
Recent Advances in the Heterologous Biosynthesis of Natural Products from Streptomyces

 , ,
, , 

Abstract
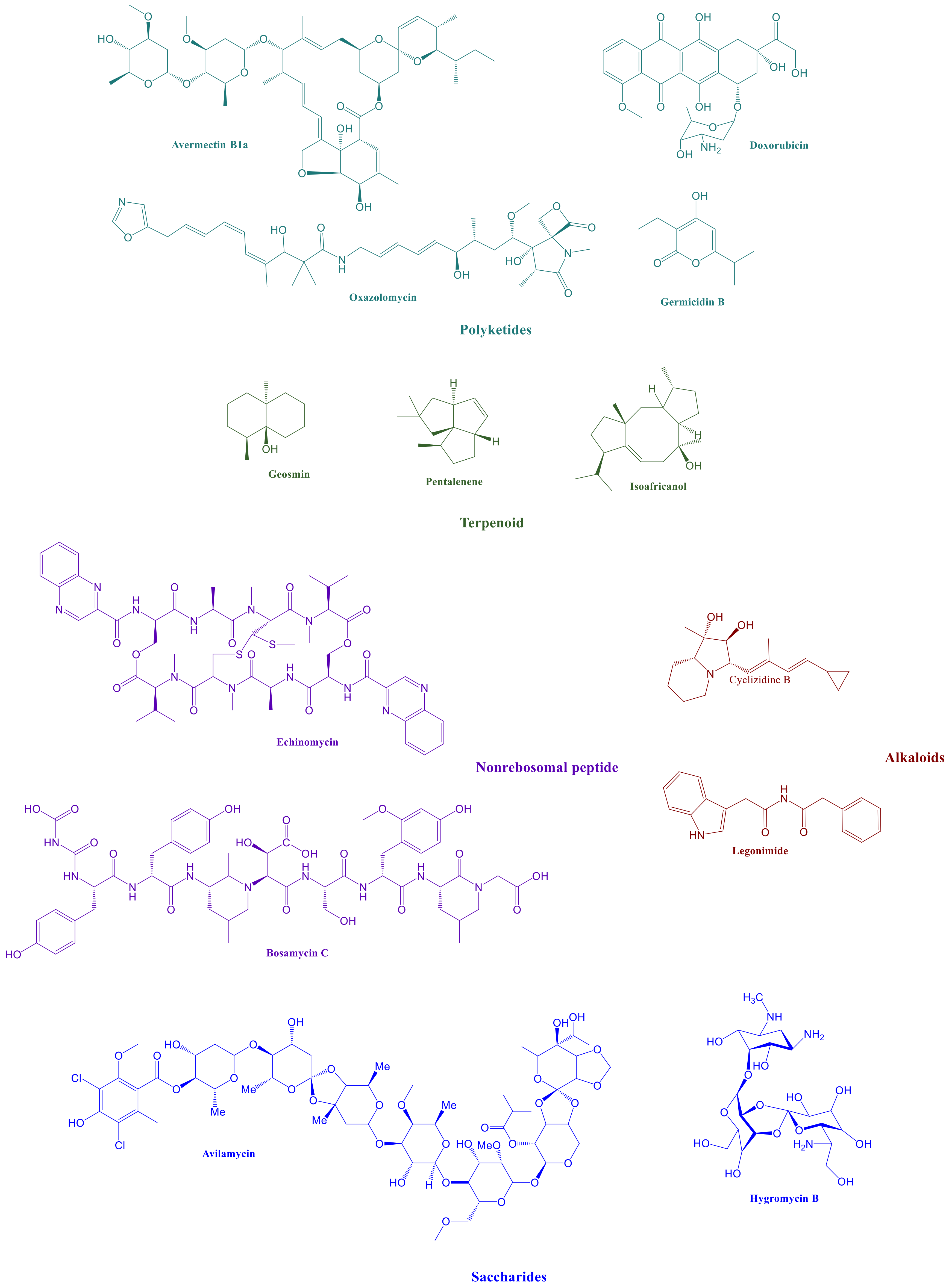
1. Introduction
2. In Silico Prediction of BGCs
3. Selecting a Suitable Host for Heterologous Streptomyces Products
3.1. Streptomyces
3.2. Other Hosts
4. Host Engineering Approaches
4.1. Host Cleaning
4.2. Transcription Terminators
4.3. Riboswitches
4.4. tRNA Engineering
4.5. Regulators
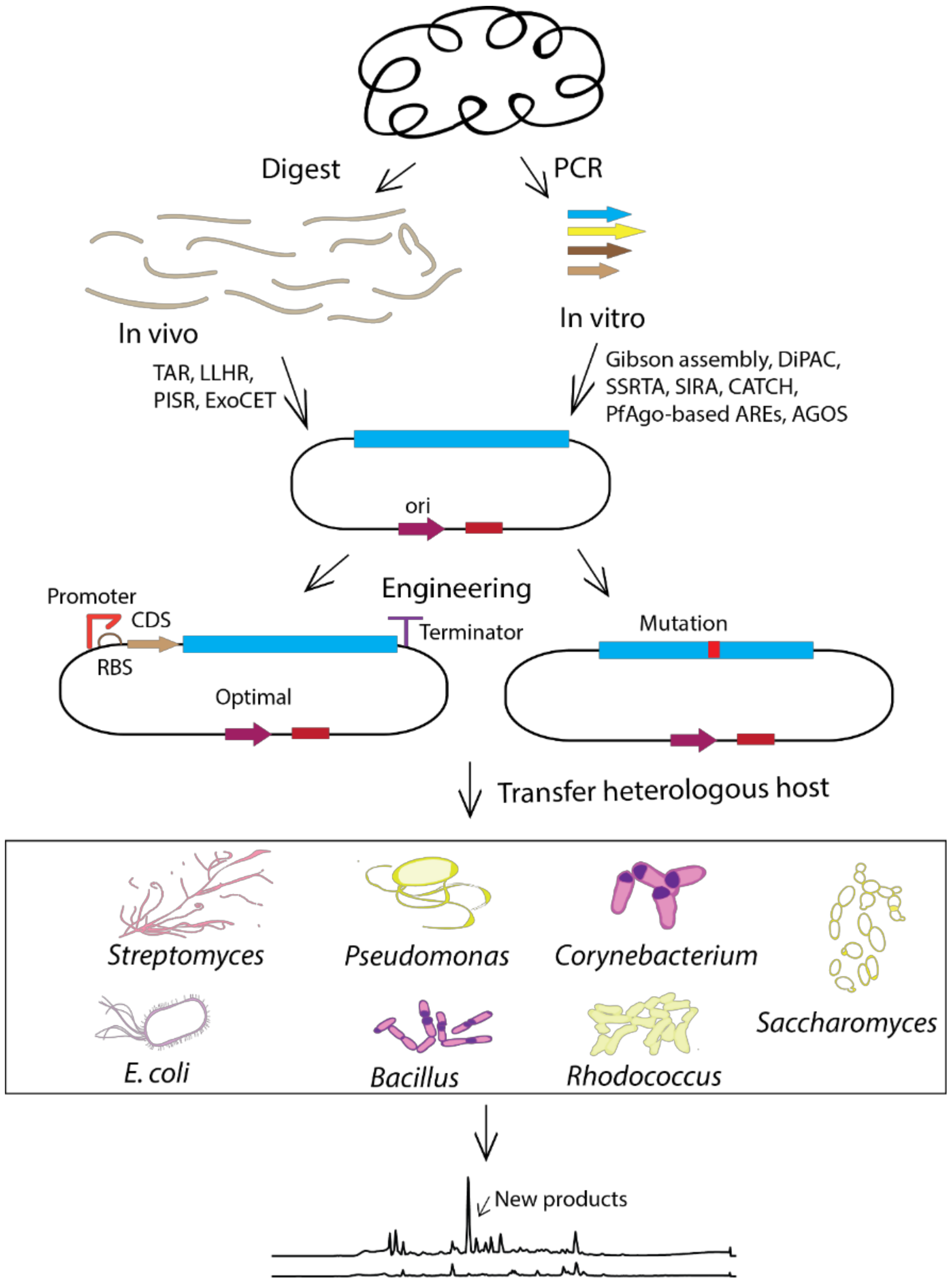
5. Strategies for the Construction of a Biosynthetic Gene Cluster
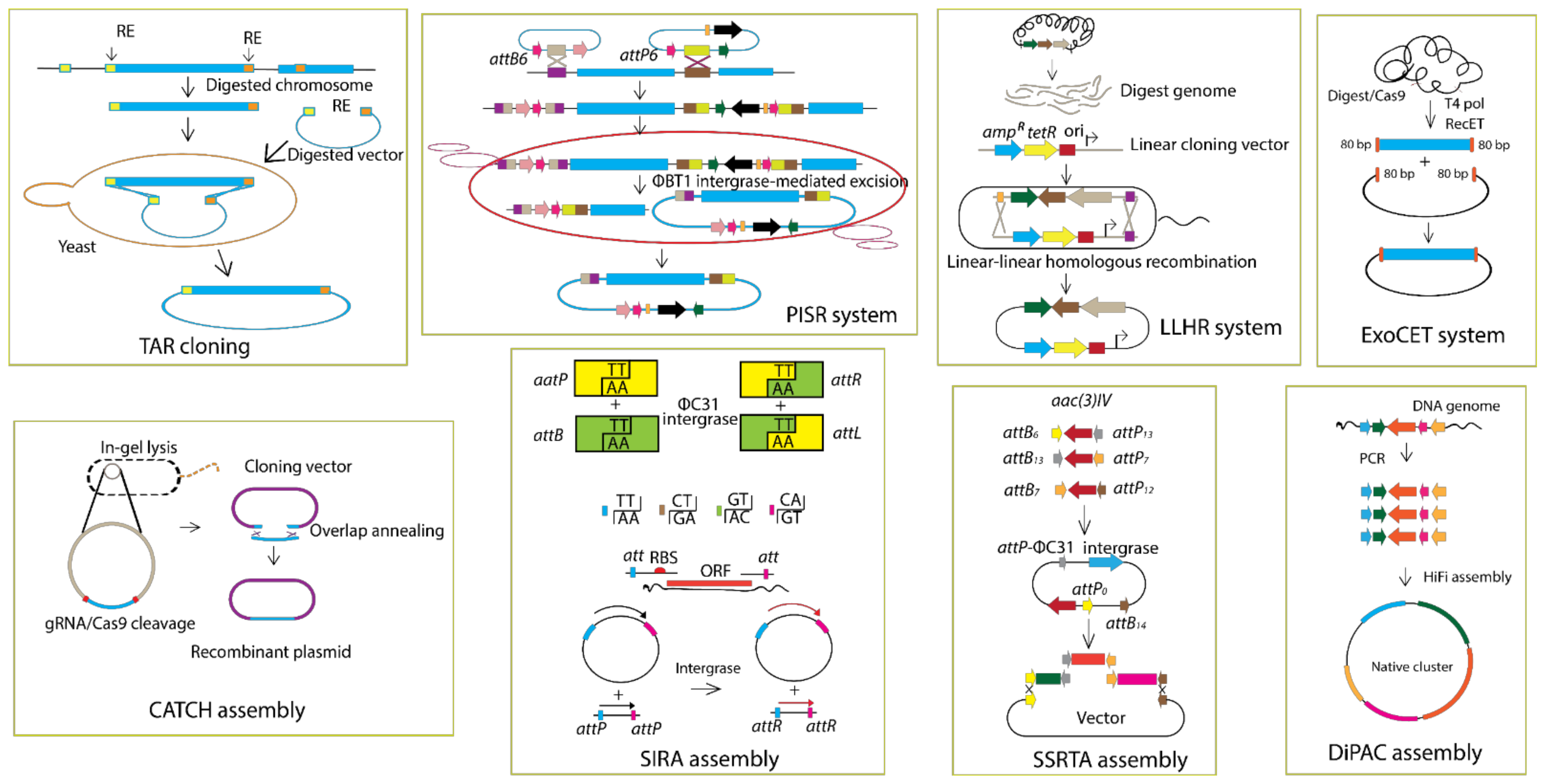
5.1. Vectors and Cloning Methods
5.2. Promoter Engineering
5.3. RBS Tuning
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Nguyen, C.T.; Dhakal, D.; Pham, V.T.T.; Nguyen, H.T.; Sohng, J.K. Recent Advances in Strategies for Activation and Discovery/Characterization of Cryptic Biosynthetic Gene Clusters in Streptomyces. Microorganisms 2020, 8, 616. [Google Scholar] [CrossRef]
- Deng, Q.; Zhou, L.; Luo, M.; Deng, Z.; Zhao, C. Heterologous expression of Avermectins biosynthetic gene cluster by construction of a Bacterial Artificial Chromosome library of the producers. Synth. Syst. Biotechnol. 2017, 2, 59–64. [Google Scholar] [CrossRef]
- Xu, Z.F.; Bo, S.T.; Wang, M.J.; Shi, J.; Jiao, R.H.; Sun, Y.; Xu, Q.; Tan, R.X.; Ge, H.M. Discovery and biosynthesis of bosamycins from Streptomyces sp. 120454. Chem. Sci. 2020, 11, 9237–9245. [Google Scholar] [CrossRef]
- Martín-Sánchez, L.; Singh, K.S.; Avalos, M.; Van Wezel, G.P.; Dickschat, J.S.; Garbeva, P. Phylogenomic analyses and distribution of terpene synthases among Streptomyces. Beilstein J. Org. Chem. 2019, 15, 1181–1193. [Google Scholar] [CrossRef]
- McCulloch, K.M.; McCranie, E.K.; Smith, J.A.; Sarwar, M.; Mathieu, J.L.; Gitschlag, B.L.; Du, Y.; Bachmannb, B.O.; Iversona, T.M. Oxidative cyclizations in orthosomycin biosynthesis expand the known chemistry of an oxygenase superfamily. Proc. Natl. Acad. Sci. USA 2015, 112, 11547–11552. [Google Scholar] [CrossRef]
- Fang, Q.; Maglangit, F.; Mugat, M.; Urwald, C.; Kyeremeh, K.; Deng, H. Targeted isolation of indole alkaloids from Streptomyces sp. CT37. Molecules 2020, 25, 1108. [Google Scholar] [CrossRef]
- Tetzlaff, C.N.; You, Z.; Cane, D.E.; Takamatsu, S.; Omura, S.; Ikeda, H. A gene cluster for biosynthesis of the sesquiterpenoid antibiotic pentalenolactone in Streptomyces avermitilis. Biochemistry 2006, 45, 6179–6186. [Google Scholar] [CrossRef]
- Pokhrel, A.R.; Chaudhary, A.K.; Nguyen, H.T.; Dhakal, D.; Le, T.T.; Shrestha, A.; Liou, K.; Sohng, J.K. Overexpression of a pathway specific negative regulator enhances production of daunorubicin in bldA deficient Streptomyces peucetius ATCC 27952. Microbiol. Res. 2016, 192, 96–102. [Google Scholar] [CrossRef]
- Singh, B.; Oh, T.J.; Sohng, J.K. Exploration of geosmin synthase from Streptomyces peucetius ATCC 27952 by deletion of doxorubicin biosynthetic gene cluster. J. Ind. Microbiol. Biotechnol. 2009, 36, 1257–1265. [Google Scholar] [CrossRef]
- Helfrich, E.J.N.; Piel, J. Biosynthesis of polyketides by trans-AT polyketide synthases. Nat. Prod. Rep. 2016, 33, 231–316. [Google Scholar] [CrossRef]
- Ahmed, Y.; Rebets, Y.; Estévez, M.R.; Zapp, J.; Myronovskyi, M.; Luzhetskyy, A. Engineering of Streptomyces lividans for heterologous expression of secondary metabolite gene clusters. Microb. Cell Factories 2020, 19, 5. [Google Scholar] [CrossRef]
- Migita, A.; Watanabe, M.; Hirose, Y.; Watanabe, K.; Tokiwano, T.; Kinashi, H.; Oikawa, H. Identification of a gene cluster of polyether antibiotic lasalocid from Streptomyces lasaliensis. Biosci. Biotechnol. Biochem. 2009, 73, 169–176. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Li, J.Q.; Zhang, H.J.; Ding, W.J.; Ma, Z.J. Cyclizidine-Type Alkaloids from Streptomyces sp. HNA39. J. Nat. Prod. 2018, 81, 394–399. [Google Scholar] [CrossRef]
- Arenz, S.; Juette, M.F.; Graf, M.; Nguyen, F.; Huter, P.; Polikanov, Y.S.; Blanchard, S.C.; Wilson, D.N. Structures of the orthosomycin antibiotics avilamycin and evernimicin in complex with the bacterial 70S ribosome. Proc. Natl. Acad. Sci. USA 2016, 113, 7527–7532. [Google Scholar] [CrossRef]
- Mann, R.L.; Bromer, W.W. The Isolation of a Second Antibiotic from Streptomyces hygroscopicus. J. Am. Chem. Soc. 1958, 80, 2714–2716. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Luzhetskyy, A. Heterologous production of small molecules in the optimized: Streptomyces hosts. Nat. Prod. Rep. 2019, 36, 1281–1294. [Google Scholar] [CrossRef]
- Lachance, H.; Wetzel, S.; Kumar, K.; Waldmann, H. Charting, navigating, and populating natural product chemical space for drug discovery. J. Med. Chem. 2012, 55, 5989–6001. [Google Scholar] [CrossRef]
- Lahlou, M. The Success of Natural Products in Drug Discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef]
- Waldetoft, K.W.; Gurney, J.; Lachance, J.; Hoskisson, P.A.; Brown, S.P. Evolving antibiotics against resistance: A potential platform for natural product development? mBio 2019, 10, e02946-19. [Google Scholar] [CrossRef]
- Moussa, M.; Ebrahim, W.; Bonus, M.; Gohlke, H.; Mándi, A.; Kurtán, T.; Hartmann, R.; Kalscheuer, R.; Lin, W.; Liu, Z.; et al. Co-culture of the fungus Fusarium tricinctum with Streptomyces lividans induces production of cryptic naphthoquinone dimers. RSC Adv. 2019, 9, 1491–1500. [Google Scholar] [CrossRef]
- Zhao, Z.; Shi, T.; Xu, M.; Brock, N.L.; Zhao, Y.L.; Wang, Y.; Deng, Z.; Pang, X.; Tao, M. Hybrubins: Bipyrrole tetramic acids obtained by crosstalk between a truncated undecylprodigiosin pathway and heterologous tetramic acid biosynthetic genes. Org. Lett. 2016, 18, 572–575. [Google Scholar] [CrossRef]
- Pyeon, H.R.; Nah, H.J.; Kang, S.H.; Choi, S.S.; Kim, E.S. Heterologous expression of pikromycin biosynthetic gene cluster using Streptomyces artificial chromosome system. Microb. Cell Factories 2017, 16, 96. [Google Scholar] [CrossRef]
- Linares-Otoya, L.; Linares-Otoya, V.; Armas-Mantilla, L.; Blanco-Olano, C.; Crüsemann, M.; Ganoza-Yupanqui, M.L.; Campos-Florian, J.; König, G.M.; Schäberle, T.F. Identification and heterologous expression of the kocurin biosynthetic gene cluster. Microbiology 2017, 163, 1409–1414. [Google Scholar] [CrossRef]
- Choi, S.; Nah, H.J.; Choi, S.; Kim, E.S. Heterologous expression of daptomycin biosynthetic gene cluster via Streptomyces artificial chromosome vector system. J. Microbiol. Biotechnol. 2019, 29, 1931–1937. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, H.; Haltli, B.; Kulowski, K.; Muszynska, E.; Feng, X.; Summers, M.; Young, M.; Graziani, E.; Koehn, F.; et al. Rapid Cloning and Heterologous Expression of the Meridamycin Biosynthetic Gene Cluster Using a Versatile Escherichia coli–Streptomyces Artificial Chromosome Vector, pSBAC. J. Nat. Prod. 2009, 72, 389–395. [Google Scholar] [CrossRef]
- Kepplinger, B.; Morton-Laing, S.; Seistrup, K.H.; Marrs, E.C.L.; Hopkins, A.P.; Perry, J.D.; Strahl, H.; Hall, M.J.; Errington, J.; Ellis Allenby, N.E. Mode of Action and Heterologous Expression of the Natural Product Antibiotic Vancoresmycin. ACS Chem. Biol. 2018, 13, 207–214. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, G.; Ding, X. Tandem assembly of the epothilone biosynthetic gene cluster by in vitro site-specific recombination. Sci. Rep. 2011, 1, 141. [Google Scholar] [CrossRef]
- Kim, J.H.; Feng, Z.; Bauer, J.D.; Kallifidas, D.; Calle, P.Y.; Brady, S.F. Cloning large natural product gene clusters from the environment: Piecing environmental DNA gene clusters back together with TAR. Biopolymers 2010, 93, 833–844. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Ziemert, N.; Podell, S.; Penn, K.; Badger, J.H.; Allen, E.; Jensen, P.R. The natural product domain seeker NaPDoS: A phylogeny based bioinformatic tool to classify secondary metabolite gene diversity. PLoS ONE 2012, 7, e34064. [Google Scholar] [CrossRef]
- Ziemert, N.; Lechner, A.; Wietz, M.; Millań-Aguiñaga, N.; Chavarria, K.L.; Jensen, P.R. Diversity and evolution of secondary metabolism in the marine actinomycete genus Salinispora. Proc. Natl. Acad. Sci. USA 2014, 111, E1130–E1139. [Google Scholar] [CrossRef]
- Röttig, M.; Medema, M.H.; Blin, K.; Weber, T.; Rausch, C.; Kohlbacher, O. NRPSpredictor2—A web server for predicting NRPS adenylation domain specificity. Nucleic Acids Res. 2011, 39, 362–367. [Google Scholar] [CrossRef]
- Cimermancic, P.; Medema, M.H.; Claesen, J.; Kurita, K.; Wieland Brown, L.C.; Mavrommatis, K.; Pati, A.; Godfrey, P.A.; Koehrsen, M.; Clardy, J.; et al. Insights into secondary metabolism from a global analysis of prokaryotic biosynthetic gene clusters. Cell 2014, 158, 412–421. [Google Scholar] [CrossRef]
- Skinnider, M.A.; Johnston, C.W.; Gunabalasingam, M.; Merwin, N.J.; Kieliszek, A.M.; MacLellan, R.J.; Li, H.; Ranieri, M.R.M.; Webster, A.L.H.; Cao, M.P.T.; et al. Comprehensive prediction of secondary metabolite structure and biological activity from microbial genome sequences. Nat. Commun. 2020, 11, 6058. [Google Scholar] [CrossRef]
- Starcevic, A.; Zucko, J.; Simunkovic, J.; Long, P.F.; Cullum, J.; Hranueli, D. ClustScan: An integrated program package for the semi-automatic annotation of modular biosynthetic gene clusters and in silico prediction of novel chemical structures. Nucleic Acids Res. 2008, 36, 6882–6892. [Google Scholar] [CrossRef]
- Li, M.H.T.; Ung, P.M.U.; Zajkowski, J.; Garneau-Tsodikova, S.; Sherman, D.H. Automated genome mining for natural products. BMC Bioinform. 2009, 10, 185. [Google Scholar] [CrossRef]
- Cruz-Morales, P.; Kopp, J.F.; Martínez-Guerrero, C.; Yáñez-Guerra, L.A.; Selem-Mojica, N.; Ramos-Aboites, H.; Feldmann, J.; Barona-Gómez, F. Phylogenomic analysis of natural products biosynthetic gene clusters allows discovery of arseno-organic metabolites in model Streptomycetes. Genome Biol. Evol. 2016, 8, 1906–1916. [Google Scholar] [CrossRef]
- Alanjary, M.; Kronmiller, B.; Adamek, M.; Blin, K.; Weber, T.; Huson, D.; Philmus, B.; Ziemert, N. The Antibiotic Resistant Target Seeker (ARTS), an exploration engine for antibiotic cluster prioritization and novel drug target discovery. Nucleic Acids Res. 2017, 45, W42–W48. [Google Scholar] [CrossRef]
- Khater, S.; Gupta, M.; Agrawal, P.; Sain, N.; Prava, J.; Gupta, P.; Grover, M.; Kumar, N.; Mohanty, D. SBSPKSv2: Structure-based sequence analysis of polyketide synthases and non-ribosomal peptide synthetases. Nucleic Acids Res. 2017, 45, W72–W79. [Google Scholar] [CrossRef]
- Van Heel, A.J.; De Jong, A.; Song, C.; Viel, J.H.; Kok, J.; Kuipers, O.P. BAGEL4: A user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Res. 2018, 46, W278–W281. [Google Scholar] [CrossRef]
- Santos-Aberturas, J.; Chandra, G.; Frattaruolo, L.; Lacret, R.; Pham, T.H.; Vior, N.M.; Eyles, T.H.; Truman, A.W. Uncovering the unexplored diversity of thioamidated ribosomal peptides in Actinobacteria using the RiPPER genome mining tool. Nucleic Acids Res. 2019, 47, 4624–4637. [Google Scholar] [CrossRef]
- De Los Santos, E.L.C. NeuRiPP: Neural network identification of RiPP precursor peptides. Sci. Rep. 2019, 9, 13406. [Google Scholar] [CrossRef]
- Agrawal, P.; Khater, S.; Gupta, M.; Sain, N.; Mohanty, D. RiPPMiner: A bioinformatics resource for deciphering chemical structures of RiPPs based on prediction of cleavage and cross-links. Nucleic Acids Res. 2017, 45, W80–W88. [Google Scholar] [CrossRef]
- Tietz, J.I.; Schwalen, C.J.; Patel, P.S.; Maxson, T.; Blair, P.M.; Tai, H.C.; Zakai, U.I.; Mitchell, D.A. A new genome-mining tool redefines the lasso peptide biosynthetic landscape. Nat. Chem. Biol. 2017, 13, 470–478. [Google Scholar] [CrossRef]
- Liu, W.; Sun, F.; Hu, Y. Genome Mining-Mediated Discovery of a New Avermipeptin Analogue in Streptomyces actuosus ATCC 25421. ChemistryOpen 2018, 7, 558–561. [Google Scholar] [CrossRef]
- Kloosterman, A.M.; Medema, M.H.; van Wezel, G.P. Omics-based strategies to discover novel classes of RiPP natural products. Curr. Opin. Biotechnol. 2021, 69, 60–67. [Google Scholar] [CrossRef]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; Van Der Hooft, J.J.J.; Van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef]
- MIBiG Logo Minimum Information about a Biosynthetic Gene Cluster. Available online: https://mibig.secondarymetabolites.org/ (accessed on 18 March 2020).
- Luo, Y.; Cobb, R.E.; Zhao, H. Recent Advances in Natural Product Discovery Yunzi. Curr. Opin. Biotechnol. 2014, 30, 230–237. [Google Scholar] [CrossRef]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; Van Der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, 1743–1752. [Google Scholar] [CrossRef]
- Vincenti, F.; Montesano, C.; Di Ottavio, F.; Gregori, A.; Compagnone, D.; Sergi, M.; Dorrestein, P. Molecular Networking: A Useful Tool for the Identification of New Psychoactive Substances in Seizures by LC–HRMS. Front. Chem. 2020, 8, 572952. [Google Scholar] [CrossRef]
- Leipoldt, F.; Santos-Aberturas, J.; Stegmann, D.P.; Wolf, F.; Kulik, A.; Lacret, R.; Popadić, D.; Keinhörster, D.; Kirchner, N.; Bekiesch, P.; et al. Warhead biosynthesis and the origin of structural diversity in hydroxamate metalloproteinase inhibitors. Nat. Commun. 2017, 8, 1965. [Google Scholar] [CrossRef]
- Raheem, D.J.; Tawfike, A.F.; Abdelmohsen, U.R.; Edrada-Ebel, R.A.; Fitzsimmons-Thoss, V. Application of metabolomics and molecular networking in investigating the chemical profile and antitrypanosomal activity of British bluebells (Hyacinthoides non-scripta). Sci. Rep. 2019, 9, 2547. [Google Scholar] [CrossRef]
- Crone, W.J.K.; Vior, N.M.; Santos-Aberturas, J.; Schmitz, L.G.; Leeper, F.J.; Truman, A.W. Dissecting Bottromycin Biosynthesis Using Comparative Untargeted Metabolomics. Angew. Chem. Int. Ed. 2016, 55, 9639–9643. [Google Scholar] [CrossRef]
- Nepal, K.K.; Wang, G. Streptomycetes: Surrogate Hosts for the Genetic Manipulation of Biosynthetic Gene Clusters and Production of Natural Products. Biotechnol. Adv. 2019, 37, 1–20. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkränzer, B.; Stierhof, M.; Petzke, L.; Seiser, T.; Luzhetskyy, A. Identification and heterologous expression of the albucidin gene cluster from the marine strain Streptomyces albus subsp. chlorinus NRRL B-24108. Microorganisms 2020, 8, 237. [Google Scholar] [CrossRef]
- Liu, X.; Liu, D.; Xu, M.; Tao, M.; Bai, L.; Deng, Z.; Pfeifer, B.A.; Jiang, M. Reconstitution of Kinamycin Biosynthesis within the Heterologous Host Streptomyces albus J1074. J. Nat. Prod. 2018, 81, 72–77. [Google Scholar] [CrossRef]
- Jiang, G.; Zhang, Y.; Powell, M.M.; Zhang, P.; Zuo, R.; Zhang, Y.; Kallifidas, D.; Tieu, A.M.; Luesch, H.; Loria, R.; et al. High-Yield Production of Herbicidal Thaxtomins and Thaxtomin Analogs in a Nonpathogenic Streptomyces Strain. Appl. Environ. Microbiol. 2018, 84, e00164-18. [Google Scholar] [CrossRef]
- Shi, Y.; Jiang, Z.; Li, X.; Zuo, L.; Lei, X.; Yu, L.; Wu, L.; Jiang, J.; Hong, B. Biosynthesis of antibiotic chuangxinmycin from Actinoplanes tsinanensis. Acta Pharm. Sin. B 2018, 8, 283–294. [Google Scholar] [CrossRef]
- Chen, W.; Qu, D.; Zhai, L.; Tao, M.; Wang, Y.; Lin, S.; Price, N.P.J.; Deng, Z. Characterization of the tunicamycin gene cluster unveiling unique steps involved in its biosynthesis. Protein Cell 2010, 1, 1093–1105. [Google Scholar] [CrossRef]
- Shuai, H.; Myronovskyi, M.; Nadmid, S.; Luzhetskyy, A. Identification of a biosynthetic gene cluster responsible for the production of a new pyrrolopyrimidine natural product—Huimycin. Biomolecules 2020, 10, 1074. [Google Scholar] [CrossRef]
- Lombó, F.; Velasco, A.; Castro, A.; De La Calle, F.; Braña, A.F.; Sánchez-Puelles, J.M.; Méndez, C.; Salas, J.A. Deciphering the biosynthesis pathway of the antitumor thiocoraline from a marine actinomycete and its expression in two Streptomyces species. ChemBioChem 2006, 7, 366–376. [Google Scholar] [CrossRef]
- Dhakal, D.; Han, J.M.; Mishra, R.; Pandey, R.P.; Kim, T.S.; Rayamajhi, V.; Jung, H.J.; Yamaguchi, T.; Sohng, J.K. Characterization of Tailoring Steps of Nargenicin A1 Biosynthesis Reveals a Novel Analogue with Anticancer Activities. ACS Chem. Biol. 2020, 15, 1370–1380. [Google Scholar] [CrossRef]
- Alduina, R.; Giardina, A.; Gallo, G.; Renzone, G.; Ferraro, C.; Contino, A.; Scaloni, A.; Donadio, S.; Puglia, A.M. Expression in Streptomyces lividans of Nonomuraea genes cloned in an artificial chromosome. Appl. Microbiol. Biotechnol. 2005, 68, 656–662. [Google Scholar] [CrossRef]
- Flinspach, K.; Kapitzke, C.; Tocchetti, A.; Sosio, M.; Apel, A.K. Heterologous expression of the thiopeptide antibiotic GE2270 from Planobispora rosea ATCC 53733 in Streptomyces coelicolor requires deletion of ribosomal genes from the expression construct. PLoS ONE 2014, 9, e90449. [Google Scholar]
- Miyamoto, K.T.; Komatsu, M.; Ikeda, H. Discovery of gene cluster for mycosporine-like amino acid biosynthesis from Actinomycetales microorganisms and production of a novel mycosporine-like amino acid by heterologous expression. Appl. Environ. Microbiol. 2014, 80, 5028–5036. [Google Scholar] [CrossRef]
- Reynolds, K.A.; Luhavaya, H.; Li, J.; Dahesh, S.; Nizet, V.; Yamanaka, K.; Moore, B.S. Isolation and structure elucidation of lipopeptide antibiotic taromycin B from the activated taromycin biosynthetic gene cluster. J. Antibiot. 2018, 71, 333–338. [Google Scholar] [CrossRef]
- Saugar, I.; Molloy, B.; Sanz, E.; Blanca Sánchez, M.; Fernández-Lobato, M.; Jiménez, A. Characterization of the biosynthetic gene cluster (ata) for the A201A aminonucleoside antibiotic from Saccharothrix mutabilis subsp. capreolus. J. Antibiot. 2017, 70, 404–413. [Google Scholar] [CrossRef][Green Version]
- Shepherd, M.D.; Kharel, M.K.; Bosserman, M.A.; Rohr, J. Laboratory maintenance of Streptomyces species. Curr. Protoc. Microbiol. 2010, 18, 10E.1.1–10E.1.8. [Google Scholar] [CrossRef]
- Ro, D.K.; Paradise, E.M.; Quellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940–943. [Google Scholar] [CrossRef]
- Luo, X.; Reiter, M.A.; d’Espaux, L.; Wong, J.; Denby, C.M.; Lechner, A.; Zhang, Y.; Grzybowski, A.T.; Harth, S.; Lin, W.; et al. Complete biosynthesis of cannabinoids and their unnatural analogues in yeast. Nature 2019, 567, 123–126. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, W.; Xiao, G.; Zhan, J. Efficient production of gamma-aminobutyric acid by engineered Saccharomyces cerevisiae with glutamate decarboxylases from Streptomyces. Biotechnol. Appl. Biochem. 2020, 67, 240–248. [Google Scholar] [CrossRef]
- Thomas, F.; Schmidt, C.; Kayser, O. Bioengineering studies and pathway modeling of the heterologous biosynthesis of tetrahydrocannabinolic acid in yeast. Appl. Microbiol. Biotechnol. 2020, 104, 9551–9563. [Google Scholar] [CrossRef]
- Burdett, I.D.; Kirkwood, T.B.; Whalley, J.B. Growth Kinetics of Individual Bacillus subtilis Cells and Correlation with Nucleoid Extension. J. Bacteriol 1986, 167, 219–230. [Google Scholar] [CrossRef]
- Kumpfmüller, J.; Methling, K.; Fang, L.; Pfeifer, B.A.; Lalk, M.; Schweder, T. Production of the polyketide 6-deoxyerythronolide B in the heterologous host Bacillus subtilis. Appl. Microbiol. Biotechnol. 2016, 100, 1209–1220. [Google Scholar] [CrossRef]
- Zobel, S.; Kumpfmüller, J.; Süssmuth, R.D.; Schweder, T. Bacillus subtilis as heterologous host for the secretory production of the non-ribosomal cyclodepsipeptide enniatin. Appl. Microbiol. Biotechnol. 2015, 99, 681–691. [Google Scholar] [CrossRef]
- Jaruszewicz-Błońska, J.; Lipniacki, T. Genetic toggle switch controlled by bacterial growth rate. BMC Syst. Biol. 2017, 11, 117. [Google Scholar] [CrossRef]
- Pontrelli, S.; Chiu, T.Y.; Lan, E.I.; Chen, F.Y.H.; Chang, P.; Liao, J.C. Escherichia coli as a host for metabolic engineering. Metab. Eng. 2018, 50, 16–46. [Google Scholar] [CrossRef]
- Boghigian, B.A.; Pfeifer, B.A. Current status, strategies, and potential for the metabolic engineering of heterologous polyketides in Escherichia coli. Biotechnol. Lett. 2008, 30, 1323–1330. [Google Scholar] [CrossRef]
- Wu, J.; Boghigian, B.A.; Myint, M.; Zhang, H.; Zhang, S.; Pfeifer, B.A. Construction and performance of heterologous polyketide-producing K-12- and B-derived Escherichia coli. Lett. Appl. Microbiol. 2010, 51, 196–204. [Google Scholar] [CrossRef]
- Yang, D.; Park, S.Y.; Park, Y.S.; Eun, H.; Lee, S.Y. Metabolic Engineering of Escherichia coli for Natural Product Biosynthesis. Trends Biotechnol. 2020, 38, 745–765. [Google Scholar] [CrossRef]
- Huo, L.; Hug, J.J.; Fu, C.; Bian, X.; Zhang, Y.; Müller, R. Heterologous expression of bacterial natural product biosynthetic pathways. Nat. Prod. Rep. 2019, 41, 425–431. [Google Scholar] [CrossRef]
- Cummings, M.; Peters, A.D.; Whitehead, G.F.S.; Menon, B.R.K.; Micklefield, J.; Webb, S.J.; Takano, E. Assembling a plug-and-play production line for combinatorial biosynthesis of aromatic polyketides in Escherichia coli. PLoS Biol. 2019, 17, e3000347. [Google Scholar] [CrossRef]
- Liu, X.; Hua, K.; Liu, D.; Wu, Z.L.; Wang, Y.; Zhang, H.; Deng, Z.; Pfeifer, B.A.; Jiang, M. Heterologous Biosynthesis of Type II Polyketide Products Using E. coli. ACS Chem. Biol. 2020, 15, 1177–1183. [Google Scholar] [CrossRef]
- Kallscheuer, N.; Kage, H.; Milke, L.; Nett, M.; Marienhagen, J. Microbial synthesis of the type I polyketide 6-methylsalicylate with Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2019, 103, 9619–9631. [Google Scholar] [CrossRef]
- Mora-Lugo, R.; Stegmüller, J.; MacK, M. Metabolic engineering of roseoflavin-overproducing microorganisms. Microbial Cell Factories 2019, 18, 146. [Google Scholar] [CrossRef]
- Molin, G. Effect of carbon dioxide on growth of Pseudomonas putida ATCC 11172 on asparagine, citrate, glucose, and lactate in batch and continuous culture. Can. J. Microbiol. 1985, 31, 763–766. [Google Scholar] [CrossRef]
- Gross, F.; Gottschalk, D.; Müller, R. Posttranslational modification of myxobacterial carrier protein domains in Pseudomonas sp. by an intrinsic phosphopantetheinyl transferase. Appl. Microbiol. Biotechnol. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Loeschcke, A.; Thies, S. Pseudomonas putida—A versatile host for the production of natural products. Appl. Microbiol. Biotechnol. 2015, 99, 6197–6214. [Google Scholar] [CrossRef]
- Choi, K.R.; Cho, J.S.; Cho, I.J.; Park, D.; Lee, S.Y. Markerless gene knockout and integration to express heterologous biosynthetic gene clusters in Pseudomonas putida. Metab. Eng. 2018, 47, 463–474. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Pfeifer, B.A. Bacterial hosts for natural product production. Mol. Pharm. 2008, 5, 212–225. [Google Scholar] [CrossRef]
- Incha, M.R.; Thompson, M.G.; Blake-Hedges, J.M.; Liu, Y.; Pearson, A.N.; Schmidt, M.; Gin, J.W.; Petzold, C.J.; Deutschbauer, A.M.; Keasling, J.D. Leveraging host metabolism for bisdemethoxycurcumin production in Pseudomonas Putida. Metab. Eng. Commun. 2020, 10, e00119. [Google Scholar] [CrossRef]
- Kasuga, K.; Sasaki, A.; Matsuo, T.; Yamamoto, C.; Minato, Y.; Kuwahara, N.; Fujii, C.; Kobayashi, M.; Agematu, H.; Tamura, T.; et al. Heterologous production of kasugamycin, an aminoglycoside antibiotic from Streptomyces kasugaensis, in Streptomyces lividans and Rhodococcus erythropolis L-88 by constitutive expression of the biosynthetic gene cluster. Appl. Microbiol. Biotechnol. 2017, 101, 4259–4268. [Google Scholar] [CrossRef]
- Gomez-Escribano, J.P.; Castro, J.F.; Razmilic, V.; Jarmusch, S.A.; Saalbach, G.; Ebel, R.; Jaspars, M.; Andrews, B.; Asenjo, J.A.; Bibb, M.J. Heterologous Expression of a Cryptic Gene Cluster from Streptomyces leeuwenhoekii C34T Yields a Novel Lasso Peptide, Leepeptin. Appl. Environ. Microbiol. 2019, 85, e01752-19. [Google Scholar] [CrossRef]
- Tu, J.; Li, S.; Chen, J.; Song, Y.; Fu, S.; Ju, J.; Li, Q. Characterization and heterologous expression of the neoabyssomicin/abyssomicin biosynthetic gene cluster from Streptomyces koyangensis SCSIO 5802. Microb. Cell Factories 2018, 17, 28. [Google Scholar] [CrossRef]
- Bilyk, O.; Sekurova, O.N.; Zotchev, S.B.; Luzhetskyy, A. Cloning and heterologous expression of the grecocycline biosynthetic gene cluster. PLoS ONE 2016, 11, e0158682. [Google Scholar] [CrossRef]
- Greunke, C.; Duell, E.R.; D’Agostino, P.M.; Glöckle, A.; Lamm, K.; Gulder, T.A.M. Direct Pathway Cloning (DiPaC) to unlock natural product biosynthetic potential. Metab. Eng. 2018, 47, 334–345. [Google Scholar] [CrossRef]
- Li, J.; Jaitzig, J.; Theuer, L.; Legala, O.E.; Süssmuth, R.D.; Neubauer, P. Type II thioesterase improves heterologous biosynthesis of valinomycin in Escherichia coli. J. Biotechnol. 2015, 193, 16–22. [Google Scholar] [CrossRef]
- Hug, J.J.; Dastbaz, J.; Adam, S.; Revermann, O.; Koehnke, J.; Krug, D.; Müller, R. Biosynthesis of Cittilins, Unusual Ribosomally Synthesized and Post-translationally Modified Peptides from Myxococcus xanthus. ACS Chem. Biol. 2020, 15, 2221–2231. [Google Scholar] [CrossRef]
- Nara, A.; Hashimoto, T.; Komatsu, M.; Nishiyama, M.; Kuzuyama, T.; Ikeda, H. Characterization of bafilomycin biosynthesis in Kitasatospora setae KM-6054 and comparative analysis of gene clusters in Actinomycetales microorganisms. J. Antibiot. 2017, 70, 616–624. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkränzer, B.; Nadmid, S.; Pujic, P.; Normand, P.; Luzhetskyy, A. Generation of a cluster-free Streptomyces albus chassis strains for improved heterologous expression of secondary metabolite clusters. Metab. Eng. 2018, 49, 316–324. [Google Scholar] [CrossRef]
- Wang, X.; Yin, S.; Bai, J.; Liu, Y.; Fan, K.; Wang, H.; Yuan, F.; Zhao, B.; Li, Z.; Wang, W. Heterologous production of chlortetracycline in an industrial grade Streptomyces rimosus host. Appl. Microbiol. Biotechnol. 2019, 103, 6645–6655. [Google Scholar] [CrossRef]
- Xu, Z.; Li, Y. A MarR-family transcriptional factor MapR positively regulates actinorhodin production in Streptomyces coelicolor. FEMS Microbiol. Lett. 2020, 367, fnaa140. [Google Scholar] [CrossRef]
- Herrmann, S.; Siegl, T.; Luzhetska, M.; Jilg, L.P.; Welle, E.; Erb, A.; Leadlay, P.F.; Bechthold, A.; Luzhetskyy, A. Site-specific recombination strategies for engineering Actinomycete genomes. Appl. Environ. Microbiol. 2012, 78, 1804–1812. [Google Scholar] [CrossRef]
- Bu, Q.T.; Yu, P.; Wang, J.; Li, Z.Y.; Chen, X.A.; Mao, X.M.; Li, Y.Q. Rational construction of genome-reduced and high-efficient industrial Streptomyces chassis based on multiple comparative genomic approaches. Microb. Cell Factories 2019, 18, 16. [Google Scholar] [CrossRef]
- Lu, Z.; Xie, P.; Qin, Z. Promotion of markerless deletion of the actinorhodin biosynthetic gene cluster in Streptomyces coelicolor. Acta Biochim. Biophys. Sin. 2010, 42, 717–721. [Google Scholar] [CrossRef][Green Version]
- Fazal, A.; Thankachan, D.; Harris, E.; Seipke, R.F. A chromatogram-simplified Streptomyces albus host for heterologous production of natural products. Antonie Van Leeuwenhoek 2020, 113, 511–520. [Google Scholar] [CrossRef]
- Gust, B.; Challis, G.L.; Fowler, K.; Kieser, T.; Chater, K.F. PCR-targeted Streptomyces gene replacement identifies a protein domain needed for biosynthesis of the sesquiterpene soil odor geosmin. Proc. Natl. Acad. Sci. USA 2003, 100, 1541–1546. [Google Scholar] [CrossRef]
- Komatsu, M.; Uchiyama, T.; Omura, S.; Cane, D.E.; Ikeda, H. Genome-minimized Streptomyces host for the heterologous expression of secondary metabolism. Proc. Natl. Acad. Sci. USA 2010, 107, 2646–2651. [Google Scholar] [CrossRef]
- Fernández-Martínez, L.T.; Bibb, M.J. Use of the meganuclease I-SceI of Saccharomyces cerevisiae to select for gene deletions in Actinomycetes. Sci. Rep. 2014, 4, 7100. [Google Scholar] [CrossRef]
- Tong, Y.; Weber, T.; Lee, S.Y. CRISPR/Cas-based genome engineering in natural product discovery. Nat. Prod. Rep. 2019, 36, 1262–1280. [Google Scholar] [CrossRef]
- Huang, H.; Zheng, G.; Jiang, W.; Hu, H.; Lu, Y. One-step high-efficiency CRISPR/Cas9-mediated genome editing in Streptomyces. Acta Biochim. Biophys. Sin. 2015, 47, 231–243. [Google Scholar] [CrossRef]
- Cobb, R.E.; Wang, Y.; Zhao, H. High-Efficiency Multiplex Genome Editing of Streptomyces Species Using an Engineered CRISPR/Cas System. ACS Synth. Biol. 2015, 4, 723–728. [Google Scholar] [CrossRef]
- Salem, S.M.; Weidenbach, S.; Rohr, J. Two Cooperative Glycosyltransferases Are Responsible for the Sugar Diversity of Saquayamycins Isolated from Streptomyces sp. KY 40-1. ACS Chem. Biol. 2017, 12, 2529–2534. [Google Scholar] [CrossRef]
- Yeo, W.L.; Heng, E.; Tan, L.L.; Lim, Y.W.; Lim, Y.H.; Hoon, S.; Zhao, H.; Zhang, M.M.; Wong, F.T. Characterization of Cas proteins for CRISPR-Cas editing in Streptomycetes. Biotechnol. Bioeng. 2019, 116, 2330–2338. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, D.; Zhu, J.; Liu, H.; Liang, S.; Luo, Y. Efficient Multiplex Genome Editing in Streptomyces via Engineered CRISPR-Cas12a Systems. Front. Bioeng. Biotechnol. 2020, 8, 726. [Google Scholar] [CrossRef]
- Sarokin, L.; Carlson, M. Optimization of gene expression in Streptomyces lividans by a transcription terminator. Nucleic Acids Res. 1987, 15, 4227–4240. [Google Scholar]
- Horbal, L.; Siegl, T.; Luzhetskyy, A. A set of synthetic versatile genetic control elements for the efficient expression of genes in Actinobacteria. Sci. Rep. 2018, 8, 491. [Google Scholar] [CrossRef]
- Curran, K.A.; Karim, A.S.; Gupta, A.; Alper, H.S. Use of High Capacity Terminators in Saccharomyces cerevisiae to Increase mRNA half-life and Improve Gene Expression Control for Metabolic Engineering Applications. Metab. Eng. 2013, 19, 88–97. [Google Scholar] [CrossRef]
- Pulido, D.; Jiménez, A.; Salas, M.; Mellado, R.P. A Bacillus subtilis phage φ29 transcription terminator is efficiently recognized in Streptomyces lividans. Gene 1987, 56, 277–282. [Google Scholar] [CrossRef]
- Garst, A.D.; Edwards, A.L.; Batey, R.T. Riboswitches: Structures and mechanisms. Cold Spring Harb. Perspect. Biol. 2011, 3, a003533. [Google Scholar] [CrossRef]
- Rudolph, M.M.; Vockenhuber, M.P.; Suess, B. Synthetic riboswitches for the conditional control of gene expression in Streptomyces coelicolor. Microbiology 2013, 159, 1416–1422. [Google Scholar] [CrossRef]
- Eyles, T.H.; Vior, N.M.; Truman, A.W. Rapid and Robust Yeast-Mediated Pathway Refactoring Generates Multiple New Bottromycin-Related Metabolites. ACS Synth. Biol. 2018, 7, 1211–1218. [Google Scholar] [CrossRef]
- Horbal, L.; Luzhetskyy, A. Dual control system—A novel scaffolding architecture of an inducible regulatory device for the precise regulation of gene expression. Metab. Eng. 2016, 37, 11–23. [Google Scholar] [CrossRef]
- Gondry, M.; Sauguet, L.; Belin, P.; Thai, R.; Amouroux, R.; Tellier, C.; Tuphile, K.; Jacquet, M.; Braud, S.; Courçon, M.; et al. Cyclodipeptide synthases are a family of tRNA-dependent peptide bond-forming enzymes. Nat. Chem. Biol. 2009, 5, 414–420. [Google Scholar] [CrossRef]
- Meng, S.; Han, W.; Zhao, J.; Jian, X.H.; Pan, H.X.; Tang, G.L. A Six-Oxidase Cascade for Tandem C−H Bond Activation Revealed by Reconstitution of Bicyclomycin Biosynthesis. Angew. Chem. Int. Ed. 2018, 57, 719–723. [Google Scholar] [CrossRef]
- Liu, J.; Kaganjo, J.; Zhang, W.; Zeilstra-Ryalls, J. Investigating the bifunctionality of cyclizing and “classical” 5-aminolevulinate synthases. Protein Sci. 2018, 27, 402–410. [Google Scholar] [CrossRef]
- Zhu, L.; Qian, X.; Chen, D.; Ge, M. Role of two 5-aminolevulinic acid biosynthetic pathways in heme and secondary metabolite biosynthesis in Amycolatopsis orientalis. J. Basic Microbiol. 2018, 58, 198–205. [Google Scholar] [CrossRef]
- Leskiw, B.K.; Lawlor, E.J.; Fernandez-Abalos, J.M.; Chater, K.F. TTA codons in some genes prevent their expression in a class of developmental, antibiotic-negative, Streptomyces mutants. Proc. Natl. Acad. Sci. USA 1991, 88, 2461–2465. [Google Scholar] [CrossRef]
- Tercero, J.A.; Espinosa, J.C.; Jiménez, A. Expression of the Streptomyces alboniger pur cluster in Streptomyces lividans is dependent on the bldA-encoded tRNALeu. FEBS Lett. 1998, 421, 221–223. [Google Scholar] [CrossRef]
- Garg, R.P.; Gonzalez, J.M.; Parry, R.J. Biochemical characterization of VlmL, a seryl-tRNA synthetase encoded by the valanimycin biosynthetic gene cluster. J. Biol. Chem. 2006, 281, 26785–26791. [Google Scholar] [CrossRef]
- Jiang, M.X.; Yin, M.; Wu, S.H.; Han, X.L.; Ji, K.Y.; Wen, M.L.; Lu, T. GdmRIII, a TetR Family Transcriptional Regulator, Controls Geldanamycin and Elaiophylin Biosynthesis in Streptomyces autolyticus CGMCC0516. Sci. Rep. 2017, 7, 4803. [Google Scholar] [CrossRef]
- Lyu, M.; Cheng, Y.; Han, X.; Wen, Y.; Song, Y.; Li, J.; Chen, Z. AccR, a TetR family transcriptional repressor, coordinates short-chain acyl coenzyme A homeostasis in Streptomyces avermitilis. Appl. Environ. Microbiol. 2020, 86, e00508–e00520. [Google Scholar] [CrossRef]
- Shen, J.; Kong, L.; Li, Y.; Zheng, X.; Wang, Q.; Yang, W.; Deng, Z.; You, D. A LuxR family transcriptional regulator AniF promotes the production of anisomycin and its derivatives in Streptomyces hygrospinosus var. beijingensis. Synth. Syst. Biotechnol. 2019, 4, 40–48. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.; Lu, X.; Liu, W.; Chen, Z.; Li, J.; Deng, L.; Wen, Y. SAV4189, a MarR-family regulator in Streptomyces avermitilis, activates avermectin biosynthesis. Front. Microbiol. 2018, 9, 1358. [Google Scholar] [CrossRef]
- Peng, Q.; Gao, G.; Lü, J.; Long, Q.; Chen, X.; Zhang, F.; Xu, M.; Liu, K.; Wang, Y.; Deng, Z.; et al. Engineered Streptomyces lividans strains for optimal identification and expression of cryptic biosynthetic gene clusters. Front. Microbiol. 2018, 9, 3042. [Google Scholar] [CrossRef]
- Ish-Horowicz, D.; Burke, J.F. Rapid and efficient cosmid cloning. Nucleic Acids Res. 1981, 9, 2989–2998. [Google Scholar] [CrossRef]
- Saraswathy, N.; Ramalingam, P. High capacity vectors. Concepts Tech. Genom. Proteom. 2011, 49–56. [Google Scholar] [CrossRef]
- Zimmer, R.; Gibbins, A.M.V. Construction and Characterization of a Large-Fragment Chicken Bacterial Artificial Chromosome Library. Genomics 1997, 226, 217–226. [Google Scholar] [CrossRef]
- Jones, A.C.; Gust, B.; Kulik, A.; Heide, L.; Buttner, M.J.; Bibb, M.J. Phage P1-Derived Artificial Chromosomes Facilitate Heterologous Expression of the FK506 Gene Cluster. PLoS ONE 2013, 8, e69319. [Google Scholar] [CrossRef]
- Kouprina, N.; Larionov, V. TAR cloning: Insights into gene function, long-range haplotypes and genome structure and evolution. Nat. Rev. Genet. 2006, 7, 805–812. [Google Scholar] [CrossRef]
- Yamanaka, K.; Reynolds, K.A.; Kersten, R.D.; Ryan, K.S.; Gonzalez, D.J.; Nizet, V.; Dorrestein, P.C.; Moore, B.S. Direct cloning and refactoring of a silent lipopeptide biosynthetic gene cluster yields the antibiotic taromycin A. Proc. Natl. Acad. Sci. USA 2014, 111, 1957–1962. [Google Scholar] [CrossRef]
- Fu, J.; Bian, X.; Hu, S.; Wang, H.; Huang, F.; Seibert, P.M.; Plaza, A.; Xia, L.; Müller, R.; Stewart, A.F.; et al. Full-length RecE enhances linear-linear homologous recombination and facilitates direct cloning for bioprospecting. Nat. Biotechnol. 2012, 30, 440–446. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.; Jia, R.; Yin, J.; Li, A.; Xia, L.; Yin, Y.; Müller, R.; Fu, J.; Stewart, A.F.; et al. ExoCET: Exonuclease in vitro assembly combined with RecET recombination for highly efficient direct DNA cloning from complex genomes. Nucleic Acids Res. 2018, 46, e28. [Google Scholar] [CrossRef]
- Du, D.; Wang, L.; Tian, Y.; Liu, H.; Tan, H.; Niu, G. Genome engineering and direct cloning of antibiotic gene clusters via phage φBT1 integrase-mediated site-specific recombination in Streptomyces. Sci. Rep. 2015, 5, 8740. [Google Scholar] [CrossRef]
- Vior, N.M.; Lacret, R.; Chandra, G.; Dorai-Raj, S.; Trick, M.; Truman, A.W. Discovery and biosynthesis of the antibiotic bicyclomycin in distantly related bacterial classes. Appl. Environ. Microbiol. 2018, 84, e02828-17. [Google Scholar] [CrossRef]
- Mevaere, J.; Goulard, C.; Schneider, O.; Sekurova, O.N.; Ma, H.; Zirah, S.; Afonso, C.; Rebuffat, S.; Zotchev, S.B.; Li, Y. An orthogonal system for heterologous expression of actinobacterial lasso peptides in Streptomyces hosts. Sci. Rep. 2018, 8, 8232. [Google Scholar] [CrossRef]
- Duell, E.R.; D’Agostino, P.M.; Shapiro, N.; Woyke, T.; Fuchs, T.M.; Gulder, T.A.M. Direct pathway cloning of the sodorifen biosynthetic gene cluster and recombinant generation of its product in E. coli. Microb. Cell Factories 2019, 18, 32. [Google Scholar] [CrossRef]
- D’Agostino, P.M.; Gulder, T.A.M. Direct Pathway Cloning Combined with Sequence- and Ligation-Independent Cloning for Fast Biosynthetic Gene Cluster Refactoring and Heterologous Expression. ACS Synth. Biol. 2018, 7, 1702–1708. [Google Scholar] [CrossRef]
- Colloms, S.D.; Merrick, C.A.; Olorunniji, F.J.; Stark, W.M.; Smith, M.C.M.; Osbourn, A.; Keasling, J.D.; Rosser, S.J. Rapid metabolic pathway assembly and modification using serine integrase site-specific recombination. Nucleic Acids Res. 2014, 42, e23. [Google Scholar] [CrossRef]
- Gao, H.; Taylor, G.; Evans, S.K.; Fogg, P.C.M.; Smith, M.C.M. Application of serine integrases for secondary metabolite pathway assembly in Streptomyces. Synth. Syst. Biotechnol. 2020, 5, 111–119. [Google Scholar] [CrossRef]
- Wang, X.; Tang, B.; Ye, Y.; Mao, Y.; Lei, X.; Zhao, G.; Ding, X. Bxb1 integrase serves as a highly efficient DNA recombinase in rapid metabolite pathway assembly. Acta Biochim. Biophys. Sin. 2017, 49, 44–50. [Google Scholar] [CrossRef]
- Enghiad, B.; Zhao, H. Programmable DNA-Guided Artificial Restriction Enzymes. ACS Synth. Biol. 2017, 6, 752–757. [Google Scholar] [CrossRef]
- Basitta, P.; Westrich, L.; Rösch, M.; Kulik, A.; Gust, B.; Apel, A.K. AGOS: A Plug-and-Play Method for the Assembly of Artificial Gene Operons into Functional Biosynthetic Gene Clusters. ACS Synth. Biol. 2017, 6, 817–825. [Google Scholar] [CrossRef]
- Jiang, W.; Zhao, X.; Gabrieli, T.; Lou, C.; Ebenstein, Y.; Zhu, T.F. Cas9-Assisted Targeting of CHromosome segments CATCH enables one-step targeted cloning of large gene clusters. Nat. Commun. 2015, 6, 8101. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, N.; Jeong, Y.; Hwang, S.; Kim, W.; Cho, S.; Palsson, B.O.; Cho, B.K. The Transcription Unit Architecture of Streptomyces lividans TK24. Front. Microbiol. 2019, 10, 2074. [Google Scholar] [CrossRef]
- Ji, C.H.; Kim, J.P.; Kang, H.S. Library of Synthetic Streptomyces Regulatory Sequences for Use in Promoter Engineering of Natural Product Biosynthetic Gene Clusters. ACS Synth. Biol. 2018, 7, 1946–1955. [Google Scholar] [CrossRef]
- Šmídová, K.; Ziková, A.; Pospíšil, J.; Schwarz, M.; Bobek, J.; Vohradsky, J. DNA mapping and kinetic modeling of the HrdB regulon in Streptomyces coelicolor. Nucleic Acids Res. 2019, 47, 621–633. [Google Scholar] [CrossRef]
- Hwang, S.; Lee, N.; Jeong, Y.; Lee, Y.; Kim, W.; Cho, S.; Palsson, B.O.; Cho, B.K. Primary transcriptome and translatome analysis determines transcriptional and translational regulatory elements encoded in the Streptomyces clavuligerus genome. Nucleic Acids Res. 2019, 47, 6114–6129. [Google Scholar] [CrossRef]
- Zhou, Q.; Ning, S.; Luo, Y. Coordinated regulation for nature products discovery and overproduction in Streptomyces. Synth. Syst. Biotechnol. 2020, 5, 49–58. [Google Scholar] [CrossRef]
- Takano, E.; White, J.; Thompson, C.J.; Bibb, M.J. Construction of thiostrepton-inducible, high-copy-number expression vectors for use in Streptomyces spp. Gene 1995, 166, 133–137. [Google Scholar] [CrossRef]
- Herai, S.; Hashimoto, Y.; Higashibata, H.; Maseda, H.; Ikeda, H.; Omura, S.; Kobayashi, M. Hyper-inducible expression system for Streptomycetes. Proc. Natl. Acad. Sci. USA 2004, 101, 14031–14035. [Google Scholar] [CrossRef]
- Bibb, M.J.; Janssen, G.R.; Ward, J.M. Cloning and analysis of the promoter region of the erythromycin-resistance gene (ermE) of Streptomyces erythraeus. Gene 1985, 38, 215–226. [Google Scholar] [CrossRef]
- Labes, G.; Bibb, M.; Wohlleben, W. Isolation and characterization of a strong promoter element from the Streptomyces ghanaensis phage 119 using the gentamicin resistance gene (aacC1) of Tn1696 as reporter. Microbiology 1997, 143, 1503–1512. [Google Scholar] [CrossRef]
- Wang, W.; Li, X.; Wang, J.; Xiang, S.; Feng, X.; Yang, K. An engineered strong promoter for Streptomycetes. Appl. Environ. Microbiol. 2013, 79, 4484–4492. [Google Scholar] [CrossRef]
- Wang, K.; Liu, X.F.; Bu, Q.T.; Zheng, Y.; Chen, X.A.; Li, Y.Q.; Mao, X.M. Transcriptome-Based Identification of a Strong Promoter for Hyper-production of Natamycin in Streptomyces. Curr. Microbiol. 2019, 76, 95–99. [Google Scholar] [CrossRef]
- Farasat, I.; Kushwaha, M.; Collens, J.; Easterbrook, M.; Guido, M.; Salis, H.M. Efficient search, mapping, and optimization of multi-protein genetic systems in diverse bacteria. Mol. Syst. Biol. 2014, 10, 731. [Google Scholar] [CrossRef]
- Na, D.; Lee, D. RBSDesigner: Software for designing synthetic ribosome binding sites that yields a desired level of protein expression. Bioinformatics 2010, 26, 2633–2634. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Native Strain | Heterologous Host | Method Clone | Products |
|---|---|---|---|
| Streptomyces leeuwenhoekii C34T | S. coelicolor M1152 and M1154 | PCR, cloning to pIJ10257 | Leepeptin [95] |
| S. variabilis Snt24 | S. lividans SBT5 | BAC clone | Tetramic acid [21] |
| Streptomyces koyangensis SCSIO 5802 | S. coelicolor M1152 | Phage-P1-derived artificial chromosome (PAC) library | Neoabyssomicin, abyssomicin [96] |
| Streptomyces sp. Acta1362 | S. albus J1074 | TAR system | Grecocycline [97] |
| S. venezuelae | S. lividans and S. coelicolor | BAC cloning | Pikromycin [22] |
| S. galtieri Sgt26 | S. albus J1074 | BAC cloning | Kinamycin [58] |
| S. albus subsp. chlorinus NRRL B-24108 | S. albus Del14, S. lividans TK24 | BAC clone | Albucidin [57] |
| S. erythraea DSM 40517 | S. coelicolor M1152 and M1154 | Direct pathway cloning (DiPaC) | Erythromycin [98] |
| Streptomyces tsusimaensis ATCC 15141 | E. coli | Co-expression genes | Valinomycin [99] |
| S. erythraea | B. subtilis | PCR, cloning | 6-deoxyerythronolide B [76] |
| S. davaonensis | C. glutamicum | PCR, cloning | Roseoflavin [87] |
| S. griseus | P. putida | PCR, cloning | Flaviolin [91] |
| S. kasugaensis | R. erythropolis | PCR, cloning | Kasugamycin [94] |
| Actinosynnema mirum DSM 43827 | S. avermitilis SUK22 | PCR, cloning | Mycosporine-glycine-alanine [67] |
| Kocuria rosea s17 | S. coelicolor M1146, S. sp. s120 | Gibson assembly, integration vector pSET152 | Kocurin [23] |
| Amycolatopsis sp. DEM30355 | S. coelicolor M1152 | PAC library | Vancoresmycin [26] |
| Myxococcus xanthus DK1622 | S. albus Del14 | PCR and subcloned | Cittilins [100] |
| Actinoplanes tsinanensis CPCC 200056 | S. coelicolor M1146 | DNA assembler by yeast | Chuangxinmycin [60] |
| Kutzneria albida DSM 43870 | S. albus Del14 | BAC vector | Huimycin [62] |
| Planobisporarosea ATCC 53733 | S.coelicolor M1146 | Cosmid vector | GE2270 [66] |
| A. mirum DSM 43827 | S. avermitilis SUKA22 | In vivo by phage λ-Red recombining system | Shinorine, porphyra-334, mycosporine-glycine, and mycosporine-glycine-alanine [67] |
| Kitasatospora setae KM-6054 | Streptomyces lohii JCM 14114, S. griseus DSM 2608 | BAC cloning | Bafilomycins A1, C1, and B1 (setamycin) [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, V.T.T.; Nguyen, C.T.; Dhakal, D.; Nguyen, H.T.; Kim, T.-S.; Sohng, J.K. Recent Advances in the Heterologous Biosynthesis of Natural Products from Streptomyces. Appl. Sci. 2021, 11, 1851. https://doi.org/10.3390/app11041851
Pham VTT, Nguyen CT, Dhakal D, Nguyen HT, Kim T-S, Sohng JK. Recent Advances in the Heterologous Biosynthesis of Natural Products from Streptomyces. Applied Sciences. 2021; 11(4):1851. https://doi.org/10.3390/app11041851
Chicago/Turabian StylePham, Van Thuy Thi, Chung Thanh Nguyen, Dipesh Dhakal, Hue Thi Nguyen, Tae-Su Kim, and Jae Kyung Sohng. 2021. "Recent Advances in the Heterologous Biosynthesis of Natural Products from Streptomyces" Applied Sciences 11, no. 4: 1851. https://doi.org/10.3390/app11041851
APA StylePham, V. T. T., Nguyen, C. T., Dhakal, D., Nguyen, H. T., Kim, T.-S., & Sohng, J. K. (2021). Recent Advances in the Heterologous Biosynthesis of Natural Products from Streptomyces. Applied Sciences, 11(4), 1851. https://doi.org/10.3390/app11041851