Featured Application
In these times, in accordance with the European Green Deal and the “Farm to Fork Strategy” there should be a strong emphasis on efforts to create a healthier and more sustainable food system. One of the key targets in this plan is the reduction by 2030 of the overall use of chemical pesticides by 50%, which prompts scientists to intensify research in the search for alternative solutions. Healthy food means safe food, free from pathogens and toxic metabolites, such as Fusarium fungi and their mycotoxins. Essential oils are a noteworthy alternative, since more and more studies have proven their strong antifungal and antimycotoxigenic activity. The presented research broadens and complements the existing knowledge in the field of practical application of oils in biological control of plant pathogens.
Abstract
The presence of Fusarium fungi and their toxic metabolites in agricultural crops contributes to significant quantitative and qualitative losses of crops, causing a direct threat to human and animal health and life. Modern strategies for reducing the level of fungi and mycotoxins in the food chain tend to rely on natural methods, including plant substances. Essential oils (EOs), due to their complex chemical composition, show high biological activity, including fungistatic properties, which means that they exhibit high potential as a biological plant protection factor. The aim of this study was to determine the fungistatic activity of three EOs against F. graminearum, and the reduction of mycotoxin biosynthesis in corn and wheat grain. All tested EOs effectively suppressed the growth of F. graminearum in concentrations of 5% and 10%. Cinnamon and verbena EOs also effectively reduced the ergosterol (ERG) content in both grains at the concentration of 1%, while at the 0.1% EO concentration, the reduction in the ERG amount depended on the EO type as well as on the grain. The degree of zearalenone (ZEA) reduction was consistent with the inhibition of ERG biosynthesis, while the reduction in deoxynivalenol (DON) was not consistent with this parameter.
1. Introduction
Wheat (Triticum aestivum L.) and maize (Zea mays L.) are the most important cereal crops worldwide, with over 775.8 million and 1125 billion metric tons produced in 2020/21, respectively [1]. Both cereals are used as food ingredients and animal feed, as they are good sources of nutrients. Moreover, different types of food and industrial products, including sweeteners, starch, oil, beverages, glue, industrial alcohol, and fuel ethanol, can be produced from wheat and maize. The consumption of these cereals varies mainly by region; maize is consumed highly in Central America, Southern and Eastern Africa, and Mexico, while wheat is preferred mainly in Central Asia, the Middle East, South and North America, and Europe [2]. A serious problem in wheat and corn crops consists of infections caused by the Fusarium genus, the filamentous fungi commonly found in soil and on plants, contaminating agricultural commodities both on the field and in storage [3]. Fusarium graminearum is one of the most prevalent species that infect cereals in humid and semi-humid areas worldwide [4], responsible for devastating and hazardous plant diseases, such as stalk and ear rot of maize, as well as Fusarium head blight (FHB), crown rot, and seedling blight on wheat and other cereals [5]. Fusarium diseases cause losses not only due to yield, quality and nutritional properties reduction, but also because contamination of grains with mycotoxins affects food and feed safety, posing a threat to human and animals’ health [4]. The most widespread mycotoxins associated with F. graminearum infection in cereals include zearalenone (ZEA) and trichothecenes, such as nivalenol (NIV), deoxynivalenol (DON), and its acetylated derivatives 3-acetyl-deoxynivalenol (3-ADON), 15-acetyl-deoxynivalenol (15-ADON) and deoxynivalenol 3 β–D-glucoside (DON-3-G) [6,7,8]. ZEA is considered an estrogenic mycotoxin and found to be toxic for the liver, kidney, and immune system [9], while trichothecenes can inhibit protein synthesis and induce anorexia, diarrhea, vomiting, and cell death, along with altering immune function, phosphokinase-mediated stress pathways, proinflammatory gene expression, gastrointestinal function, and the action of growth hormones [6,10].
Up to now, synthetic fungicides have been the primary tool to control contamination by Fusarium fungi and their mycotoxins, while many other policies have been developed and implemented to inhibit or reduce fungal growth and mycotoxin biosynthesis in different crops, notably in cereals. In recent years, the use of chemicals has increased consumer awareness, and their use is becoming more challenging due to carcinogenic effects, residual toxicity problems, environmental pollution, and increasing microbial resistance [11]. Therefore, finding alternatives to chemical fungicides that are considered safe, with negligible risk to human health and the environment, has become an exciting topic among researchers. EOs, which are naturally antimicrobial and antifungal substances, have received much attention and are considered as the biological antifungal agents that can replace synthetic pesticides [4,12]. Their significant benefit is that many EOs are on the GRAS (Generally Recognized as Safe) list approved by the FDA (Food and Drug Administration), therefore they can be stated as safe for the consumers and the environment [13].
EOs are natural secondary metabolites, consisting mainly of monoterpenes and sesquiterpenes [14]. EOs are accumulated in cells, secretory cavities, or glandular hairs of plants, and they can be derived from one or more plant parts, such as the root (e.g., vetiver grass, angelica, valerian), stems and leaves (e.g., verbena, petitgrain, patchouli, geranium), leaves (e.g., mint, lemongrass, lemon balm, citronella), buds (e.g., clove), flowers (e.g., lavender, rose, jasmine, carnation, clove, mimosa, rosemary), fruits (e.g., lemon, orange, juniper) or their peels (e.g., orange, lemon, grapefruit), bark (e.g., cinnamon), rhizomes (e.g., ginger, calamus, curcuma, orris), and seeds (e.g., fennel, coriander, caraway, dill, nutmeg, pepper) [14,15]. There are different methods for EO extraction, such as hydrodistillation, steam distillation, maceration, CO2 extraction, solvent extraction, enfleurage, and cold press extraction [15]. Furthermore, different studies have shown that the extraction method has an influence on the content of individual compounds, the yield of EOs, and their activities. Due to different constituents of EOs and their biological properties (antioxidant, antibacterial, antifungal), they have wide applications, among others, in cosmetics (e.g., fragrances and lotions), in food products (e.g., flavorings and preservatives), in agriculture (e.g., insecticides and repellents), and in pharmaceutical products (e.g., therapeutic action and sanitary activity) [16,17,18,19].
Studies conducted in recent years show that the application of EOs to control the pathogenic Fusarium spp. that attack cereals and cereal products is very effective. For instance, mycelial growth was significantly or completely inhibited in the tested isolates of Fusarium (F. oxysporum, F. solani, F. verticillioides, and F. subglutinans) by EOs of eucalyptus, clove, lemongrass, and mint [20,21]. In turn, the EOs of bay leaf, cinnamon, clove and oregano reduced fungal growth of F. culmorum and F. verticillioides by 90% to almost 100% [22]. In research conducted by Kalagatur et al. [23], the use of Cymbopogon martinii EO (from leaves of tropical herbaceous grass) in the encapsulated form with chitosan has significantly inhibited the growth of F. graminearum through the reduction of ergosterol (ERG) content and increase in intracellular reactive oxygen species and lipid peroxidation [23]. In a similar study, Ferreira and co-workers have reported the powerful effects of ginger EO against F. graminearum in vitro through the ERG and DON reduction [4]. The inhibitory effect of EOs showed dose-dependent activity on the tested Fusarium spp.
In the presented study, the antifungal and anti-mycotoxigenic effects of various concentrations of commercial palmarosa, cinnamon, and verbena EOs on the growth of F. graminearum and biosynthesis of mycotoxins in wheat and maize grain were analyzed. The aim of the present study was to determine an inhibitory dose that effectively inhibits both the growth of F. graminearum and mycotoxin biosynthesis. To our best knowledge, there are still limited data of complex EO analyses taking into account the relationship between the concentration and the effectiveness of their action. In relation to earlier research, where the main purpose was comparison of the antifungal and antimycotoxigenic activity of selected EOs against F. graminearum and F. culmorum in wheat and maize grains, a wider range of experimental concentrations was used, as well as extended scope of research including advanced microscopic analyses. It is also worth underlining that the issues raised in this work are especially important considering the potential application of EOs.
2. Materials and Methods
2.1. Plant Material
Wheat (cultivar Jantarka) and maize (cultivar Wiarus) grain for the research was obtained from the Research Centre for Registration of Agrochemicals, Institute of Plant Protection, National Research Institute, in Poznań, Poland. The chemical composition of wheat grains was as follows: protein 11.2% DM, starch 67.5% DM, gluten 18.7%, moisture after storage 11.5%, and thousand grain weight (TGW) 33.7 g, while the composition of maize grains was as follows: protein 12.5 % DM, starch 68.4 % DM, oil 4.6%, moisture after storage 13%, and thousand grain weight (TGW) 312.5 g. Analyses were conducted with the FOSS Infratec™ 1241 Grain Analyser. The samples (50 g) were mixed with 10 mL of deionized water and sterilized at 121 °C.
2.2. Fusarium Strain
F. graminearum, KZF-1 was obtained from the collection of the Research Centre for Registration of Agrochemicals, Institute of Plant Protection, National Research Institute, in Poznań, Poland. The tested strain was cultured before the experiment in Petri dishes (9 cm diameter) on a PDA medium (Potato Dextrose Agar, A&A Biotechnology) at 25 °C for 5–7 days.
2.3. Standards, Chemicals, and Reagents
ZEA, DON, and ERG analytical standards were purchased from Sigma-Aldrich (Steinheim, Germany). LC-MS acetonitrile, methanol, and water (MS grade) were purchased from J.T. Baker (Deventer, The Netherlands). Chemical reagents necessary for the extraction and purification process were obtained from Sigma-Aldrich and POCh (Gliwice, Poland). Individual stock solutions for all analytes were prepared by dissolving in acetonitrile.
2.4. EOs Preparations
Studies included selected EOs, such as cinnamon bark (Cinnamomum zeylanicum, Indonesia), and palmarosa leaves (Cymbopogon martini, India), as well as verbena leaves and flowers (Thymus hiemalis, Spain). Composition of tested EOs was as following: (i) cinnamon bark: cinnamic aldehyde ≤ 70%, eugenol ≤ 4.4%, linalool ≤ 2.6%, limonene ≤ 1.1%, benzyl benzoate ≤ 1.1%, benzaldehyde 0.5%, cinnamic alcohol ≤ 0.4%, and cuminaldehyde ≤ 0.2%; (ii) palmarosa: geraniol 85%, linalool 2–3%, limonene 1%, and citral 1%; (iii) verbena: citral 42%, and limonene 40%. Commercial EOs were purchased from Ecospa s.c., Poland. The studies included four different concentrations of EO deionized water solutions (10%, 5%, 1% and 0.1%) containing 10% Tween 80.
2.5. Antifungal Activity of EOs by Disc Volatilization Method
The antifungal activity of EOs was determined by the disc volatilization method according to [24,25], with some modifications. The Petri plates, containing 10 mL of the PDA medium, were inoculated with 6 mm diameter plugs of the actively growing F. graminearum mycelium from 5-day old cultures. Next, 6 mm diameter sterile paper discs (Oxoid, Basingstoke, Hants, United Kingdom) were placed on the inner surface of the Petri dishes lids (one on each) and soaked with 5 µL of four different concentrations of the EOs. A sample with a water-soaked blank paper disc was used as a control. The Petri plates were immediately closed, sealed with parafilm and incubated until the mycelium in the control reached the edge of the plate. After incubation, the diameter of the mycelium was measured, and the antifungal activity was expressed as the percentage inhibition of mycelial growth in relation to the control.
2.6. EOs and the Growth of Fusarium and Mycotoxins Biosynthesis
EO solution samples in the amount of 5 mL were mixed thoroughly with 50 g of sterilized wheat and maize grains (depending on the samples) in Erlenmeyer flasks, under sterile conditions. Three plugs (6 mm) of 5-day old fungal mycelium grown on PDA were added to the maize grains. For a given series of samples/replicates, the mycelium plugs were cut out from the same plate at an equal distance from the edges of the colony to ensure the same state of development of the fungal mycelium. Next, the samples were incubated in a dark room at 25 °C for 28 days. After incubation, the samples were dried, milled, homogenized, and prepared for chromatographic analysis. Control samples were grains with the addition of deionized water and Tween 80 (without the addition of EOs).
2.7. Chemical Analysis
2.7.1. Ergosterol
ERG is a typical component of fungal cell membranes that is not present in higher plants; therefore, it can be used as a natural, selective indicator for the presence of fungi in different matrices. Dried and milled samples of maize or wheat grain in the amount of 100 mg were suspended in 2 mL of methanol and 0.5 mL of a 2 M aqueous solution of sodium hydroxide. Next, samples in tubes were tightly screwed and microwaved three times (370 W power) for 10 s, and stored at room temperature for cooling down. Subsequently, samples were neutralized with 1 mL of 1 M hydrochloric acid solution, and 2 mL of methanol. After mixing, triple extraction by 4 mL of n-pentane was performed. Each time, the n-pentane layer was transferred to the vials and dried under a stream of nitrogen. The dry residue was dissolved in 1 mL of methanol and filtered through a 0.20 µm syringe filter (Chromafil, Macherey-Nagel, Duren, Germany) before HPLC/PDA analysis. ERG was detected on a HPLC Waters Alliance system with a Waters 2996 Photodiode Array Detector (Waters Division of Millipore, Milford, MA, USA) set at 282 nm and a 3.9 × 150 mm Nova Pak C-18 chromatographic column. The mobile phase was methanol:acetonitrile (90:10, v/v) at a flow rate of 1.0 mL/min. The concentration of ERG was determined as a result of the comparison of retention times with the external standard. The detection limit was 10 ng/g.
2.7.2. Mycotoxins Analysis
The extraction of mycotoxins was carried out by adding up to 5 g of ground samples to 20 mL of the extraction mixture (acetonitrile:water:acetic acid, 79:20:1, v/v/v), followed by vortexing (about 30 s) and mixing using a horizontal shaker for 24 h. After extraction, the samples were centrifuged at 3000 rpm for 10 min, and finally, all samples were filtered through a 0.20 µm syringe filter (Chromafil, Macherey-Nagel, Duren, Germany) before LC/MS/MS analyses.
The analytical system consisted of the Aquity UPLC chromatography (Waters, Manchester, MA, USA) coupled to an electrospray ionization triple quadrupole mass spectrometer (TQD) (Waters, Manchester, MA, USA). A Waters ACQUITY UPLC HSS T3 (100 × 2.1 mm/ID, with 1.8 µm particle size) (Waters, Manchester, MA, USA) was used for chromatographic separation with a flow rate 0.35 mL/min at room temperature. Gradient elution was applied using water buffered with 10 mM ammonium acetate (A) and acetonitrile (B). The solvent gradient was modified as follows: 0–2 min at 5% B and 2–7 min at 55% B. Next, these conditions were maintained for 2 min and 9–15 min at 90% B with isocratic elution for 2 min, followed by a return to the initial conditions. Nitrogen above 99% purity was used. The collision-induced decomposition was run using argon as the collision gas, with a collision energy of 14–22 eV. Multiple reaction monitoring (MRM) was used for quantitative analysis of the compounds. The analytes were identified by comparing the retention times and m/z values obtained by MS and MS2 with the mass spectra (317.1/174.9 and 297.3/249.1 for ZEA and DON, respectively) of the corresponding standards tested under the same conditions. All samples were injected in triplicate.
2.8. Morphology Observations
F. graminearum was cultured in PDB (Potato Dextrose Broth) for 5 days at 25 °C. Then cultures were centrifugated and treated with different concentrations of EOs for 1 h at 25 °C. Samples were transferred on glass slides and photographed with a light microscope (Olympus BX53, Olympus Corporation, Tokyo, Japan) at 400 × magnification. Fungal samples treated with sterile water were used as control samples. Each assay was repeated in triplicate.
2.9. Staining and Fluorescence Microscopy
The fluorescent probes, carboxyfluorescein diacetate (cFDA), and propidium iodide (PI) (Sigma Aldrich, Steinheim, Germany) were applied to assess the viability of F. graminearum. The procedure of staining was based on the method described by [26,27] with some modifications. A stock solution of cFDA (1 mg/mL) was prepared in dimethyl sulfoxide (DMSO), and a stock solution of PI (1 mg/mL) was prepared in distilled water and stored in the refrigerator. Centrifuged cultures of F. graminearum, prepared as described in Section 2.8, were suspended in 5 mL of 50 mM phosphate buffer with 25 μL cFDA and 25 μL PI at 30 °C for 15 min. Next, the staining solution was removed by centrifugation (8000 rpm, 1 min). The pellet was examined under the fluorescence microscope Olympus BX53 equipped with specific wavelength filters filter set (CFDA excitation/emission: 485/530; PI excitation/emission: 538/617).
2.10. Statistical Analysis
The results are presented as the mean (±standard deviation) of three parallel replicates. The effect of EOs on the reduction of ERG, ZEA, and DON was estimated by one-way analysis of variance (ANOVA). The homogeneity of variance was tested by Levene’s test and based on the results for homogeneous samples the Tukey’s test, and for nonhomogeneous samples, the Games–Howell test with a p-value < 0.05 was applied. Analyses were conducted using the IBM SPSS Statistics program.
3. Results
3.1. The Inhibition of the Growth of F. graminearum Depending on the EO Concentration
The EOs used in the presented work were selected on the basis of the earlier studies [28,29]. Four different concentrations of cinnamon, palmarosa and verbena EOs were used. The results of antifungal activity towards F. graminearum obtained by the disc volatilization method are presented in Table 1 and Figure 1. Cinnamon and verbena EOs demonstrated strong antifungal activity at concentrations of 10% and 5%, reaching complete growth inhibition in the highest used dose, while at a concentration of 5%, the strongest activity was shown by cinnamon EO. Palmarosa EO at the highest concentration showed only a weak impact, up to 5%, however no statistical differences compared to the control were observed. At the concentrations of 1% and 0.1%, none of the used EOs showed statistically significant fungistatic activity against F. graminearum.

Table 1.
Volatile effect of EOs on the mycelial growth of F. graminearum.
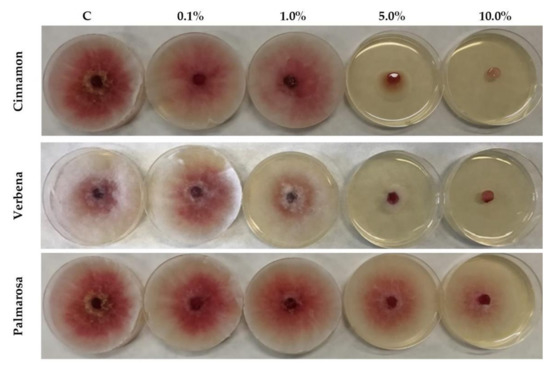
Figure 1.
Volatile effects of different concentrations of palmarosa, verbena and cinnamon EOs on the mycelial growth of F. graminearum on PDA medium (C–control; EO tested concentrations: 0.1%; 1.0%; 5.0%; 10.0%).
3.2. Staining and Microscopy
The microscopy observations revealed the differences in the morphological appearance of hyphae. Figure 2 presents the morphology of fungal cells after incubation with cinnamon EO. In the untreated sample, hyphae were gentle and rather thin, as well as having homogeneous cell structure (Figure 2a), while in samples treated with EOs, hyphae became thicker and the internal structure was heterogeneous (Figure 2b–e). With the increasing concentrations of EOs, the changes became more visible. While in hyphae treated with 0.1% and 1% EO solutions, the heterogeneity of the internal structure was noticeable in the hyphae fragments, fungal cells treated with higher EO concentrations were completely changed. Similar observations were made for the morphology of F. graminearum hyphae treated with verbena EO (Figure S1 in Supplementary Material). Under the light microscope (magnification × 400), changes in the morphology of F. graminearum hyphae) treated with palmarosa EO were not observed, due to exhibiting the weakest activity (up to 5% growth reduction for an EO concentration of 10%).

Figure 2.
Morphology of F. graminearum hyphae under light microscope (magnification × 400) treated with cinnamon EO: (a) Control; (b) EO concentration 0.1%; (c) EO concentration 1.0%; (d) EO concentration 5.0%; (e) EO concentration 10.0%.
The fluorescence method using cFDA and PI was employed to evaluate the viability of fungal cells. The results of cFDA/ PI staining of F. greminearum treated with cinnamon EO are shown in Figure 3. cFDA is a probe that can pass through the membranes of cells [30]. In living cells, it is hydrolysed by non-specific esterases to release the fluorescent carboxyfluorescein (green fluorescence). PI is a dye that can only pass through the membranes of damaged or dead cells due to the permeability of the plasma membrane and releases red fluorescence [31].
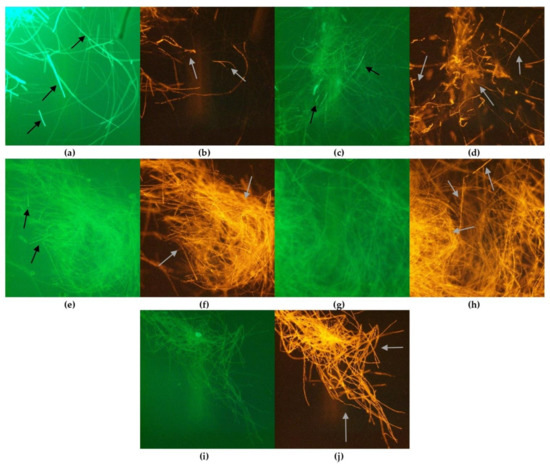
Figure 3.
Morphology of F. graminearum hyphae under optical microscope (magnification × 400): untreated: (a) control, stained with cFDA; (b) control, stained with PI; treated with cinnamon EO: stained with cFDA, EO concentration: (c) 0.1%; (e) 1.0%; (g) 5.0%; (i) 10.0%; stained with PI, EO concentration: (d) 0.1%; (f) 1.0%; (h) 5.0%; (j) 10.0%.
Great differences were observed between samples treated and untreated with tested EOs. Untreated, control hyphae (Figure 3a,b) show strong green fluorescence demonstrating high viability, with only some single cells stained in red. In the samples treated with EOs, the viability of fungal cells depended on the EO concentration, as shown in the example of cinnamon oil (Figure 3c–i). In samples with 0.1%, high viability of cells was still observed, as the green fluorescence was visible and very few cells demonstrated red fluorescence. However, a higher concentration of EO caused a drastic decrease in viable cells. The fungal cells observed under the microscope showed strong red fluorescence, while green fluorescence was found. Similar findings were observed for the morphology of F. graminearum treated with all tested EOs (Figures S2 and S3 in Supplementary Material).
3.3. The Effect of EO Concentration on the Growth of F. graminearum on Maize and Wheat Grain
The effect of EOs on the F. graminearum growth on maize and wheat grains was determined by the measurement of ERG concentration by HPLC analysis (Figure 4). The reduction percentage of ERG in grain samples treated with EO preparations was calculated in comparison to the control samples, where the grains were treated with a mixture of deionized water and Tween 80 without the addition of EO. The ERG was reduced significantly in the concentration range of 1% to 10% by all tested EOs, in both maize and wheat grain. Cinnamon and verbena EOs effectively reduced the ERG content in both wheat and maize grains at the concentration range of 1% to 10% (ERG level reduction in the range of 95–100%, depending on the EO and grain type). Palmarosa EO effectively inhibited the growth of F. graminearum at concentrations of 5% and 10%, with the ERG reduction at a level of 96–100%. In the 0.1% EO concentration, the reduction in ERG amount was strongly differentiated, depending on both the EO type and the cereal grain. In wheat grain, ERG was reduced at levels of 55–65%, depending on the EO type, while in the maize grain, only cinnamon EO exhibited significant fungistatic activity compared to the control sample.
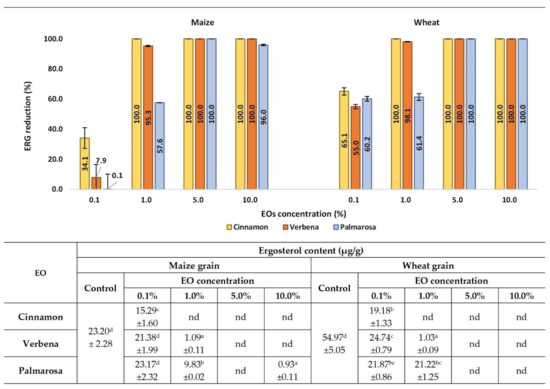
Figure 4.
Effects of EOs on ERG contamination in maize and wheat samples after inoculation with the F. graminearum. nd-not detected, below the limit of quantification (<0.01 µg/g). Averages with different letters (a–d) for each cereal are significantly different at the p < 0.05.
3.4. The Effect of EO Concentration on the ZEA Level in Maize and Wheat Grain
The reduction of ZEA concentrations in cereals treated with EOs in general depended on EO concentration (Figure 5). The greatest degree of ZEA content reduction by all tested EOs, both in maize and wheat grains, was observed at EO concentrations from 5% to 10%. The percentage of ZEA reduction was close to or exceeded 99%, depending on the EO. All tested EOs significantly decreased ZEA content at a concentration of 1%, however, cinnamon and verbena EOs were more effective, with ZEA reduction at levels of 96% to 100%, depending on the EO and grain type. The percentage values of ZEA reduction by palmarosa EO were 84.6% and 66.2% in maize and wheat grain, respectively. At the concentration of 0.1%, only cinnamon and verbena EOs significantly reduced ZEA in cereal samples, with the highest efficacy being cinnamon EO in maize (62.2% ZEA content reduction) and verbena EO in wheat (88.3% ZEA reduction). Based on the obtained result, it should be noticed that ZEA concentrations in samples treated with palmarosa EO at a concentration of 0.1% was higher compared to the control, however, only in wheat grain was the difference significant.
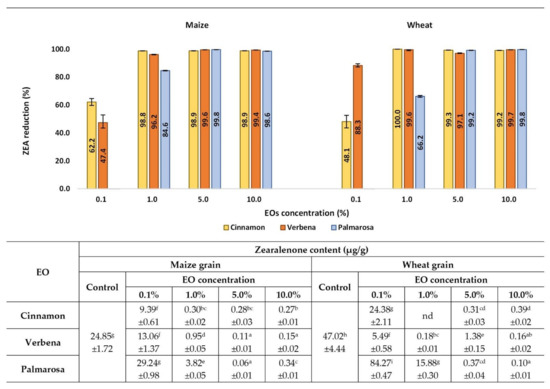
Figure 5.
Effects of EOs on ZEA contamination in maize and wheat samples after inoculation with the F. graminearum. nd-not detected, below the limit of quantification (<0.01 µg/g). Averages with different letters (a–i) for each cereal are significantly different at the p < 0.05.
3.5. The Effect of EO Concentration on the DON Level in Maize and Wheat Grain
DON contamination in cereal samples also depended on the concentration of EOs, however, the types of cereals and EOs likewise influenced the level of toxin reduction (Figure 6). It is worth underlining that F. graminearum produced a low amount of DON compared to ZEA, both in maize and wheat grain (the level of DON concentration in control samples reached 3.49 and 3.19 μg/g, respectively). In wheat grain samples impregnated with tested EOs in concentrations from 1% to 10%, DON was not detected (100% of toxin reduction was noticed compared to control). At the concentration of 0.1%, no significant reduction in DON content was observed, while in samples treated with cinnamon and palmarosa EOs, a significantly higher content of the toxin was observed. In maize grain, the highest efficacy in DON reduction was demonstrated by cinnamon EO, decreasing toxin content in the whole tested range of concentration. The highest percentage reduction of the toxin was observed at the range of concentration 1% to 5% with lower efficacy at the concentration 10% (72.4% of DON reduction). Verbena and palmarosa EOs effectively reduced DON content at a concentration of 10%, while at lower concentrations, their effectiveness was much weaker. It is worth underlining that at a concentration of 0.1%, only cinnamon EO showed a significant reduction, while in samples treated with verbena and palmarosa, significantly higher content was observed.
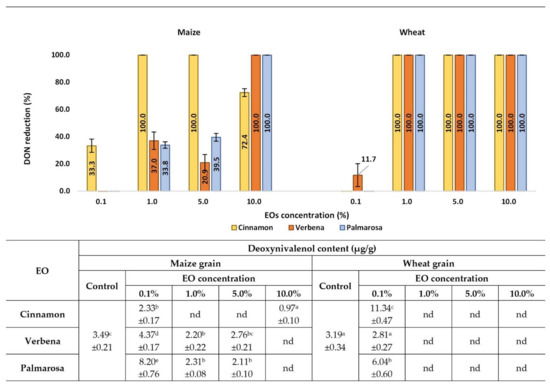
Figure 6.
Effects of EOs on DON contamination in maize and wheat samples after inoculation with the F. graminearum. nd-not detected, below the limit of quantification (<0.01 µg/g). Averages with different letters (a–e) for each cereal are significantly different at the p < 0.05.
4. Discussion
The occurrence of filamentous fungi and their toxic metabolites in food and feed has become one of the biggest global problems [32]. Fusarium fungi and mycotoxins constitute a significant negative factor in food production, due to the infestation of economically important cereal crops and adverse effects on human and animal health [33]. Therefore, providing a suitable amount of good quality and safe food with respect to environmental conditions is one of the priority goals in modern food production, especially at the agricultural stage [34]. There are many concepts focused on the issue of sustainable, eco-friendly food production, such as sustainable agriculture development [35], One Health concept [36] or Climate-smart agriculture (CSA) [34]. Among various methods currently used to reduce the presence of fungi and mycotoxins in food, such as physical, chemical, biological methods or good agrotechnical practices [37,38], more emphasis is placed on biological methods, especially in the context of limiting the use of pesticides in agriculture production [39]. Many data indicate the high potential of natural substances of plant origin as biological control agents, such as EOs, which are regarded as efficient, safe, environmentally friendly factors [40]. In this research, three EOs in different concentrations were tested as biological control agents against Fusarium graminearum growth and mycotoxin synthesis in maize and wheat grain. The presented work shows a different approach to estimating the influence of EOs on fungal growth under in vitro conditions: volatilization influence on a Petri dish and chromatographic analyses of ERG in cereal grains, as well as level of DON and ZEA reduction in maize and wheat grain using the HPLC method, were estimated. Additionally, the effects of EOs on fungal hyphae were investigated using fluorescent microscope analysis.
The conducted research showed that the EOs used exhibit antagonistic activity against F. graminearum and also reduce levels of DON and ZEA in wheat and maize grain. Better results were observed for the higher concentrations of EOs (10% and 5%), and cinnamon and verbena EOs were more effective than palmarosa. Antifungal activity was strongly dependent on the EO composition. Cinnamon (Cinnamomum zeylanicum) is a spice collected from the bark of several trees from the genus Cinnamomum and Lauraceae family. The oil from cinnamon bark oil is rich in eugenol, cinnamaldehyde, and linalool. It is also rich in β-caryophyllene and other terpenes. The antifungal properties of cinnamon EOs have been reported as effective against pathogenic Fusarium culmorum and Fusarium verticillioides [22], Aspergillus flavus and Fusarium moniliforme [41], and Villosiclava virens. Additionally, cinnamon water filtrate has been tested as an antifungal Botrytis cinerea agent [42], and cinnamon oil nanoemulsion has been reported to have an inhibitory effect on the mycelial growth of Aspergillus niger [43]. Lemon verbena is a perennial flowering plant of the verbena family (Verbenaceae). Its EOs have been reported to have antimicrobial and antioxidant properties. The most constituent compounds of verbena are geranial, neral, limonene, 1,8-cineole, spathulenol, nerol, geraniol, trans-β-caryophyllene, and geranyl acetate. According to researchers’ findings, these components act as fumigants, insect repellents, or insecticides [44]. Moreover, the antimicrobial properties of verbena EOs have been reported as effective against the pathogens Shigella dysenteriae, Salmonella typhimurium, Pseudomonas aeruginosa [45], F. avenaceum, F. culmorum, F. graminearum, and F. oxysporum [46]. The palmarosa EO has an antifungal activity well documented in the literature, and its biological activity has been studied in the last years. The main constituents of palmarosa EO are geraniol (82%), geranyl acetate (9%), linalool (2%), and trans-β-ocimene (1%), whereas geraniol is the main constituent associated with its antimicrobial activity [47]. The use of palmarosa EO presents promising results in the control of phytopathogens [23]. It is worth emphasizing that growth inhibition of F. graminearum was greater in wheat and maize grain, especially in EO concentrations of 1% and 0.1%, compared to the volatilization effect on Petri dishes.
Moreover, it is worth underlining that the concentration of EOs strongly influenced not only the growth of fungi, but also the mycotoxin biosynthesis. As the results show, the higher concentrations of EOs completely inhibited the growth of fungi, however, the influence of the lower concentration (0.1%) on the fungal growth was also evinced by ERG reduction. The volatilization method showed only the effects of higher doses of EOs. Some authors also pay attention to the dependence of the degree of fungal growth inhibition and mycotoxin level decrease on the EO concentration. Wan et al. [48] used different concentrations of five encapsulated EOs (concentration in rice culture: 1250 µg/g of bulk EOs, 1250 µg/g of EO nanoemulsions and 125 µg/g of 10-fold diluted EO nanoemulsions) to study their efficacy in the inhibition of F. graminearum growth and mycotoxin (DON, 3ADON, 15ADON) production. The authors stated that inhibition of mycelium growth and reduction in mycotoxin production were dose-dependent, similarly to the present work. Olosunde et al. [49] observed that with the increasing dose of Aframomum danielli, the degree of fumonisins reduction was higher. Application of 0.25% EO caused an 8% reduction in FB1, while the dose of 10% resulted in 76% reduction. Sumalan [50] also reported that a reduction in fumonisins was dose-dependent. In this study, it should be noted that the degree of reduction in toxin amount was not always consistent with the degree of ERG reduction. It is especially important to take into account the design of the appropriate amount of EO to apply. While the reduction in ZEA was consistent with the inhibition of ERG biosynthesis, the reduction in DON was not consistent with this parameter. In the research of da Silva Bomfim et al. [51] concerning the antifungal and antimycotoxigenic properties of rosemary EO, the production of aflatoxins was inhibited at concentrations lower than the concentration demonstrating fungistatic activity. The growth of Aspergillus flavus was inhibited with both MIC and MFC at 500 µg/mL, while the production of aflatoxins B1 and B2 was reduced after treatment with 250 µg/mL of rosemary EO. Similar results were obtained by Rasooli & Owlia [52] with the essential oils from Thymus eriocalyx and Thymus X-porlock. Some other authors also reported antimycotoxigenic properties at lower concentrations of EO than the concentration showing fungicidal activity [53,54].
Microscopic analysis revealed that contact of fungal cells with EOs caused morphological changes, the intensity of which depended on the oil concentration. The deformations of individual hyphae were already visible in a concentration of 0.1% of EO, while concentrations of 1% and above caused a significant decrease in viability, visible in fluorescent staining and cell deformations. Fluorescence-based microscopic methods have been used for viability testing of different bacteria, yeasts, and filamentous fungi [55,56,57,58]. The combination of FDA or cFDA and PI stains is often used to observe the effect of some antimicrobial agents on the microbial cells. For example, Li et al. [59] described the antifungal effect of a novel compound, CF66I, produced by Burkholeria cepacia against Fusarium solani, using dual staining with propidium iodide (PI) and fluorescein diacetate (FDA) to observe morphological changes. Incubation with high doses of CF66I revealed collapsed hyphae, indicating cell death and membrane permeation. In the study of Kim et a. [60], who used only PI staining, fungal hyphae treated with PGP-C (lipopeptide isolated from Paenibacillus elgii JCK1400 culture) showed strong red color, which indicates damage of the cell membrane. PI is a DNA-intercalating fluorescent dye that can penetrate only dead cells, in response to membrane damage.
A greater knowledge and understanding the mode of action of EOs is necessary to plan the direction and form of their application, as well as to increase the scale of pro-duction. The presented characteristics of EOs, including their antifungal and antimycotoxigenic properties, clearly indicated that their activity is dependent on the concentration used. As some authors underline, application method could also affect the effectiveness of EOs. Suhr and Nielsen [61] determined the antifungal activity of some EOs, and have stated that larger compounds, such as thymol and eugenol, demonstrated the best effects against rye bread spoilage after application directly to the medium, whereas smaller compounds, such as allyl isothiocyanate and citral, were most efficient when added as volatiles. The future application should also be considered when choosing a research method and experiment conditions, such as medium or environmental factors. Some authors indicated the lack of congruence between agar media assays and real food studies [62,63] (Arras and Usai 2001; Ultee and Smid 2001). Therefore, in the presented work, the experiments that were conducted on wheat and maize grain treated with EOs simulated the potential seed treatment. Moreover, results obtained by volatilization method can be taken into account when considering the use of fumigation.
5. Conclusions
In conclusion, the data described in this study demonstrate that EOs of cinnamon bark, palmarosa, and verbena were efficient at inhibiting F. graminearum growth as decreased mycelium growth, and alterations in morphological structures and ergosterol quantification were observed. The current study demonstrated that all three tested EOs effectively suppressed the development of F. graminearum in concentrations of 5 and 10%. The in vitro fungistatic effect was dependent on the tested EOs as well as on its concentration. Among them, cinnamon and verbena EOs displayed the strongest in vitro antifungal activity against the tested fungi. Furthermore, the tested natural plant compounds proved to be effective in mycotoxin production-inhibition, and may be an alternative to synthetic chemical agents. The high importance of the tested EOs’ antimycotoxigenic properties is related to the damage that these Fusarium species mycotoxins cause to human and animal health. Recently developed fungicides often demonstrate systemic or curative properties aimed at effective disease control, but have limited applications at low rates. Furthermore, the emerging problem of fungicide-resistant microorganisms urges researchers to develop new biocontrol agents or fungicides possessing novel action mechanisms. Therefore, cinnamon bark, palmarosa leaves, and verbena leaf and flower EOs were proved to be potential biological agents for the control of F. graminearum diseases in crops and ZEA and DON mycotoxin production. Future in vitro tests are mandatory, including tests on different Fusarium species and mycotoxins, as well as concerning EO composition, in order to evaluate the broad spectrum of fungistatic activity as well as the inhibition of varied mycotoxin production. Additional advanced greenhouses and field in vivo research are needed in order to determine the potential usefulness of palmarosa, verbena, and cinnamon bark EOs in fungal control programs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app12010473/s1. Figure S1. Morphology of F. graminearum hyphae under light microscope (magnification × 400) treated with verbena EO: (a) Control; (b) EO concentration 0.1%; (c) EO concentration 1.0%; (d) EO concentration 5.0%; (e) EO concentration 10.0%; Figure S2. Morphology of F. graminearum hyphae under optical microscope (magnification × 400): untreated: (a) control, stained with cFDA; (b) control, stained with PI; treated with palmarosa EO: stained with cFDA, EO concentration: (c) 0.1%; (e) 1.0%; (g) 5.0%; (i) 10.0%; stained with PI, EO concentration: (d) 0.1%; (f) 1.0%; (h) 5.0%; (j) 10.0%; Figure S3. Morphology of F. graminearum hyphae under optical microscope (magnification × 400): untreated: (a) control, stained with cFDA; (b) control, stained with PI; treated with verbena EO: stained with cFDA, EO concentration: (c) 0.1%; (e) 1.0%; (g) 5.0%; (i) 10.0%; stained with PI, EO concentration: (d) 0.1%; (f) 1.0%; (h) 5.0%; (j) 10.0%.
Author Contributions
Conceptualization, D.G., K.M. and K.J.; methodology, D.G, K.M., K.J. and A.W.; validation, K.J. and A.W.; formal analysis, K.M., D.G., R.G. and R.K.; investigation, D.G., K.M., K.J., P.A.U., R.G., A.W. and R.K.; resources, D.G., K.M., K.J. and R.G.; data curation, K.J. and A.W.; writing—original draft preparation, D.G., K.M., K.J. and A.W.; writing—review and editing, D.G., K.M. and K.J.; visualization, D.G., K.J. and K.M.; supervision, D.G.; project administration, K.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Polish National Science Centre, grant number 2018/31/B/NZ9/03485.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shahbandeh, M. Worldwide Production of Grain in 2020/21, by Type. Statista 2021. Available online: https://www.statista.com/statistics/263977/world-grain-production-by-type/ (accessed on 24 September 2021).
- Ranum, P.; Peña-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Deepa, N.; Nagaraja, H.; Sreenivasa, M.Y. Prevalence of fumonisin producing Fusarium verticillioides associated with cereals grown in Karnataka (India). Food Sci. Hum. Wellness 2016, 5, 156–162. [Google Scholar] [CrossRef]
- Ferreira, F.M.D.; Hirooka, E.Y.; Ferreira, F.D.; Silva, M.V.; Mossini, S.A.G.; Machinski, M., Jr. Effect of Zingiber officinale Roscoe essential oil in fungus control and deoxynivalenol production of Fusarium graminearum Schwabe in vitro. Food Addit. Contam. Part A 2018, 35, 2168–2174. [Google Scholar] [CrossRef]
- Lofgren, L.; Riddle, J.; Dong, Y.; Kuhnem, P.R.; Cummings, J.A.; Del Ponte, E.M.; Bergstrom, G.C.; Kistler, H.C. A high proportion of NX-2 genotype strains are found among Fusarium graminearum isolates from northeastern New York State. Eur. J. Plant Pathol. 2018, 150, 791–796. [Google Scholar] [CrossRef]
- Karlovsky, P. Biological detoxification of the mycotoxin deoxynivalenol and its use in genetically engineered crops and feed additives. Appl. Microbiol. Biotechnol. 2011, 91, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Dweba, C.C.; Figlan, S.; Shimelis, H.A.; Motaung, T.E.; Sydenham, S.; Mwadzingeni, L.; Tsilo, T.J. Fusarium head blight of wheat: Pathogenesis and control strategies. Crop Prot. 2017, 91, 114–122. [Google Scholar] [CrossRef]
- Palacios, S.A.; Erazo, J.G.; Ciasca, B.; Lattanzio, V.M.T.; Reynoso, M.M.; Farnochi, M.C.; Torres, A.M. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat from Argentina. Food Chem. 2017, 230, 728–734. [Google Scholar] [CrossRef]
- Escrivá, L.; Font, G.; Manyes, L. In vivo toxicity studies of fusarium mycotoxins in the last decade: A review. Food Chem. Toxicol. 2015, 78, 185–206. [Google Scholar] [CrossRef]
- Kelly, A.C.; Clear, R.M.; O’Donnell, K.; McCormick, S.; Turkington, T.K.; Tekauz, A.; Gilbert, J.; Kistler, H.C.; Busman, M.; Ward, T.J. Diversity of Fusarium head blight populations and trichothecene toxin types reveals regional differences in pathogen composition and temporal dynamics. Fungal Genet. Biol. 2015, 82, 22–31. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Serrano, M.; Bailén, G.; Guillén, F.; Zapata, P.J.; Valverde, J.M.; Castillo, S.; Fuentes, M.; Valero, D. The use of a natural fungicide as an alternative to preharvest synthetic fungicide treatments to control lettuce deterioration during postharvest storage. Postharvest Biol. Technol. 2008, 47, 54–60. [Google Scholar] [CrossRef]
- Boukaew, S.; Prasertsan, P.; Sattayasamitsathit, S. Evaluation of antifungal activity of essential oils against aflatoxigenic Aspergillus flavus and their allelopathic activity from fumigation to protect maize seeds during storage. Ind. Crops Prod. 2017, 97, 558–566. [Google Scholar] [CrossRef]
- Sumalan, R.M.; Alexa, E.; Popescu, I.; Negrea, M.; Radulov, I.; Obistioiu, D.; Cocan, I. Exploring Ecological Alternatives for Crop Protection Using Coriandrum sativum Essential Oil. Molecules 2019, 24, 2040. [Google Scholar] [CrossRef] [PubMed]
- Tongnuanchan, P.; Benjakul, S. Essential Oils: Extraction, Bioactivities, and Their Uses for Food Preservation. J. Food Sci. 2014, 79, R1231–R1249. [Google Scholar] [CrossRef] [PubMed]
- Fornari, T.; Vicente, G.; Vázquez, E.; García-Risco, M.R.; Reglero, G. Isolation of essential oil from different plants and herbs by supercritical fluid extraction. J. Chromatogr. A 2012, 1250, 34–48. [Google Scholar] [CrossRef]
- Ayvaz, A.; Sagdic, O.; Karaborklu, S.; Ozturk, I. Insecticidal Activity of the Essential Oils from Different Plants Against Three Stored-Product Insects. J. Insect Sci. 2010, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Bertuzzi, G. Antioxidative Action of Citrus limonum Essential Oil on Skin. Eur. J. Med. Plants 2013, 3, 1–9. [Google Scholar] [CrossRef]
- Properzi, A.; Angelini, P.; Venanzoni, R. Some Biological Activities of Essential Oils. Med. Aromat. Plants 2013, 2, 1000136. [Google Scholar] [CrossRef]
- Prakash, B.; Kedia, A.; Mishra, P.K.; Dubey, N.K. Plant essential oils as food preservatives to control moulds, mycotoxin contamination and oxidative deterioration of agri-food commodities—Potentials and challenges. Food Control 2015, 47, 381–391. [Google Scholar] [CrossRef]
- Gakuubi, M.M.; Maina, A.W.; Wagacha, J.M. Antifungal Activity of Essential Oil of Eucalyptus camaldulensis Dehnh. against Selected Fusarium spp. Int. J. Microbiol. 2017, 2017. [Google Scholar] [CrossRef]
- Sharma, A.; Rajendran, S.; Srivastava, A.; Sharma, S.; Kundu, B. Antifungal activities of selected essential oils against Fusarium oxysporum f. sp. lycopersici 1322, with emphasis on Syzygium aromaticum essential oil. J. Biosci. Bioeng. 2017, 123, 308–313. [Google Scholar] [CrossRef]
- Roselló, J.; Sempere, F.; Sanz-Berzosa, I.; Chiralt, A.; Santamarina, M.P. Antifungal activity and potential use of essential oils against Fusarium culmorum and Fusarium verticillioides. J. Essent. Oil-Bear. Plants 2015, 18, 359–367. [Google Scholar] [CrossRef]
- Kalagatur, N.K.; Nirmal Ghosh, O.S.; Sundararaj, N.; Mudili, V. Antifungal Activity of Chitosan Nanoparticles Encapsulated With Cymbopogon martinii Essential Oil on Plant Pathogenic Fungi Fusarium graminearum. Front. Pharmacol. 2018, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Soylu, E.M.; Soylu, S.; Kurt, S. Antimicrobial activities of the essential oils of various plants against tomato late blight disease agent Phytophthora infestans. Mycopathologia 2006, 161, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Perumal, A.B.; Sellamuthu, P.S.; Nambiar, R.B.; Sadiku, E.R. Antifungal activity of five different essential oils in vapour phase for the control of Colletotrichum gloeosporioides and Lasiodiplodia theobromae in vitro and on mango. Int. J. Food Sci. Technol. 2016, 51, 411–418. [Google Scholar] [CrossRef]
- Brul, S.; Nussbaum, J.; Dielbandhoesing, S.K. Fluorescent probes for wall porosity and membrane integrity in filamentous fungi. J. Microbiol. Methods 1997, 28, 169–178. [Google Scholar] [CrossRef]
- Chitarra, G.S.; Breeuwer, P.; Nout, M.J.R.; Van Aelst, A.C.; Rombouts, F.M.; Abee, T. An antifungal compound produced by Bacillus subtilis YM 10–20 inhibits germination of Penicillium roqueforti conidiospores. J. Appl. Microbiol. 2003, 94, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Perczak, A.; Gwiazdowska, D.; Marchwińska, K.; Juś, K.; Gwiazdowski, R.; Waśkiewicz, A. Antifungal activity of selected essential oils against Fusarium culmorum and F. graminearum and their secondary metabolites in wheat seeds. Arch. Microbiol. 2019, 201, 1085–1097. [Google Scholar] [CrossRef]
- Perczak, A.; Gwiazdowska, D.; Gwiazdowski, R.; Juś, K.; Marchwińska, K.; Waśkiewicz, A. The Inhibitory Potential of Selected Essential Oils on Fusarium spp. Growth and Mycotoxins Biosynthesis in Maize Seeds. Pathogens 2020, 9, 23. [Google Scholar] [CrossRef]
- Breeuwer, P.; Drocourt, J.L.; Bunschoten, N.; Zwietering, M.H.; Rombouts, F.M.; Abee, T. Characterization of uptake and hydrolysis of fluorescein diacetate and carboxyfluorescein diacetate by intracellular esterases in Saccharomyces cerevisiae, which result in accumulation of fluorescent product. Appl. Environ. Microbiol. 1995, 61, 1614–1619. [Google Scholar] [CrossRef]
- Oparka, K.J. Uptake and compartmentation of fluorescent probes by plant cells. J. Exp. Bot. 1991, 42, 565–579. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide contamination of food-crops with mycotoxins: Validity of the widely cited ‘FAO estimate’ of 25%. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Chilaka, C.; De Boevre, M.; Atanda, O.; De Saeger, S. Occurrence of Fusarium Mycotoxins in Cereal Crops and Processed Products (Ogi) from Nigeria. Toxins 2016, 8, 342. [Google Scholar] [CrossRef]
- Singh, R.; Singh, G.S. Traditional agriculture: A climate-smart approach for sustainable food production. Energy Ecol. Environ. 2017, 2, 296–316. [Google Scholar] [CrossRef]
- Żmija, D. Zrównoważony rozwój rolnictwa i obszarów wiejskich w Polsce. Studia Ekon. 2014, 166, 149–158. [Google Scholar]
- Garcia, S.N.; Osburn, B.I.; Jay-Russell, M.T. One Health for Food Safety, Food Security, and Sustainable Food Production. Front. Sustain. Food Syst. 2020, 4, 1. [Google Scholar] [CrossRef]
- Patriarca, A.; Fernández Pinto, V. Prevalence of mycotoxins in foods and decontamination. Curr. Opin. Food Sci. 2017, 14, 50–60. [Google Scholar] [CrossRef]
- Čolović, R.; Puvača, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Đuragić, O.; Kos, J.; Pinotti, L. Decontamination of Mycotoxin-Contaminated Feedstuffs and Compound Feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef]
- Bubici, G.; Kaushal, M.; Prigigallo, M.I.; Cabanás, C.G.L.; Mercado-Blanco, J. Biological control agents against Fusarium wilt of banana. Front. Microbiol. 2019, 10, 616. [Google Scholar] [CrossRef]
- Singh, P.; Pandey, A.K. Prospective of essential oils of the genus mentha as biopesticides: A review. Front. Plant Sci. 2018, 9, 1295. [Google Scholar] [CrossRef] [PubMed]
- Massoud, M.A.; Saad, A.S.A.; Soliman, E.A.; El-Moghazy, A.Y. Antifungal activity of some essential oils applied as fumigants against two stored grains fungi. J. Adv. Agric. Res. 2012, 17, 296–306. [Google Scholar]
- Kowalska, J.; Tyburski, J.; Krzymińska, J.; Jakubowska, M. Cinnamon powder: An in vitro and in vivo evaluation of antifungal and plant growth promoting activity. Eur. J. Plant Pathol. 2020, 156, 237–243. [Google Scholar] [CrossRef]
- Wu, D.; Lu, J.; Zhong, S.; Schwarz, P.; Chen, B.; Rao, J. Influence of nonionic and ionic surfactants on the antifungal and mycotoxin inhibitory efficacy of cinnamon oil nanoemulsions. Food Funct. 2019, 10, 2817–2827. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, H.; Mehrafarin, A.; Omidi, H.; Badi, H.N.; Khalighi-Sigaroodi, F. Evaluation of Phytochemical and Biochemical Patterns of Lemon Verbena (Lippia citriodora H.B.K.) at Different Temperatures. Res. J. Pharmacogn. 2019, 6, 9–24. [Google Scholar] [CrossRef]
- Hosseini, M.; Jamshidi, A.; Raeisi, M.; Azizzadeh, M. The Antibacterial and Antioxidant Effects of Clove (Syzygium aromaticum) and Lemon Verbena (Aloysia citriodora) Essential Oils. J. Hum. Environ. Health Promot. 2019, 5, 86–93. [Google Scholar] [CrossRef]
- Krzyśko-Łupicka, T.; Sokół, S.; Piekarska-Stachowiak, A. Evaluation of Fungistatic Activity of Eight Selected Essential Oils on Four Heterogeneous Fusarium Isolates Obtained from Cereal Grains in Southern Poland. Molecules 2020, 25, 292. [Google Scholar] [CrossRef]
- da Silva Franca, K.R.; dos Santos, X.A.L.; de Alves, F.M.F.; Lima, T.S.; de Araújo, I.G.; da Nóbrega, L.P. Control of Fusarium verticillioides using Palmarosa Essential Oil (Cymbopogon martinii) Maracajá View project Ecology of waterbirds in semi-arid lands View project. Artic. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 484–494. [Google Scholar] [CrossRef]
- Wan, J.; Zhong, S.; Schwarz, P.; Chen, B.; Rao, J. Physical properties, antifungal and mycotoxin inhibitory activities of five essential oil nanoemulsions: Impact of oil compositions and processing parameters. Food Chem. 2019, 291, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Olosunde, O.O.; Adegoke, G.O.; Abiodun, O.A. Composition of sorghum-millet flour, Aframomum danielli essential oil and their effect on mycotoxins in kunu zaki. Afr. J. Food Sci. 2015, 9, 411–416. [Google Scholar] [CrossRef]
- Sumalan, R.N.; Ersilia, A.; Mariana, A.P. Assessment of inhibitory potential of essential oils on natural mycoflora and fusarium mycotoxins production in wheat. Chem. Cent. J. 2013, 7, 32. [Google Scholar] [CrossRef]
- da Silva Bomfim, N.; Kohiyama, C.Y.; Nakasugi, L.P.; Nerilo, S.B.; Mossini, S.A.G.; Romoli, J.C.Z.; Mikcha, J.M.G.; de Abreu Filho, B.A.; Machinski, M., Jr. Antifungal and antiaflatoxigenic activity of rosemary essential oil (Rosmarinus officinalis L.) against Aspergillus flavus. Food Addit. Contam. Part A 2020, 37, 153–161. [Google Scholar] [CrossRef]
- Rasooli, I.; Owlia, P. Chemoprevention by thyme oils of Aspergillus parasiticus growth and aflatoxin production. Phytochemistry 2005, 66, 2851–2856. [Google Scholar] [CrossRef]
- Tian, J.; Huang, B.; Luo, X.; Zeng, H.; Ban, X.; He, J.; Wang, Y. The control of Aspergillus flavus with Cinnamomun jensenianum hand.—Mazz essential oil and its potential use as a food preservative. Food Chem. 2012, 130, 520–527. [Google Scholar] [CrossRef]
- Ferreira, F.D.; Kemmelmeier, C.; Arroteia, C.C.; Costa, C.L.; Mallmann, C.A.; Janeiro, V.; Ferreira, F.M.D.; Mossini, S.A.G.; Silva, E.L.; Machinski, M., Jr. Inhibitory effect of the essential oil of Curcuma longa L. and curcumin on aflatoxin production by Aspergillus flavus link. Food Chem. 2013, 136, 789–793. [Google Scholar] [CrossRef]
- Kessel, G.J.T.; de Hass, B.H.; Lombaers-van der Plas, C.H.; Meijer, E.M.J.; Dewey, F.M.; Gou-driaan, J.; ven der Werf, W.; Kohl, J. Quantification of mycelium of Botrytis spp. and the antagonist Ulocladium atrum in necrotic leaf tissue of Cyclamen and Lily by fluorescence microscopy and image analysis. Phytopathology 1999, 89, 868–876. [Google Scholar] [CrossRef]
- Foglieni, C.; Meoni, C.; Davalli, A.M. Fluorescent dyes for cell viability: An application on prefixed condition. Histochem. Cell Biol. 2001, 115, 223–229. [Google Scholar] [CrossRef]
- Khan, M.M.T.; Pyle, B.H.; Camper, A.K. Specific and rapid enumeration of viable but nonculturable and viable-culturable gram-negative bacteria by using flow cytometry. Appl. Environ. Microbiol. 2010, 76, 5088–5096. [Google Scholar] [CrossRef]
- Fernandez-Miranda, E.; Majada, J.; Casares, A. Efficacy of propidium iodide and FUN-1 stains for assessing viability in basidiospores of Rhizopogon roseolus. Mycologia 2017, 109, 350–358. [Google Scholar] [CrossRef]
- Li, X.; Quan, C.S.; Yu, H.Y.; Wang, J.H.; Fan, S.D. Assessment of an-tifungal effects of a novel compound from Burkholderia cepacia against Fusarium solani by fluorescent staining. World J. Microbiol. Biotechnol. 2009, 25, 151–154. [Google Scholar] [CrossRef]
- Kim, J.; Le, K.D.; Yu, N.H.; Kim, J.I.; Kim, J.C.; Lee, C.W. Structure and antifungal activity of pelgipeptins from Paenibacillus elgii against phytopathogenic fungi. Pestic. Biochem. Physiol. 2020, 163, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Suhr, K.I.; Nielsen, P.V. Antifungal activity of essential oils evaluated by two different application techniques against rye bread spoilage fungi. J. Appl. Microbiol. 2003, 94, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Arras, G.; Usai, M. Fungitoxic activity of 12 essential oils against four postharvest citrus pathogens: Chemical analysis of Thymus capitatus oil and its effect in subatmospheric pressure conditions. J. Food Prot. 2001, 64, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Smid, E.J. Influence of Carvacrol on growth and toxin pro-duction by Bacillus cereus. Int. J. Food Microbiol. 2001, 64, 373–378. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).