
Health Promoting Effect of Phyllanthus emblica and Azadiractha indica against Advanced Glycation End Products Formation


 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Preparation of Extracts
2.3. Tests for Flavonoids (Alkaline Reagent Test) and Phenolics (FeCl3 Test) Confirmation
2.4. Total Phenol Content
2.5. Total Flavonoid Content
2.6. In Vitro Antioxidant Activity- Reducing Power
2.7. Evaluation of Antioxidant Activity of Extracts by Hydrogen Peroxide (H2O2) Radical Scavenging Activity
2.8. Estimation of DPPH Mediated Free Radical Scavenging Activity
2.9. Investigation of Anti-Inflammatory In Vitro Activity by Inhibition of Protein Denaturation
2.10. Anti-Proteinase Action
2.11. Inhibition of Egg Albumin Denaturation: To Evaluate Anti-Arthritic Activity by In Vitro Assay
2.12. Membrane Stabilization Ability
2.12.1. Red Blood Cell (RBC) Suspension Preparation
2.12.2. Hemolysis by Heat Induction
2.12.3. Hypotonicity Induced Hemolysis
2.13. Investigation of Antiglycating and AGEs Inhibiting Property through Screening of Glycation Biomarkers
2.13.1. Incubation of P. emblica and A. indica Extracts with In Vitro Glycation System
2.13.2. Measurement of Browning in Glycated Samples
2.13.3. Effect of P. emblica and A. indica Extract on Protein Aggregation Index
2.13.4. Determination of Fibrillar State by Congo Red Assay
2.14. Biophysical Studies: AGEs Formation Inhibiting Properties of Extracts
2.14.1. UV Absorption
2.14.2. AGEs Specific Fluorescence Study
2.14.3. Fibrillar Inhibition Study by Thioflavin T Specific Fluorescence Study
2.14.4. Statistical Analysis
3. Results
3.1. The Initial Screening of Phenolic and Flavonoid Contents
3.2. Ferric Reducing Power as In Vitro Antioxidant Assay
3.3. Hydrogen Peroxide (H2O2) Radical Scavenging Ability
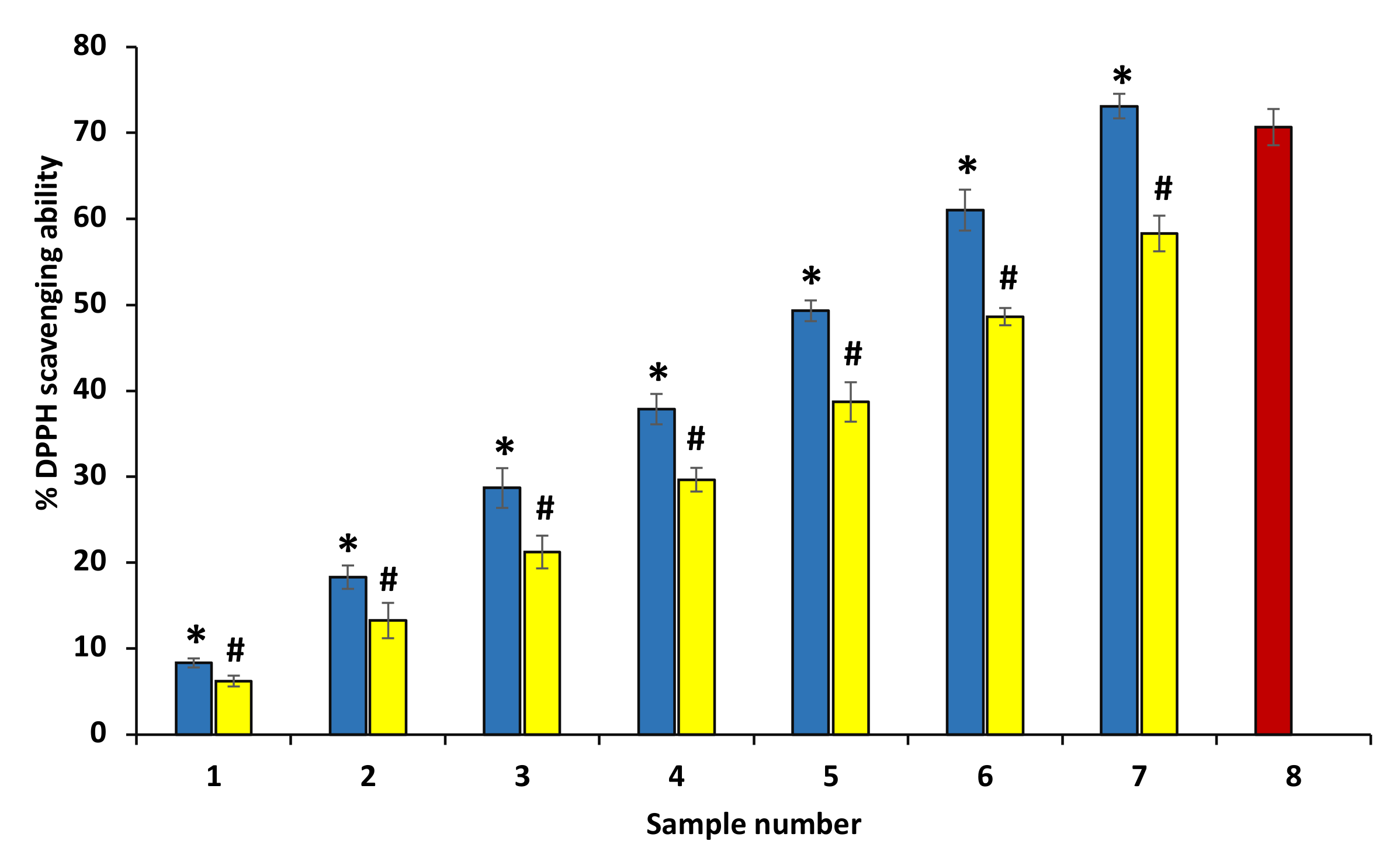
3.4. DPPH Radical Scavenging Assay
3.5. Protein Denaturation Inhibition—An Evaluation of Anti-Inflammatory Activity
3.6. Anti-Proteinase Activity
3.7. Inhibition of Egg Albumin Denaturation Inhibition to Evaluate In Vitro Anti-Arthritic Activity of P. emblica and A. indica Extract
3.8. Membrane Stabilization Test
3.8.1. Heat-Induced Hemolysis
3.8.2. Effect of P. emblica and A. indica Extract on Browning
3.8.3. Effect of Ethanol P. emblica and A. indica on Protein Aggregation Index
3.9. Congo Red Assay
3.10. UV-Absorption Studies
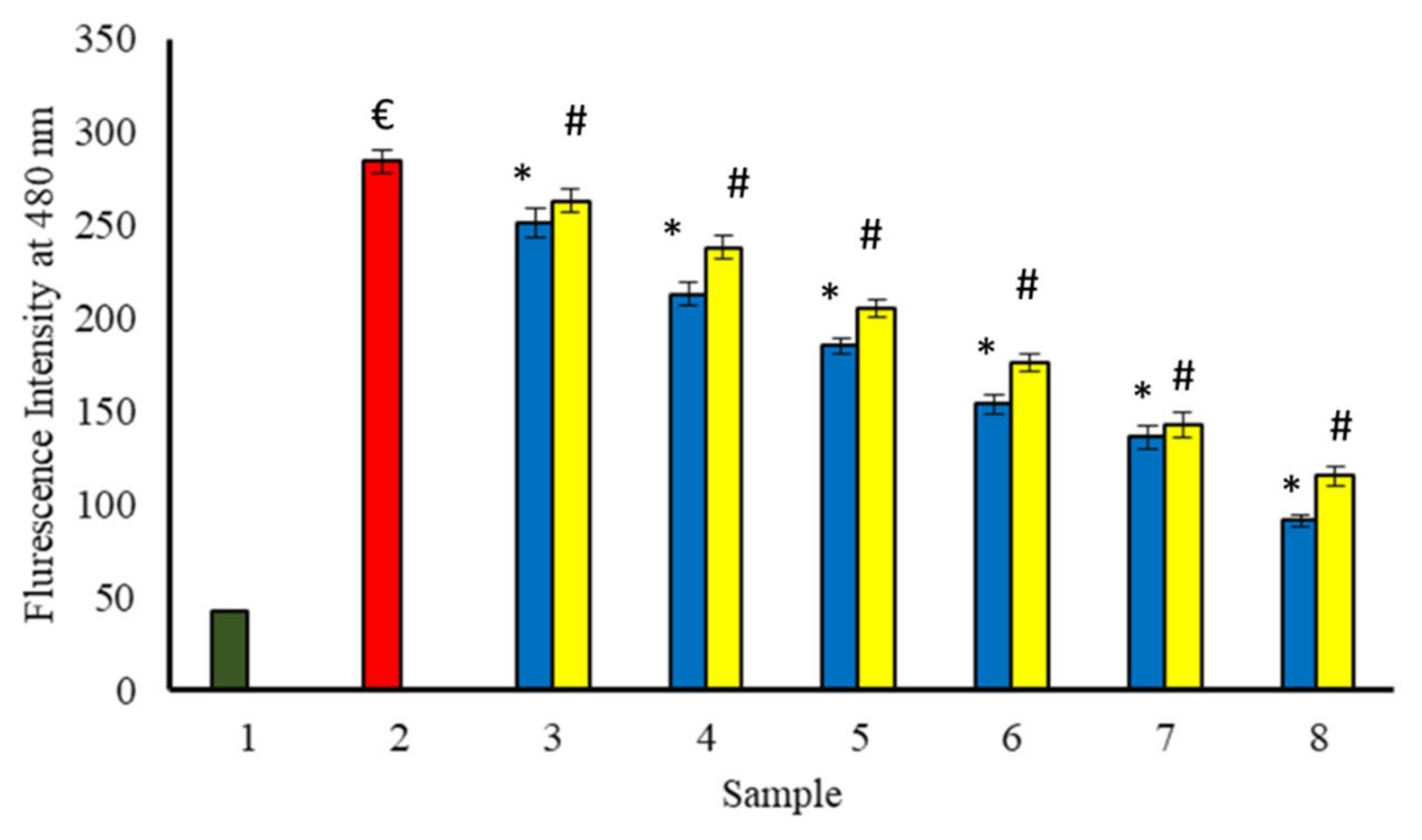
3.11. Effect of Extracts on the Formation of Fluorescent AGEs
3.12. Effect of Extracts on the Formation of Glycation Induced Fibrils
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ajith, T.A.; Janardhanan, K.K. Indian medicinal mushrooms as a source of antioxidant and antitumor agents. J. Clin. Biochem. Nutr. 2007, 40, 157–162. [Google Scholar] [CrossRef] [Green Version]
- Anwar, S.; Almatroudi, A.; Allemailem, K.S.; Jacob Joseph, R.; Khan, A.A.; Rahmani, A.H. Protective Effects of Ginger Extract against Glycation and Oxidative Stress-Induced Health Complications: An In Vitro Study. Processes 2020, 8, 468. [Google Scholar] [CrossRef]
- Saha, S.K.; Lee, S.B.; Won, J.; Choi, H.Y.; Kim, K.; Yang, G.M.; Dayem, A.A.; Cho, S.G. Correlation between Oxidative Stress, Nutrition, and Cancer Initiation. Int. J. Mol. Sci. 2017, 18, 1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storz, P. Oxidative Stress in Cancer. In Oxidative Stress and Redox Regulation; Jakob, U., Reichmann, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 427–448. [Google Scholar]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antiox Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coussens, L.M.; Werb, Z. Inflammation and cancer. Nature 2002, 420, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Anwar, S.; Khan, M.A.; Sadaf, A.; Younus, H. A structural study on the protection of glycation of superoxide dismutase by thymoquinone. Int. J. Biol. Macromol. 2014, 69, 476–481. [Google Scholar] [CrossRef]
- Khan, M.A.; Anwar, S.; Alijarbou, A.A.; Al-Orainy, M.; Aldebasi, Y.H.; Islam, S.; Younus, H. Protective effect of thymoquinone on glucose or methylglyoxal-induced glycation of superoxide dismutase. Int J Biol Macromol. 2014, 65, 16–20. [Google Scholar] [CrossRef]
- Younus, H.; Anwar, S. Prevention of non-enzymatic glycosylation (glycation): Implication in the treatment of diabetic complication. Int. J. Health Sci. 2016, 10, 261–277. [Google Scholar] [CrossRef]
- Younus, H.; Anwar, S. Antiglycating activity of Aloe vera gel extract and its active component aloin. Proteins Proteom. 2018, 9, 115–125. [Google Scholar]
- Anwar, S.; Younus, H. Inhibitory effect of alliin from Allium sativum on the glycation of superoxide dismutase. Int. J. Biol. Macromol. 2017, 103, 182–193. [Google Scholar] [CrossRef]
- Anwar, S.; Younus, H. Antiglycating potential of ellagic acid against glucose and methylglyoxal induced glycation of superoxide dismutase. J. Protein Proteom. 2017, 8, 1–12. [Google Scholar]
- Anwar, S.; Khan, S.; Almatroudi, A.; Khan, A.A.; Alsahli, M.A.; Almatroodi, S.A.; Rahmani, A.H. A review on mechanism of inhibition of advanced glycation end products formation by plant derived polyphenolic compounds. Mol. Biol. Rep. 2021, 48, 787–805. [Google Scholar] [CrossRef] [PubMed]
- Ansari, N.A.; Rasheed, Z. Non-Enzymatic glycation of proteins: From Diabetes and cancer. Biochem. Suppl. Ser. B Biomed. Chem. 2009, 56, 168–178. [Google Scholar] [CrossRef]
- Harris, C.S.; Beaulieu, L.-P.; Fraser, M.-H.; McIntyre, K.L.; Owen, P.L.; Martineau, L.C.; Cuerrier, A.; Johns, T.; Haddad, P.S.; Bennett, S.A.L.; et al. Inhibition of advanced glycation end product formation by medicinal plant extracts correlates with phenolic metabolites and antioxidant activity. Planta Med. 2011, 77, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Schröter, D.; Höhn, A. Role of Advanced Glycation End Products in Carcinogenesis and their Therapeutic Implications. Curr. Pharm. Des. 2018, 24, 5245–5251. [Google Scholar] [CrossRef]
- Turner, D.P. Advanced glycation end-products: A biological consequence of lifestyle contributing to cancer disparity. Cancer Res. 2015, 75, 1925–1929. [Google Scholar] [CrossRef] [Green Version]
- Gangemi, S.; Allegra, A.; Aguennouz, M.; Alonci, A.; Speciale, A.; Cannavò, A.; Cristani, M.; Russo, S.; Spatari, G.; Alibrandi, A.; et al. Musolino, relationship between advanced oxidation protein products, advanced glycation end products, and S-nitrosylated proteins with biological risk and MDR-1 polymorphisms in patients affected by B-chronic lymphocytic leukemia. Cancer Investig. 2012, 30, 20–26. [Google Scholar] [CrossRef]
- Khan, M.S.; Tabrez, S.; Al-Okail, M.S.; Shaik, G.M.; Bhat, S.A.; Rehman, T.M.; Husain, F.M.; Al Ajmi, M.F. Non-enzymatic glycation of protein induces cancer cell proliferation and its inhibition by quercetin: Spectroscopic, cytotoxicity and molecular docking studies. J. Biomol. Struct. Dyn. 2020, 39, 777–786. [Google Scholar] [CrossRef]
- Cragg GM, Newman DJ. Plants as a source of anti-cancer agents. J. Ethnopharmacol. 2005, 100, 72–79. [CrossRef] [Green Version]
- Sripanidkulchai, B.; Fangkrathok, N. Antioxidant, antimutagenic and antibacterial activities of extracts from Phyllanthus emblica branches. Songklanakarin J. Sci. Technol. 2014, 36, 669–674. [Google Scholar]
- Kumar, V.S.; Navaratnam, V. Neem (Azadirachta indica): Prehistory to contemporary medicinal uses to humankind. Asian Pac. J. Trop. Biomed. 2013, 3, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Ordonez, A.; Gomez, J.; Vattuone, M. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Taie, H.A.A.; Salama, Z.A.-E.R.; Radwan, S. Potential activity of basil plants as a source of antioxidants and anticancer agents as affected by organic and bio-organic fertilization. Not. Bot. Horti. Agrobo. 2010, 38, 119127. [Google Scholar]
- Bhalodia, N.R.; Nariya, P.B.; Acharya, R.N.; Shukla, V.J. In vitro antioxidant activity of hydro alcoholic extract from the fruit pulp of Cassia fistula Linn. Ayu 2013, 34, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Ruch, R.J.; Cheng, S.J.; Klaunig, J.E. Prevention of cytotoxicity and inhibition of intercellular communication by antioxidant catechins isolated from Chinese green tea. Carcinogeesis 1989, 10, 1003–1008. [Google Scholar] [CrossRef]
- Mizushima, Y.; Kobayashi, M. Interaction of anti-inflammatory drugs with serum preoteins, especially with some biologically active proteins. J. Pharma. Pharmacol. 1968, 20, 169–173. [Google Scholar] [CrossRef]
- Sakat, S.S.; Juvekar, A.R.; Gambhire, M.N. In Vitro antioxidant and anti-inflammatory activity of methanol extract of Oxalis corniculata linn. Int. J. Pharm. Pharmaceut. Sci. 2010, 2, 146–155. [Google Scholar]
- Oyedepo, O.O.; Femurewa, A.J. Anti-protease and membrane stabilizing activities of extracts of Fagra zanthoxiloides, Olax subscorpioides and Tetrapleura tetraptera. Int. J. Pharmacong. 1995, 33, 65–69. [Google Scholar]
- Chanda, S.; Juvekar, A. In vitro anti-inflammatory activity of syringic acid. Int J Pharm Pharm Sci. 2019, 11, 71–73. [Google Scholar] [CrossRef]
- Brownlee, M.; Vlassara, H.; Kooney, A.; Ulrich, P.; Cerami, A. Aminoguanidine prevents diabetes induced arterial wall protein cross-linking. Science 1968, 232, 1629–1632. [Google Scholar] [CrossRef]
- Kumar, D.; Ali, A. Antiglycation and antiaggregation potential of thymoquinone. Nat. Volatiles Essent. Oils. 2019, 6, 25–33. [Google Scholar]
- Klunk, W.E.; Jacob, R.F.; Mason, R.P. Quantifying amyloid beta-peptide (Abeta) aggregation using the Congo red-Abeta (CR-abeta) spectrophotometric assay. Anal Biochem. 1999, 266, 66–76. [Google Scholar] [CrossRef]
- Dias, M.I.; Sousa, M.J.; Alves, R.C.; Ferreira, I.C.F.R. Exploring plant tissue culture to improve the production of phenolic compounds: A review. Ind. Crop. Prod. 2016, 82, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Ajandouz, E.H.; Tchiakpe, L.S.; Ore, F.D.; Benajiba, A.; Puigserver, A. Effect of pH on caramelization and Maillard reaction kinetics in fructose-lysine model systems. J. Food Sci. 2011, 66, 926–931. [Google Scholar] [CrossRef]
- Nair, A.; Balasaravanan, T.; Jadhav, S.; Mohan, V.; Kumar, C. Harnessing the antibacterial activity of Quercus infectoria and Phyllanthus emblica against antibiotic-resistant Salmonella Typhi and Salmonella Enteritidis of poultry origin. Vet. World 2020, 13, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Acharya, C.K. Ethnicity and Scientific validation of West Bengal Amla (Phyllanthus emblica L.) with special reference to GC-MS screening. Int. J. Exp. Res. Rev. 2016, 3, 51–59. [Google Scholar]
- Balasubramanian, S.; Ganesh, D.; Surya Narayana, V.V.S. Gc-Ms Analysis of Phytocomponents in the Methanolic Extract of Azadirachta Indica (Neem). Int. J. Pharm. Bio. Sci. 2014, 5, 258–262. [Google Scholar]
- Marar, T. Amelioration of glucose induced hemolysis human erythrocytes by vitamin E. Chem. -Biol. Interact. 2011, 193, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Tanaka, T.; Kawaguchi, W.; Fang, N.R.; Ozeki, M.; Akatsuka, S.; Hiai, H.; Arouma, O.I.; Bahorun, T. Effects of the phenolic contents of Mauritian endemic plant extracts on promoter activities of antioxidant enzymes. Free Radic. Res. 2003, 37, 1215–1224. [Google Scholar] [CrossRef]
- Pourahmad, J.; Salimi, A.; Seydi, E. Role of Oxygen Free Radicals in Cancer Development and Treatment. Free Rad. Dis. Intech Open 2016. [Google Scholar] [CrossRef] [Green Version]
- El Abed, H.; Chakroun, M.; Abdelkafi-Koubaa, Z.; Drira, N.; Marrakchi, N.; Mejdoub, H.; Khemakhem, B. Antioxidant, Anti-Inflammatory, and Antitumoral Effects of Aqueous Ethanolic Extract from Phoenix dactylifera L. Parthenocarpic Dates. Biomed. Res. Int. 2018. [Google Scholar] [CrossRef] [Green Version]
- Scanlon, V.C.; Sanders, T. Essentials of Anatomy and Physiology, 6th ed.; F.A. Davis Company: Philadelphia, PA, USA, 2010; p. 287. [Google Scholar]
- Brown, J.H.; Mackey, H.K. Inhibition of heat-induced denaturation of serum proteins by mixtures of non-steroidal anti-inflammatory agents and amino acids. Proc. Soc. Exp. Biol. Med. 1968, 128, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Havsteen, B.H. The biochemistry and medical significance of the flavonoids. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar] [CrossRef]
- Srividya, S.; Sridevi, G. Anti-arthritic and anti-inflammatory activity of ethanolic leaf extract of Ormocarpum Sennoides. Int. J. Pharm. Pharm. Sci. 2016, 8, 117–121. [Google Scholar]
- Xie, W.; Du, L. Diabetes is an inflammatory disease: Evidence from traditional Chinese medicines. Diabetes Obes. Metabol. 2011, 13, 289–301. [Google Scholar] [CrossRef]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 119. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preliminary Screening | P. emblica | A. indica |
|---|---|---|
| Weight of dry powder | 100 g | 100 g |
| Yield | 8.79% | 7.19% |
| Extract | Ethanol | Ethanol |
| Color | Reddish brown | Brown |
| Odor | No specific | No specific |
| Texture | Sticky | Powder |
| Flavonoid (alkaline reagent test) | + | + |
| Phenolic compounds (FeCl3 test) | + | + |
| Concentration (µg/mL) | P. emblica | A. indica | ||
|---|---|---|---|---|
| Percentage Protection | S. E. M. | Percentage Protection | S. E. M. | |
| 100 | 9.13 | 0.1 | 7.21 | 0.003 |
| 200 | 15.91 | 1 | 11.68 | 0.07 |
| 300 | 24.62 | 1.321 | 18.47 | 0.256 |
| 400 | 30.73 | 0.038 | 25.47 | 1.18 |
| 500 | 45.56 | 1.028 | 44.16 | 0.76 |
| 600 | 54.441 | 0.631 | 50.217 | 0.313 |
| Concentration (µg/mL) | P. emblica | A. indica | ||
|---|---|---|---|---|
| Percentage Protection | S. E. M. | Percentage Protection | S. E. M. | |
| 100 | 9.43 | 0.133 | 8.01 | 0.033 |
| 200 | 17.98 | 1.079 | 15.63 | 0.078 |
| 300 | 25.69 | 0.321 | 23.24 | 1.346 |
| 400 | 35.73 | 1.058 | 33.47 | 0.588 |
| 500 | 45.56 | 0.003 | 43.16 | 1.903 |
| 600 | 59.28 | 0.711 | 57.27 | 0.283 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
A. Alsahli, M.; Anwar, S.; Alzahrani, F.M.; Almatroudi, A.; Alfheeaid, H.; Khan, A.A.; Allemailem, K.S.; Almatroodi, S.A.; Rahmani, A.H. Health Promoting Effect of Phyllanthus emblica and Azadiractha indica against Advanced Glycation End Products Formation. Appl. Sci. 2021, 11, 8819. https://doi.org/10.3390/app11198819
A. Alsahli M, Anwar S, Alzahrani FM, Almatroudi A, Alfheeaid H, Khan AA, Allemailem KS, Almatroodi SA, Rahmani AH. Health Promoting Effect of Phyllanthus emblica and Azadiractha indica against Advanced Glycation End Products Formation. Applied Sciences. 2021; 11(19):8819. https://doi.org/10.3390/app11198819
Chicago/Turabian StyleA. Alsahli, Mohammed, Shehwaz Anwar, Faisal M. Alzahrani, Ahmad Almatroudi, Hani Alfheeaid, Amjad A. Khan, Khaled S. Allemailem, Saleh A. Almatroodi, and Arshad Husain Rahmani. 2021. "Health Promoting Effect of Phyllanthus emblica and Azadiractha indica against Advanced Glycation End Products Formation" Applied Sciences 11, no. 19: 8819. https://doi.org/10.3390/app11198819
APA StyleA. Alsahli, M., Anwar, S., Alzahrani, F. M., Almatroudi, A., Alfheeaid, H., Khan, A. A., Allemailem, K. S., Almatroodi, S. A., & Rahmani, A. H. (2021). Health Promoting Effect of Phyllanthus emblica and Azadiractha indica against Advanced Glycation End Products Formation. Applied Sciences, 11(19), 8819. https://doi.org/10.3390/app11198819