiology of the condition. Int. J. Colorectal Dis. 2007, 22, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Faure, C.; Patey, N.; Gauthier, C.; Brooks, E.M.; Mawe, G.M. Serotonin signaling is altered in irritable bowel syndrome with diarrhea but not in functional dyspepsia in pediatric age patients. Gastroenterology 2010, 139, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Coyle, D.; Murph, J.M.; Doyle, B.; O’Donnell, A.M.; Gillick, J.; Puri, P. Altered tryptophan hydroxylase 2 expression in enteric serotonergic nerves in Hirschsprung’s-associated enterocolitis. World J. Gastroenterol. 2016, 22, 4662–4672. [Google Scholar] [CrossRef]
- Israelyan, N.; Margolis, K.G. Serotonin as a link between the gut-brain-microbiome axis in autism spectrum disorders. Pharmacol. Res. 2018, 132, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Del Colle, A.; Israelyan, N.; Gross Margolis, K. Novel aspects of enteric serotonergic signaling in health and brain-gut disease. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G130–G143. [Google Scholar] [CrossRef] [Green Version]
- Camilleri, M. Novel Diet; Drugs; and Gastric Interventions for Gastroparesis. Clin. Gastroenterol. Hepatol. 2016, 14, 1072–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lincoln, J.; Bokor, J.T.; Crowe, R.; Griffith, S.G.; Haven, A.J.; Burnstock, G. Myenteric plexus in streptozotocin-treated rats. Neurochemical and histochemical evidence for diabetic neuropathy in the gut. Gastroenterology 1984, 86, 654–661. [Google Scholar] [CrossRef]
- Gorio, A.; Di Giulio, A.M.; Donadoni, L.; Tenconi, B.; Germani, E.; Bertelli, A.; Mantegazza, P. Early neurochemical changes in the autonomic neuropathy of the gut in experimental diabetes. Int. J. Clin. Pharmacol. Res. 1992, 12, 217–224. [Google Scholar]
- Bodi, N.; Szalai, Z.; Chandrakumar, L.; Bagyanszki, M. Region-dependent effects of diabetes and insulin-replacement on neuronal nitric oxide synthase- and heme oxygenase-immunoreactive submucous neurons. World J. Gastroenterol. 2017, 23, 7359–7368. [Google Scholar] [CrossRef]
- Chandrakumar, L.; Bagyanszki, M.; Szalai, Z.; Mezei, D.; Bodi, N. Diabetes-Related Induction of the Heme Oxygenase System and Enhanced Colocalization of Heme Oxygenase 1 and 2 with Neuronal Nitric Oxide Synthase in Myenteric Neurons of Different Intestinal Segments. Oxidative Med. Cell. Longev. 2017, 2017, 1890512. [Google Scholar] [CrossRef]
- Bagyanszki, M.; Bodi, N. Diabetes-related alterations in the enteric nervous system and its microenvironment. World J. Diabetes 2012, 3, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Izbeki, F.; Wittman, T.; Rosztoczy, A.; Linke, N.; Bodi, N.; Fekete, E.; Bagyanszki, M. Immediate insulin treatment prevents gut motility alterations and loss of nitrergic neurons in the ileum and colon of rats with streptozotocin-induced diabetes. Diabetes Res. Clin. Pract. 2008, 80, 192–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Mello, S.T.; de Miranda Neto, M.H.; Zanoni, J.N.; Furlan, M.M. Effects of insulin treatment on HuC/HuD; NADH diaphorase; and nNOS-positive myoenteric neurons of the duodenum of adult rats with acute diabetes. Dig. Dis. Sci. 2009, 54, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Watkins, C.C.; Sawa, A.; Jaffrey, S.; Blackshaw, S.; Barrow, R.K.; Snyder, S.H.; Ferris, C.D. Insulin restores neuronal nitric oxide synthase expression and function that is lost in diabetic gastropathy. J. Clin. Investig. 2000, 106, 803. [Google Scholar] [CrossRef] [Green Version]
- King, A.J. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Yarandi, S.S.; Srinivasan, S. Diabetic gastrointestinal motility disorders and the role of enteric nervous system: Current status and future directions. Neurogastroenterol. Motil. 2014, 26, 611–624. [Google Scholar] [CrossRef]
- Ordog, T.; Hayashi, Y.; Gibbons, S.J. Cellular pathogenesis of diabetic gastroenteropathy. Minerva Gastroenterol. Dietol. 2009, 55, 315–343. [Google Scholar]
- Fujimiya, M.; Okumiya, K.; Yamane, T.; Maeda, T. Distribution of serotonin-immunoreactive nerve cells and fibers in the rat gastrointestinal tract. Histochem. Cell Biol. 1997, 107, 105–114. [Google Scholar] [CrossRef]
- Crowell, M.D.; Mathis, C.; Schettler, V.A.; Yunus, T.; Lacy, B.E. The effects of tegaserod; a 5-HT receptor agonist; on gastric emptying in a murine model of diabetes mellitus. Neurogastroenterol. Motil. 2005, 17, 738–743. [Google Scholar] [CrossRef]
- Farmer, A.D.; Bruckner-Holt, C.; Schwartz, S.; Sadler, E.; Kadirkamanthan, S. Diabetic Gastroparesis: Perspectives from a Patient and Health Care Providers. J. Patient Cent. Res. Rev. 2019, 6, 148–157. [Google Scholar] [CrossRef]
- Belai, A.; Lincoln, J.; Milner, P.; Burnstock, G. Progressive changes in adrenergic; serotonergic; and peptidergic nerves in proximal colon of streptozotocin-diabetic rats. Gastroenterology 1988, 95, 1234–1241. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Li, Z.; Chalazonitis, A.; Huang, Y.Y.; Mann, J.J.; Margolis, K.G.; Yang, Q.M.; Kim, D.O.; Cote, F.; Mallet, J.; Gershon, M.D. Essential roles of enteric neuronal serotonin in gastrointestinal motility and the development/survival of enteric dopaminergic neurons. J. Neurosci. 2011, 31, 8998–9009. [Google Scholar] [CrossRef]
- Liu, M.T.; Kuan, Y.H.; Wang, J.; Hen, R.; Gershon, M.D. 5-HT4 receptor-mediated neuroprotection and neurogenesis in the enteric nervous system of adult mice. J. Neurosci. 2009, 29, 9683–9699. [Google Scholar] [CrossRef]
- Gross, E.R.; Gershon, M.D.; Margolis, K.G.; Gertsberg, Z.V.; Li, Z.; Cowles, R.A. Neuronal serotonin regulates growth of the intestinal mucosa in mice. Gastroenterology 2012, 143, 408–417.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonscher, R.; Belkind-Gerson, J. Concise Review: Cellular and Molecular Mechanisms of Postnatal Injury-Induced Enteric Neurogenesis. Stem Cells 2019, 37, 1136–1143. [Google Scholar] [CrossRef]
- Grundmann, D.; Loris, E.; Maas-Omlor, S.; Schafer, K.H. Enteric Neurogenesis During Life Span Under Physiological and Pathophysiological Conditions. Anat. Rec. 2019, 302, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Manneras Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Backhed, F. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [Green Version]
- Wirth, R.; Bodi, N.; Maroti, G.; Bagyanszki, M.; Talapka, P.; Fekete, E.; Bagi, Z.; Kovacs, K.L. Regionally distinct alterations in the composition of the gut microbiota in rats with streptozotocin-induced diabetes. PLoS ONE 2014, 9, e110440. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) ± SEM | Blood Glucose Concentration (mmol/L) ± SEM | |||
|---|---|---|---|---|
| Initial | Final | Initial | Final (Average) | |
| Controls (n = 10) | 226.2 ± 4.3 | 457.1 ± 19.7 * | 5.46 ± 0.4 | 5.56 ± 0.1 |
| Diabetics (n = 7) | 226.7 ± 6.0 | 344.4 ± 16.0 *°° | 4.99 ± 0.4 | 25.54 ± 1.1 *°° |
| Insulin-treated diabetics (n = 8) | 241.3 ± 1.9 | 398 ± 8.9 *°+ | 4.85 ± 0.1 | 13.75 ± 1.0 *°°++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezei, D.; Bódi, N.; Szalai, Z.; Márton, Z.; Balázs, J.; Bagyánszki, M. Immediate Insulin Treatment Prevents Diabetes-Induced Gut Region-Specific Increase in the Number of Myenteric Serotonergic Neurons. Appl. Sci. 2021, 11, 5949. https://doi.org/10.3390/app11135949
Mezei D, Bódi N, Szalai Z, Márton Z, Balázs J, Bagyánszki M. Immediate Insulin Treatment Prevents Diabetes-Induced Gut Region-Specific Increase in the Number of Myenteric Serotonergic Neurons. Applied Sciences. 2021; 11(13):5949. https://doi.org/10.3390/app11135949
Chicago/Turabian StyleMezei, Diána, Nikolett Bódi, Zita Szalai, Zsuzsanna Márton, János Balázs, and Mária Bagyánszki. 2021. "Immediate Insulin Treatment Prevents Diabetes-Induced Gut Region-Specific Increase in the Number of Myenteric Serotonergic Neurons" Applied Sciences 11, no. 13: 5949. https://doi.org/10.3390/app11135949
APA StyleMezei, D., Bódi, N., Szalai, Z., Márton, Z., Balázs, J., & Bagyánszki, M. (2021). Immediate Insulin Treatment Prevents Diabetes-Induced Gut Region-Specific Increase in the Number of Myenteric Serotonergic Neurons. Applied Sciences, 11(13), 5949. https://doi.org/10.3390/app11135949