Abstract
Elevated intraocular pressure (IOP) has been considered to be the major risk factor for glaucoma. The trabecular meshwork (TM), serving as the primary outflow pathway of aqueous humor (AH), has an essential role in controlling IOP. However, the mechanism of the interaction between ocular hypertension (OHT) and TM has not been completely understood. This research intended to investigate the effect of OHT on cell stiffness, cytoskeletal structure, and vimentin expression of TM cells in rats. Polystyrene microspheres were injected into the left eye of Sprague–Dawley (SD) rats to establish the OHT animal models. TM cells were extracted from the rat eyes with 2-week duration of OHT using the trypsin digestion method. The effect of OHT on the cytoskeletal structure and expression level of vimentin in TM cells were evaluated by immunofluorescence and western blot analyses, respectively. An atomic force microscope (AFM) was used to measure the cell elastic modulus. On the eighth day after microsphere injection, the IOP of experimental eyes increased to 24.25 ± 2.21 mmHg, and the IOP was maintained above 24 mmHg until the end of the experiment. TM cells extracted from the OHT group showed cytoskeleton rearrangement and the formation of cross-linked actin networks (CLANs). Compared to the control group, the cell stiffness in the OHT group was significantly increased. Moreover, the expression level of vimentin was obviously decreased in TM cells of the OHT group. The results demonstrate that OHT induces the rearrangement of the cytoskeleton, increase of cell stiffness, and downregulation of vimentin expression in TM cells.
1. Introduction
Glaucoma is the major irreversible blinding disease worldwide, which is characterized by optic nerve atrophy and visual field defects. It is predicted that 111.8 million people will develop glaucoma by 2040 [1,2]. Primary open-angle glaucoma (POAG) is the most common subtype of glaucoma, and its pathogenesis remains unclear. The homeostasis of AH production and drainage is essential for the maintenance of IOP. Increased AH outflow resistance can lead to increased IOP, which is a significant risk factor for POAG [3]. Lowering IOP is the primary treatment for POAG [4].
The TM is an avascular, structurally complex connective tissue located at the angle between the cornea and the iris [5]. It is the main pathway for AH drainage and provides approximately 50–75% of the resistance to AH outflow [6], so it plays an essential role in regulating the resistance to AH outflow and modulating IOP. The ability of cells to resist stress and maintain shape, which relies on the cytoskeleton [7], is essential for tissue function. Cellular dysfunction in TM can lead to “extra” resistance, resulting in OHT [5]. In this process, cells actively remodel and reorient their cytoskeleton in response to cyclic forces [8]. A previous study has shown that the TM tissue in glaucomatous patients is 20-fold stiffer than that of normal people, and this significant distinction may be associated with alterations in the cytoskeleton [9]. It has been revealed that the cytoskeletal arrangement of TM cells in human glaucomatous eyes is more random and disordered than in control eyes [10]. The cytoskeleton consists mainly of F-actin, microtubules, and intermediate filaments (IFs). Vimentin is one of the major proteins in IFs, which can confer mechanical flexibility to the cell [7]. Therefore, studies of cell mechanics, cytoskeleton, and associated proteins can help to elucidate the interaction mechanisms between TM cells and OHT.
The response of cells to mechanical stimuli is essential for their biological function. Mechanobiology studies how the mechanical forces and mechanical properties of proteins, cells, and tissues exert an influence on physiology and disease [11]. Due to the special location of TM, TM cells are exposed to various mechanical stimuli [5] and are pressure-sensitive. Studies of TM mechanobiology may provide a theoretical basis for the understanding of the development and progression of OHT. To simulate the mechanical changes induced by OHT, researchers applied mechanical stretch, compression, or shear forces to the TM cells cultured in vitro using Flexcell or self-constructed equipment [12]. However, most of the published studies on TM response to mechanical stimuli are performed on a single mechanical factor in vitro, such as stretch, compression, or shear, which is different from the combination of mechanical stimuli TM experience in vivo. Studies of the TM cells extracted from the eyes of the OHT animal model will be helpful to further understand the mechanobiological response of TM to OHT and may contribute to the discovery of new and improved treatments for POAG.
Rodents are the most favored animals in glaucoma models, and the anterior segment of the rat has a high degree of anatomical and developmental similarity to the human eye. The method of inducing OHT by microsphere injection was initially applied to mammals [13], but was later successfully applied to rats and mice [14]. Microspheres accumulate in the TM and Schlemm’s canal (SC) and physically block the AH outflow pathway [15]. To understand the effect of OHT on TM cells, this study first established an OHT rat model by microsphere injection. Then, TM cells were extracted from the OHT and control eyes respectively. Finally, the mechanobiological alterations of these cells, including cytoskeletal morphology, cell elastic modulus, and vimentin expression level, were detected.
2. Materials and Methods
2.1. Animals
This experiment used a total of 8 rats as experimental animals. These rats were healthy adult male SD without eye diseases, weighing 200–250 g. The left eyes were experimental eyes, and the right eyes were control eyes. All animals were reared under standard conditions with free access to water and food. Experimental procedures in this study abided by the ARVO statement for the use of animals in ophthalmic and vision research. In addition, the protocols and standards in this experiment were approved by the Animals Care and Use Committee (IACUC) of Capital Medical University.
2.2. Microspheres Injection
According to the reported method [16,17,18], the rats were anesthetized by intraperitoneal injection of 1% pentobarbital sodium (0.4 mL/100 g body weight), and the surface of the eyeball was anesthetized by oxybuprocaine hydrochloride and compound tropicamide eye drops. Before the injection of microspheres, 10 μL AH was extracted. Then, 10 μL phosphate-buffered saline (PBS) containing 4 × 105 beads/mL microspheres with a diameter of 10 μm was injected into the anterior chamber of the left eyes, using a 30 G needle paralleling to the iris. The right eyes were not injected and acted as control [17,19]. The IOP of the rats was measured using a TonoLab Rebound Tonometer (Icare, Vantaa, Finland) at pre-surgery and every 2 days after injection. The measurement was repeated 5 times for each eye and the average value was taken as the IOP value for the day. After 2 weeks of OHT maintenance, 2 rats were randomly selected for sectioning and staining with hematoxylin-eosin (HE) and used to observe the position of microspheres. Inverted microscopy (Leica, Wetzlar, Germany) was used to detect the distribution of polystyrene microspheres in TM. The remaining six rats were used for extracting TM cells.
2.3. HE Staining
Eyeballs removed from the OHT rats and control rats were fixed in paraformaldehyde for 12 h at 4 °C. Subsequently, the samples were washed 3 times with PBS, dehydrated in ethanol, embedded in paraffin, and sectioned to a thickness of 5 μm. The sections were immersed in xylene and then in alcohol. Finally, they were stained with hematoxylin and eosin.
2.4. Cell Isolation and Culture
Due to the small size of rat TM tissue, we used the trypsin digestion method to extract the primary TM cells. The rat eyeballs were carefully removed and soaked in 0.5% iodoform solution for 5 min and rinsed in PBS 3 times. Under a surgical microscope, the vitreous body, lens, and posterior tissue were removed, and the anterior segment was cut in half. The network tissue with pigment between the cornea and ciliary body is TM. The removed TM tissue was washed in sterile PBS containing penicillin and streptomycin, then cut into small pieces and collected in an EP tube containing 0.125% trypsin solution at 37 ℃ for 1 h. DMEM/F12 medium containing 10% fetal bovine serum was added into the EP tube to terminate digestion. To remove the undigested tissue masses, 70 µm cell strainers were used. The microspheres were removed by centrifugation at 500× g for 3 min. The supernatant was collected and then centrifuged at 106× g for 5 min to collect cell pellets. After that, the pelleted cells were resuspended in the complete medium, plated into 25 mL flasks, and cultured in a 5% CO2 incubator at 37 ℃. The medium was changed every 2 to 3 days. At cell 90% confluence, 0.25% trypsin (Hyclone, Logan, UT, USA) was used for cell passage, and the 2nd to 4th generations of cells were used in this study.
2.5. Immunohistochemical Analyses
In this experiment, TM cells were identified using the Immunohistochemical SABC method, including fibronectin (FN), laminin (LN), and neuron-specific enolase (NSE) staining. The TM cells were fixed with 4% formaldehyde for 30 min at room temperature, and then incubated with 0.2% H2O2 dilution in methanol for 30 min. The samples were permeabilized with 0.3% Triton X-100 for 30 min at room temperature, and blocked with 5% goat serum for 30 min. The primary antibodies were added for incubation for 1 h, and then the samples were washed with PBS 3 times. The primary antibodies used were as follows: FN (1:300, M00564-3, Boster, Wuhan, China), LN (1:300, A03522, Boster), and NSE (1:50, bs-10445R, Bioss, Beijing, China). Biotinylated goat anti-rabbit IgG diluted at 1:100 was added for 30 min. After the samples were washed 3 times, 1:100 diluted SABC was added for 30 min. After washing 3 times, the samples were placed in diaminobenzidine (DAB) solution for color development. The samples were washed with distilled water, then re-stained with hematoxylin. An optical microscope was used to observe the images.
2.6. Cytoskeleton Staining
Phalloidin staining was used to investigate differences in cell cytoskeleton morphology between OHT and control groups, when the cells reached 60% confluency. The TM cells were fixed with 4% paraformaldehyde for 15 min and permeabilized with 0.5% Triton X-100 for 15 min, then blocked with 2.5% BSA for 1 h at room temperature. The TM cells were stained with Phalloidin FITC Phalloidin (1:200, 40735ES75, Yeasen, Shanghai, China) for 30 min to detect cytoskeletal organization. Nuclei were stained with DAPI. A Nikon confocal laser-scanning microscope was used to view and obtain images.
2.7. Atomic Force Microscopy
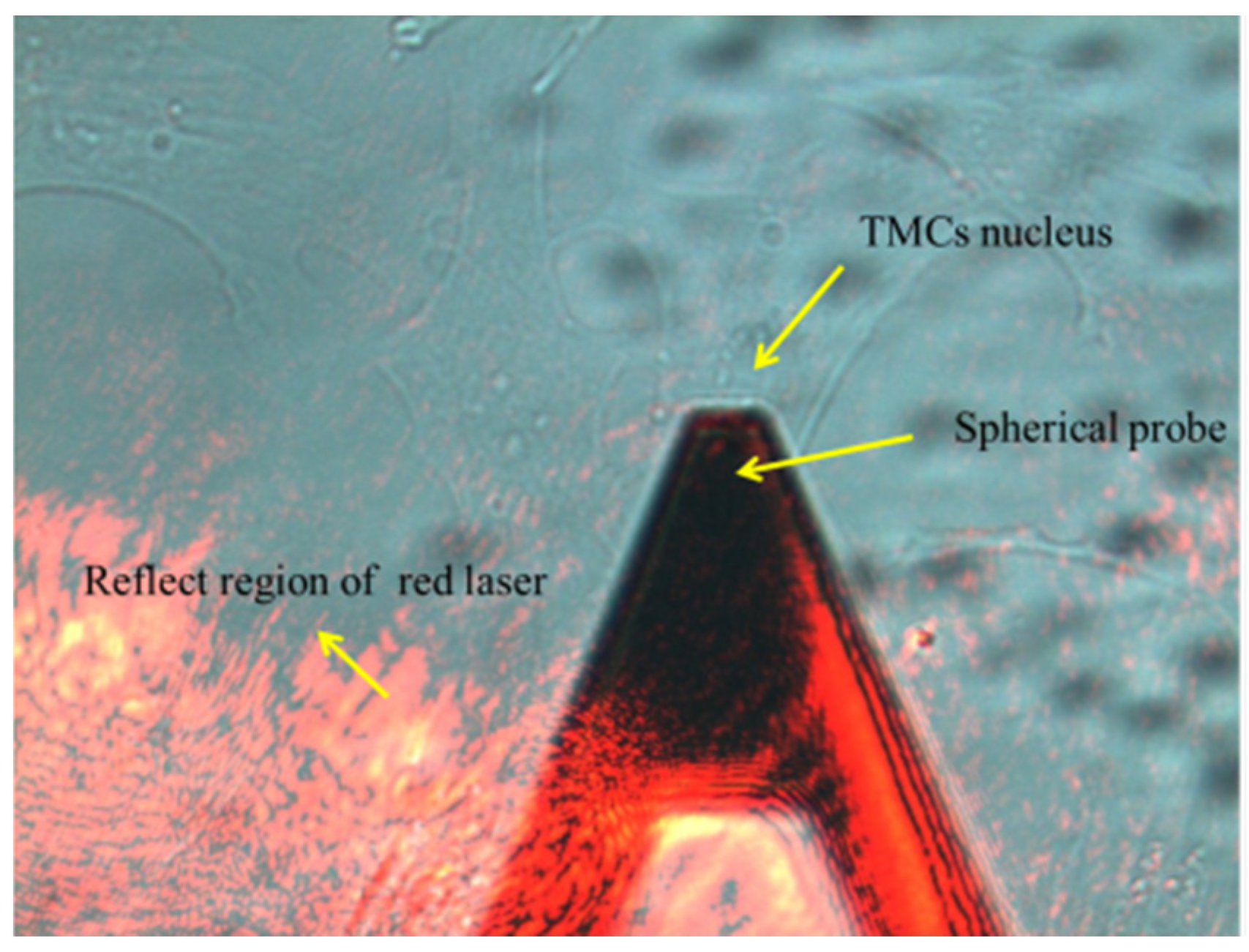
The stiffness of TM cells was detected by atomic force microscopy (AFM, NT-MDT, Ntegra, Moscow, Russia). The cells in each group were inoculated into 3 dishes for the AFM test. The cells were rinsed in Hank’s Balanced Salt Solution (HBSS), and the sensitivity of the four-quadrant photodetector was calibrated before measurement. The culture dish was then placed on the AFM stage, and the probe was moved over the cell under an optical microscope (Figure 1). The spring constant of the spherical probes used in this experiment is 0.05 N/m. Thirty cells were selected from each sample randomly, and each cell was measured 3 times. The elastic modulus was obtained by fitting the curve with the following Hertz equation:
where F denotes force applied by the indenter, E denotes Young’s elastic modulus, and v denotes Poisson’s ratio (v = 0.5) [19]; δ is the indentation depth and R is the probe radius (R = 2.5 μm).
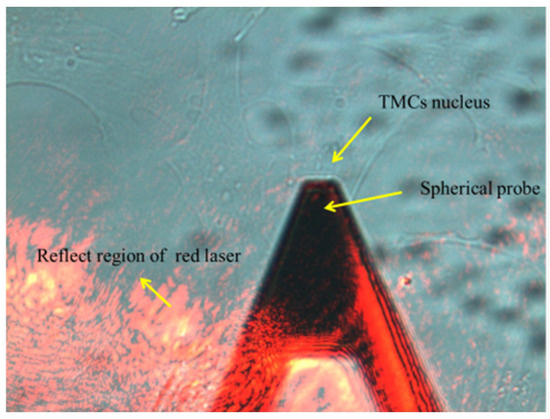
Figure 1.
Mechanical testing of TM cells by AFM.
2.8. Western Blot
Cells were lysed in RIPA buffer containing 1% phosphatase and protease inhibitors on ice. Subsequently, cell lysates were centrifuged at 20,817× g for 20 min to collect proteins. Protein concentrations were subjected to the BCA assay. Then, protein samples were boiled in protein loading buffer and subjected to SDS polyacrylamide gel electrophoresis. Next, proteins were transferred onto PVDF membranes. Membranes were blocked in 5% skimmed milk for 1 h at room temperature, and incubated with primary vimentin antibody (1:1000, bs-8533R, Bioss) overnight at 4 °C. On the following day, the membranes were incubated with horseradish peroxidase-conjugated goat anti-rabbit IgG (1:2000, bs-0295G, Bioss) for 1.5 h at room temperature. Protein bands were visualized using a chemiluminescence imager; β-actin (1:1000, bs-8770R, Bioss) was used as the loading control.
2.9. Statistics
The data were analyzed for normal distribution first. Then, for data that fit the normal distribution, the Student’s t-test was used, and for data that did not, the Mann–Whitney test was used. Statistical analyses were performed using GraphPad Prism version 9.0.0 (GraphPad Software, San Diego, CA, USA). The data were shown as the mean ± standard error of the mean or median with an interquartile range; p < 0.05 was considered significant.
3. Results
3.1. IOP Elevation
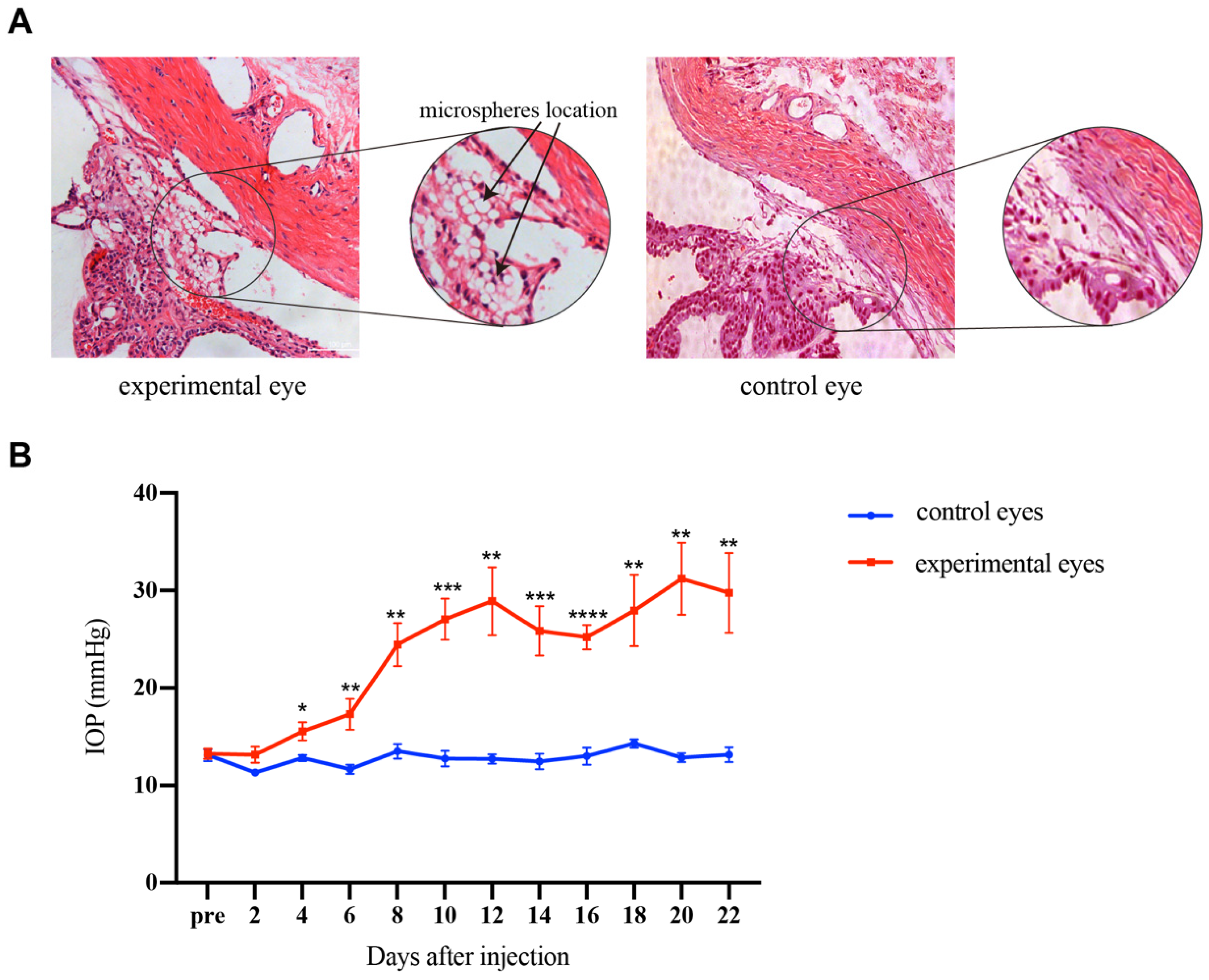
After injecting 10 μL microsphere solution into the anterior chamber, the microspheres were quickly dispersed with the flow of AH and finally accumulated at the anterior chamber angle clearly. There were no serious complications that occurred after the injection. Compared with the normal tissues, the HE staining revealed the location of microspheres in the experimental eyes (Figure 2A). No adhesion was observed between the cornea, iris, and sclera in the eyes. The microspheres did block the TM. Figure 2B shows the change in IOP after injection in the experimental eyes and control eyes in rats. IOP elevated gradually in the experimental eyes. The average IOP of the experimental group was significantly higher than the control group on the 4th day after injection (p < 0.01). The average IOP was maintained above 24 mmHg from the 8th day to the end of the experiment. The results indicated that the OHT rat model was successfully established by the microsphere injection method. Consistent with the results of others [20,21], the model prolonged elevations in IOP for 2 weeks following a single injection.
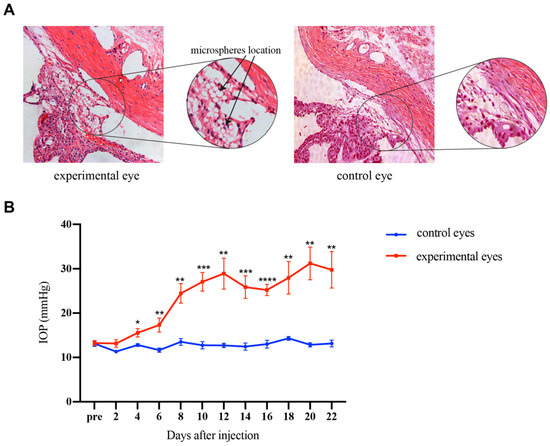
Figure 2.
The distribution of microspheres in TM was viewed by HE staining, and the IOPs of rats were elevated by injection of microspheres. (A) HE staining of the experimental and control eyes. The magnified image showed the position of the injected microspheres, and the black arrow indicated the microspheres. The microspheres were located in the uveoscleral pathway between the sclera and ciliary body and in the corneoscleral angle, which were seen as transparent beads. (B) IOP changes in the experimental and control eyes of rats. On the 4th day after microsphere injection, the IOP in the experimental eyes was significantly higher than in the control eyes and elevated gradually with time. For comparison between groups, IOPs were compared every 2 days after injection, and p values were generated by the Student’s t-test. Scale bar: 100 μm. * p < 0. 05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
3.2. Characteristics of TM Cells
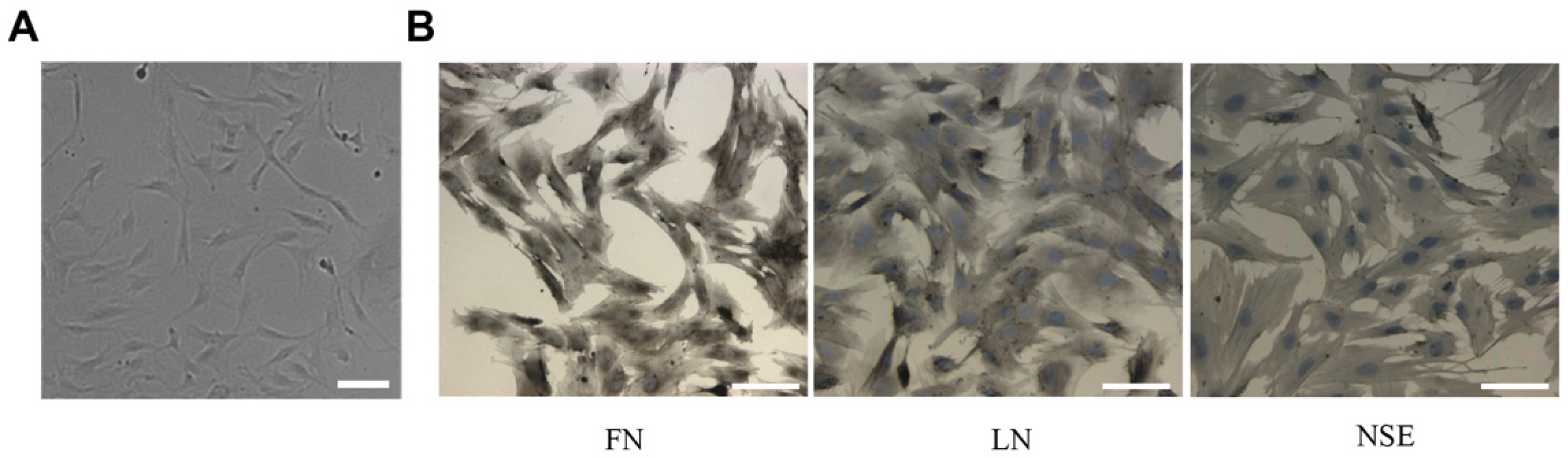
The primary cultured TM cells extracted from the rat models adhered to the substrate after 48 h and reached a fused state after about 10 days. The cells showed typical TM cell morphology: elongated and spindle-like cell (Figure 3A) [5,19]. According to the descriptions in [22,23,24], we detected the TM cells biomarker—FN, LN, and NSE expressions using the immunohistochemical method. As shown in Figure 3B, the TM cells expressed FN, LN, and NSE proteins. By examining cell morphology and biomarkers we could determine that the cells were trabecular meshwork cells.
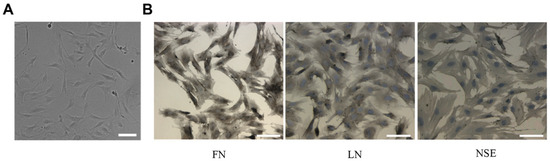
Figure 3.
The primary cultured TM cells identification. (A) Phase-contrast microscopic images of TM cells. The image showed that the cell morphology is mainly spindle-shaped. (B) TM cell biomarker proteins were detected by the immunohistochemical method. The immunohistochemical images of TM cell biomarkers—FN, LN, and NSE showed that the cells positively express FN, LN, and NSE. Scale bar: 50 μm.
3.3. Effects of OHT on Cytoskeleton Morphology in TM Cells
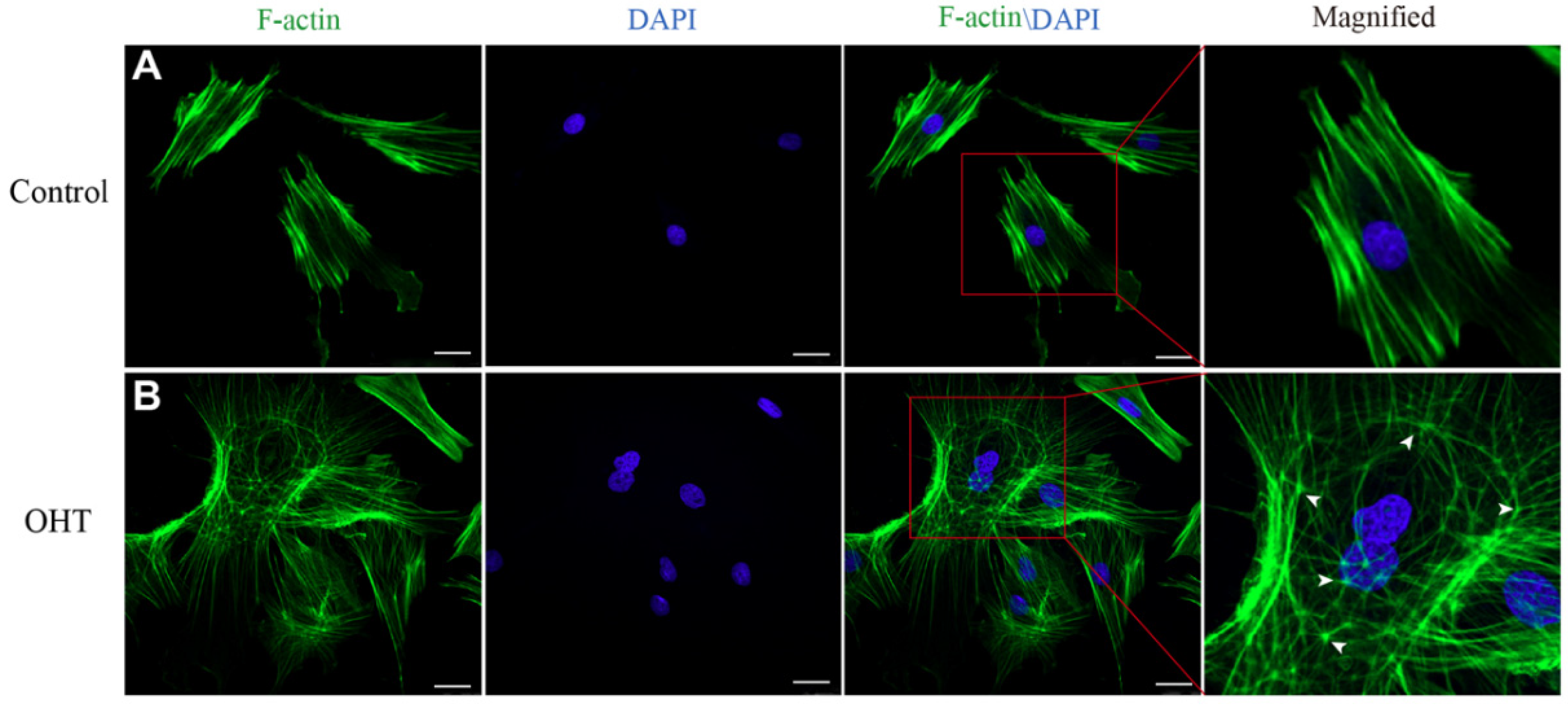
In order to investigate the cytoskeletal changes of TM cells under the effects of OHT, the cytoskeletons of TM cells were stained with FITC Phalloidin. F-actin showed green fluorescence, while the nucleus showed blue fluorescence. We found that the F-actin fibers of control TM cells were regularly arranged along the axial direction, with a bundle-like distribution and a shuttle-shaped cytoskeleton morphology. In contrast, the OHT group had an interlaced and complex arrangement of actin fibers, forming a cross-linked actin networks (CLANs) structure, which was more expanded with thin filamentous pseudopods compared to the control group (Figure 4).
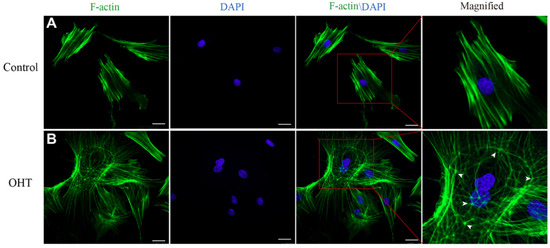
Figure 4.
The cytoskeleton morphology of TM cells. For a clearer view, the cytoskeleton image of a single cell was enlarged. (A) The phalloidin staining images of cytoskeleton F-actin fibers (green) and nuclei (blue) in the control group, and (B) in the OHT group. F-actin fibers of TM cells in the control group were clear and arranged along the long axis, while those in the OHT group were crisscrossed and exhibited affluent thin filamentous pseudopods. The location of CLAN vertices was shown by arrowheads and observed with F-actin labeling. Scale bar: 25 μm.
3.4. Increase of Elastic Moduli of TM Cells Induced by OHT
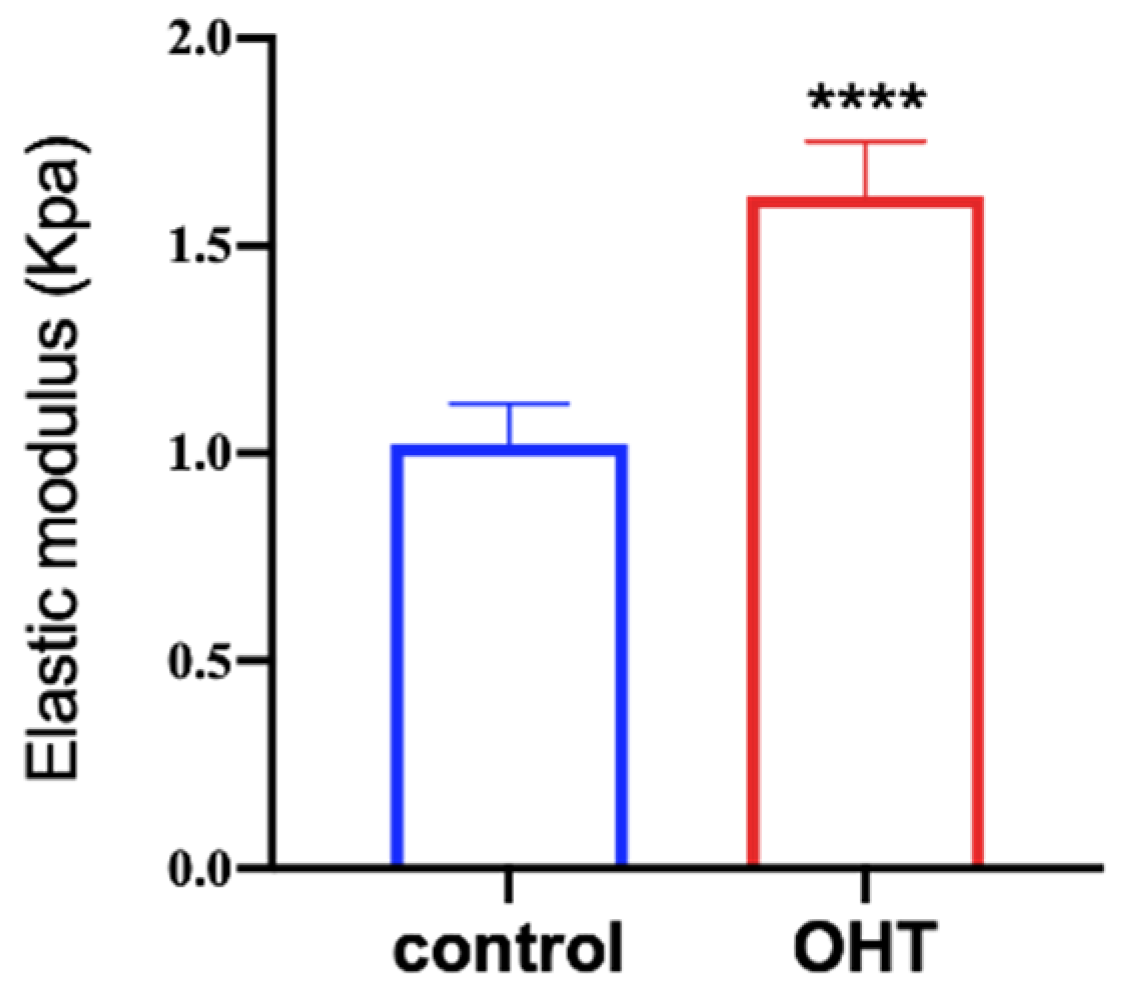
The AFM was used to measure the elastic moduli of TM cells. We randomly selected 30 cells from the OHT group and control group, respectively. Cells were pressed at the nuclei to measure their elastic moduli. The mean elastic moduli of TM cells in the control and OHT groups were 1.02 ± 0.10 KPa and 1.62 ± 0.13 KPa, respectively (as shown in Figure 5). The elastic moduli of TM cells were significantly higher in the OHT group than in the control group (p < 0.0001).
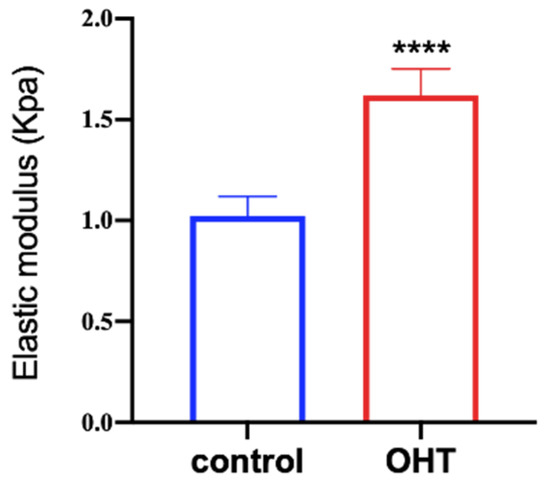
Figure 5.
The stiffness of the cells was tested by AFM. The Hertz equation was applied to fit the curve to obtain the elastic modulus. Elastic moduli of TM cells in the OHT group were greater than in the control group (**** p < 0.0001, n = 30).
3.5. The Expression of Vimentin in TM Cells
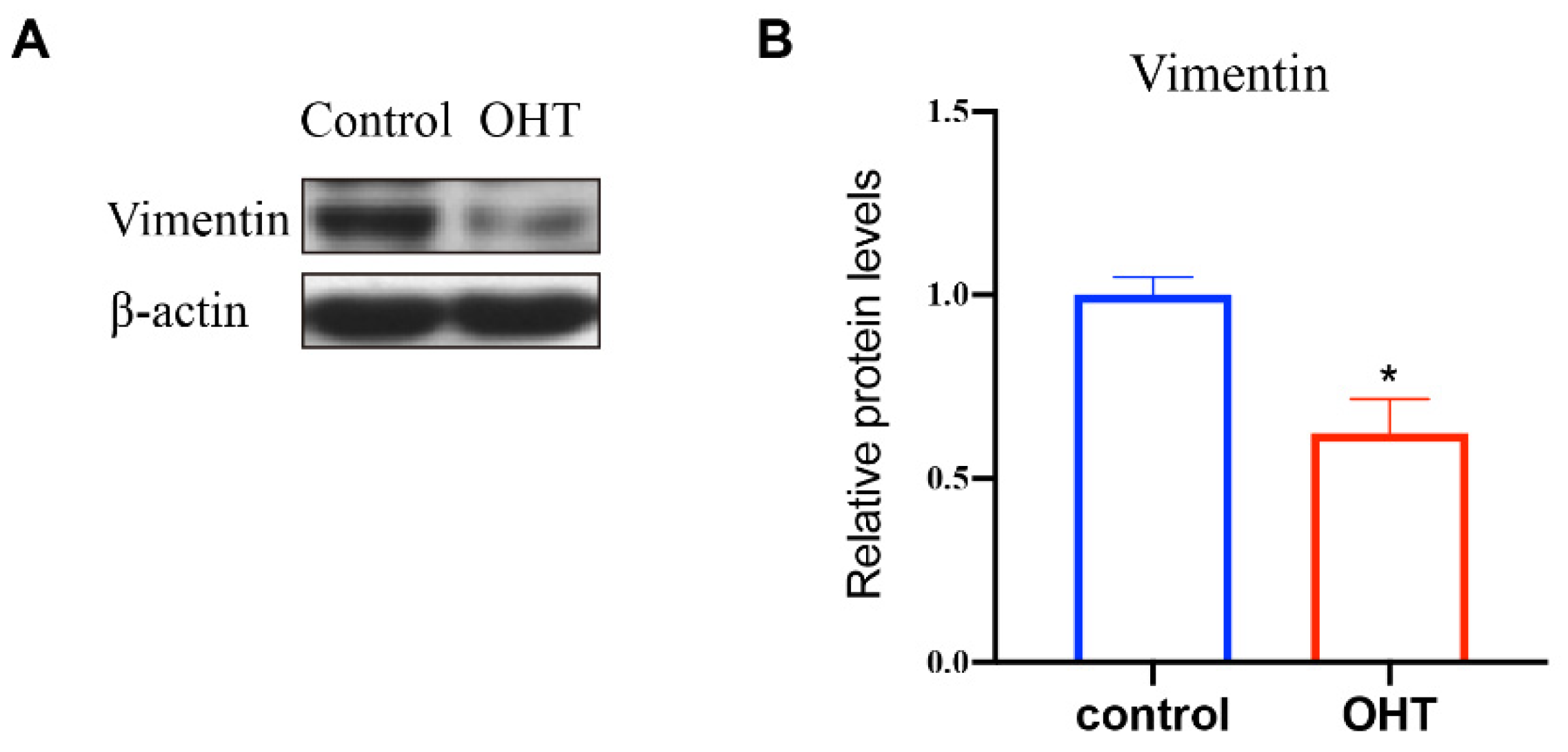
Vimentin is one of the cytoskeleton proteins in cells, which can confer mechanical flexibility to the cell [7]. In order to explore the effect of OHT on the expression of vimentin, western blot was used to detect the expression level of the proteins in the control and OHT groups. The bands and quantification analysis of vimentin expression, as shown in Figure 6, revealed that OHT significantly promoted the expression level of vimentin (p < 0.05) compared to the control group.
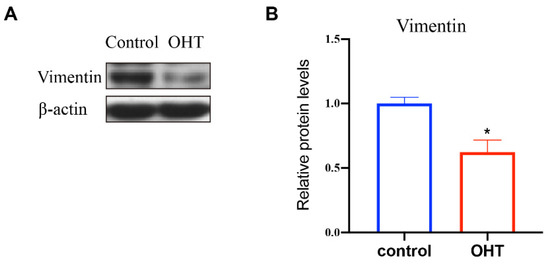
Figure 6.
Effects of OHT on the expression of vimentin in TM cells. (A) The bands of vimentin. Analysis of the bands was performed using Image J. (B) Quantification analysis of the expression level of vimentin. TM cells in the OHT group showed significantly decreased level of vimentin compared with control cells based on western blot analyses. * p < 0.05.
4. Discussion
Optic nerve head (ONH) damage and progressive visual field loss are common features of all types of glaucoma [14]. POAG is the most prevalent subtype of glaucoma. Pathological IOP elevation due to increased AH outflow resistance is a significant risk factor for POAG blindness, but the mechanism is still unclear. This research aimed to explain the effect of OHT on TM at the cellular level.
To reveal the effects of OHT on TM tissues and cells, researchers have applied stretch, compression, and shear stresses to normal TM cells cultured in vitro [25,26,27], or cultured the cells in 3D scaffolds [28,29]. However, this does not comprehensively mimic the actual mechanical stimuli effects in vivo, making it difficult to obtain a complete understanding of the TM cell response to biomechanical stimuli. There are also studies based on TM cells from glaucomatous patients, wherein the cells are affected by many factors such as age, medications, and the course of the disease [30], so there may be some limitations in using the TM cells of glaucomatous patients directly for research. Therefore, establishing a proper animal model is essential to investigating the biomechanical effects of OHT on the structure and function of TM cells. Due to the similarity in structures of human and rat TMs [31,32,33] and the damage of the laser method to the TM tissue, we established rat OHT models using the microsphere injection method [17,18] to investigate the mechanical properties, cytoskeletal changes, and protein expression of TM cells in this study. The average IOP of the experimental group remained above 24 mmHg from the 8th day to the end of the experiment. On the 22nd day after injection of the microspheres, the rats were executed, and TM cells were extracted for subsequent studies.
We observed that the actin fibers of the control TM cells were regularly arranged in the same direction. In contrast, the actin fiber arrangement of the TM cells in the OHT group was significantly changed. These rearranged fibers showed a disordered morphology. The cytoskeletal network’s structure is crucial for transmitting stresses and perceiving the mechanical microenvironment cues [34]. TM cells respond to mechanics by remodeling the cytoskeleton and extracellular matrix (ECM) pathways, thereby inhibiting their ability to drain AH [35]. Furthermore, we observed that OHT induced TM cells to exhibit affluent thin filamentous pseudopods and to form CLANs. In previous studies, CLAN formation has been found to lead to the suppression of TM cell functions, including proliferation, migration, and phagocytosis [36,37,38]. At the molecular level, changes in the cytoskeleton structure induce the activation of related proteins and pathways. For example, increased ECM stiffness (mimicking the glaucomatous TM matrix) induces YAP/TAZ activation by rearranging the cytoskeleton in TM cells [39], and YAP/TAZ is associated with increased TM cell fibrotic activity [40], which may lead to impaired AH outflow. We believe that the OHT-induced cytoskeletal rearrangement of TM cells is not only the result of OHT effects, but may also induce a series of intracellular mechanotransduction leading to a sustained increase in IOP.
Then, we measured the elastic moduli of TM cells using AFM. The results showed that the elastic modulus in the control group was 1.02 ± 0.10 KPa, which was consistent with the reported results [40]. In addition, our results demonstrated that OHT induced a significant increase in TM cell stiffness compared with the control cells. To the authors’ knowledge, there are few reports on changes in TM cell mechanical properties under the effect of OHT. Previous studies have revealed that mammalian cells rely on the cytoskeleton to modify cell shape, while the cytoskeletal structure is correlated with cellular mechanical properties [7,41,42,43]. Latrunculin B (Lat-B), a cytoskeletal disrupting agent, can obviously reduce the stiffness of human TM cells [44]. These results suggest that the OHT-induced increase in the elastic modulus of TM cells is probably due to the changes in cytoskeleton structure. Researchers have found that the mechanical properties of cells are closely linked to the cell function and development of disease [45,46]. The senescent human TM cells showed increased stiffness [47]. DEX-treated human TM cells showed increased elastic moduli and increased cellular fibrotic activity [40]. In addition, there is a relationship between cell stiffness and tissue stiffness [48]. Therefore, the OHT-induced increase in TM cell stiffness in this study may also indicate the increase in TM tissue stiffness. It was found that the elastic modulus of TM tissue in glaucomatous patients was about 20-fold higher than that of normal people [9]. Increased stiffness of the TM would cause higher resistance to AH outflow [49], causing IOP to be elevated. By mathematical modeling, the researchers also found that the resistance to flow increases as the TM elastic modulus increases [9].
The function of the cytoskeleton relies on its three main components, actin filaments, microtubules, and intermediate filaments (IFs). IFs are critical in establishing and maintaining the structural and mechanical properties of cells [7]. Vimentin, an abundant cytoskeletal protein, is a major member of the IF family [50]. We observed that OHT downregulated the vimentin expression level compared with the control group. Studies have demonstrated that decreased expression levels of vimentin lead to increased assembly and contractility of stress fibers [51]. In addition, several studies have shown that the expression of vimentin is related to cell stiffness [52,53,54,55]. Loss of vimentin increases contractile stress 3-fold [56]. Therefore, decreased expression of vimentin in TM cells may be one of the reasons for the increased cell stiffness induced by OHT. Actin filaments, microtubules, and IFs, as three distinct cytoskeletal filaments, are interconnected by protein–protein interactions, and if one of these filaments changes, the others are likely to change as well [57,58]. This interaction provides flexibility for the entire cytoskeleton, allowing it to be continually rearranged to fulfill the needs of the cell under different conditions [59]. Consequently, the change in vimentin expression in this study may be the cause of the OHT-induced cytoskeletal rearrangement. Since vimentin IFs are tightly connected to the cell nucleus, forming a cage-like network surrounding the nucleus [60,61], vimentin is a key protein that connects the actin cytoskeleton to the nucleus and transduces mechanical signals to the nucleus [7]. This study found that vimentin is sensitive to OHT, which may be a key component of TM mechanotransduction, and its detailed function deserves further investigation.
One limitation of this study is that we established an OHT model lasting only 2 weeks. In future studies, more animals can be involved to increase the OHT duration time points and explore the changes in TM cells’ mechanobiological characteristics with OHT duration time. The other limitation is that the TM cells were extracted from an OHT rat model, and then cultured and tested in vitro. The differences between cells in vivo and cultured in vitro need to be further explored. In addition, we can also extract cellular proteins or RNA for proteomic analyses or transcriptome sequencing analyses to investigate the biological response of TM cells from the perspective of overall protein/RNA changes, to further reveal OHT-induced mechanobiological changes in TM.
In summary, this study suggests that TM cells respond to OHT by rearranging the cytoskeletal structure, which may be caused by downregulating vimentin expression, thereby increasing the cell elastic modulus. This in turn may provide a basis for understanding the mechanobiological effects of OHT on TM cells.
Author Contributions
Conceptualization, Z.L., S.L. and L.L.; Funding acquisition, Z.L.; Investigation, S.H., Y.Y. and S.L.; Methodology, S.H., Y.Y. and S.L.; Writing—original draft, S.H.; Writing—review & editing, S.H., S.L. and L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by a grant from the National Natural Science Foundation of China (grant no. 31570952).
Institutional Review Board Statement
The animal study protocol was approved and monitored by the Institutional Animal Care and Use Committee of the Capital Medical University of Beijing. The approval code is AEEI-2017-085 and the approval date is August 2017.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of Interest
The authors declare that they have no conflict of interest to report regarding the present study.
References
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.M.; Tanna, A.P. Glaucoma. Med. Clin. N. Am. 2021, 105, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.F. The cell and molecular biology of glaucoma: Biomechanical factors in glaucoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2473–2475. [Google Scholar] [CrossRef]
- Zhu, W.; Hou, F.; Fang, J.; Fard, M.R.B.; Liu, Y.; Ren, S.; Wu, S.; Qi, Y.; Sui, S.; Read, A.T.; et al. The role of Piezo1 in conventional aqueous humor outflow dynamics. iScience 2021, 24, 102042. [Google Scholar] [CrossRef]
- Stamer, W.D.; Clark, A.F. The many faces of the trabecular meshwork cell. Exp. Eye Res. 2017, 158, 112–123. [Google Scholar] [CrossRef] [Green Version]
- Goel, M.; Pacciani, R.G.; Lee, R.K.; Battacharya, S.K. Aqueous humor dynamics: A review. Open Ophthalmol. J. 2010, 4, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patteson, A.E.; Carroll, R.J.; Iwamoto, D.V.; A Janmey, P. The vimentin cytoskeleton: When polymer physics meets cell biology. Phys. Biol. 2020, 18, 011001. [Google Scholar] [CrossRef]
- Humphrey, J.D.; Dufresne, E.R.; Schwartz, M.A. Mechanotransduction and extracellular matrix homeostasis. Nat. Rev. Mol. Cell Biol. 2014, 15, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Last, J.A.; Pan, T.; Ding, Y.; Reilly, C.M.; Keller, K.; Acott, T.S.; Fautsch, M.P.; Murphy, C.J.; Russell, P. Elastic modulus determination of normal and glaucomatous human trabecular meshwork. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2147–2152. [Google Scholar] [CrossRef] [PubMed]
- Read, A.T.; Chan, D.W.-H.; Ethier, C.R. Actin structure in the outflow tract of normal and glaucomatous eyes. Exp. Eye Res. 2007, 84, 214–226. [Google Scholar] [CrossRef]
- Dos Santos, Á.; Fili, N.; Pearson, D.S.; Hari-Gupta, Y.; Toseland, C.P. High-throughput mechanobiology: Force modulation of ensemble biochemical and cell-based assays. Biophys. J. 2021, 120, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-F.; Sun, Y.Y.; Peters, D.M.; Keller, K.E. The effects of mechanical stretch on integrins and filopodial-associated proteins in normal and glaucomatous trabecular meshwork cells. Front. Cell Dev. Biol. 2022, 10, 886706. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.J.; Zelenak, D. Experimental glaucoma in the primate induced by latex microspheres. J. Neurosci. Methods. 2001, 111, 39–48. [Google Scholar] [CrossRef]
- Pang, I.-H.; Clark, A.F. Inducible rodent models of glaucoma. Prog. Retin. Eye Res. 2020, 75, 100799. [Google Scholar] [CrossRef] [PubMed]
- Cone, F.E.; Gelman, S.E.; Son, J.L.; Pease, M.E.; Quigley, H.A. Differential susceptibility to experimental glaucoma among 3 mouse strains using bead and viscoelastic injection. Exp. Eye Res. 2010, 91, 415–424. [Google Scholar] [CrossRef] [Green Version]
- Rho, S.; Park, I.; Seong, G.J.; Lee, N.; Lee, C.K.; Hong, S.; Kim, C.Y. Chronic ocular hypertensive rat model using microbead injection: Comparison of polyurethane, polymethylmethacrylate, silica and polystyene microbeads. Curr. Eye Res. 2014, 39, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Samsel, P.A.; Kisiswa, L.; Erichsen, J.; Cross, S.D.; Morgan, J.E. A novel method for the induction of experimental glaucoma using magnetic microspheres. Investig. Ophthalmol. Vis. Sci. 2011, 52, 1671–1675. [Google Scholar] [CrossRef] [Green Version]
- Sappington, R.; Carlson, B.J.; Crish, S.D.; Calkins, D.J. The microbead occlusion model: A paradigm for induced ocular hypertension in rats and mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 207–216. [Google Scholar] [CrossRef]
- Mao, W.; Liu, Y.; Wordinger, R.J.; Clark, A.F. A magnetic bead-based method for mouse trabecular meshwork cell isolation. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3600–3606. [Google Scholar] [CrossRef] [Green Version]
- Eastlake, K.; Jayaram, H.; Luis, J.; Hayes, M.; Khaw, P.T.; Limb, G.A. Strain specific responses in a microbead rat model of experimental glaucoma. Curr. Eye Res. 2021, 46, 387–397. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Kanamori, A.; Nakamura, M.; Negi, A. Rat chronic glaucoma model induced by intracameral injection of microbeads suspended in sodium sulfate-sodium hyaluronate. Jpn J. Ophthalmol. 2014, 58, 290–297. [Google Scholar] [CrossRef]
- Ge, P.; Navarro, I.D.; Kessler, M.M.; Bernier, S.G.; Perl, N.R.; Sarno, R.; Masferrer, J.; Hannig, G.; Stamer, W.D. The soluble guanylate cyclase stimulator IWP-953 increases conventional outflow facility in mouse eyes. Investig. Ophthalmol. Vis. Sci. 2016, 57, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Khaw, P.T.; Occleston, N.L.; Schultz, G.; Grierson, I.; Sherwood, M.B.; Larkin, G. Activation and suppression of fibroblast function. Eye 1994, 8 Pt 2, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, L.; Liu, Z. Experimental research on the relationship between the stiffness and the expressions of fibronectin proteins and adaptor proteins of rat trabecular meshwork cells. BMC Ophthalmol. 2017, 17, 268. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Utsunomiya, T.; Ishibazawa, A.; Song, Y.-S.; Udo, M.S.B.; Tasaki, Y.; Yoshida, A. Extracellular matrix gene expression in human trabecular meshwork cells following mechanical fluid flow stimulation. Int. J. Ophthalmol. 2022, 15, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Shim, M.S.; Nettesheim, A.; Hirt, J.; Liton, P.B. The autophagic protein LC3 translocates to the nucleus and localizes in the nucleolus associated to NUFIP1 in response to cyclic mechanical stress. Autophagy 2020, 16, 1248–1261. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, S.; Tan, Y.; Wang, Y. Effects of mechanical compression on cell morphology and function in human corneal fibroblasts. Curr. Eye Res. 2021, 46, 1467–1473. [Google Scholar] [CrossRef] [PubMed]
- Tirendi, S.; Saccà, S.C.; Vernazza, S.; Traverso, C.; Bassi, A.M.; Izzotti, A. A 3D model of human trabecular meshwork for the research study of glaucoma. Front. Neurol. 2020, 11, 591776. [Google Scholar] [CrossRef]
- Osmond, M.; Bernier, S.M.; Pantcheva, M.B.; Krebs, M.D. Collagen and collagen-chondroitin sulfate scaffolds with uniaxially aligned pores for the biomimetic, three dimensional culture of trabecular meshwork cells. Biotechnol. Bioeng. 2017, 114, 915–923. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Read, A.T.; Sulchek, T.; Ethier, C.R. Trabecular meshwork stiffness in glaucoma. Exp. Eye Res. 2017, 158, 3–12. [Google Scholar] [CrossRef]
- Smith, R.; Sundberg, J.; John, S. The anterior segment and ocular adnexae. In Systematic Evaluation of the Mouse Eye; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Cameron, M.J.; Clark, A.F.; Pang, I.H. Assessment of aqueous humor dynamics in the mouse by a novel method of constant-flow infusion. Investig. Ophthalmol. Vis. Sci. 2011, 52, 685. [Google Scholar]
- Millar, J.C.; Pang, I.H. Non-continuous measurement of intraocular pressure in laboratory animals. Exp. Eye Res. 2015, 141, 74–90. [Google Scholar] [CrossRef] [PubMed]
- Janmey, P.A.; McCulloch, C.A. Cell mechanics: Integrating cell responses to mechanical stimuli. Annu. Rev. Biomed. Eng. 2007, 9, 1–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overby, D.R.; Bertrand, J.; Schicht, M.; Paulsen, F.; Stamer, W.D.; Lütjen-Drecoll, E. The structure of the trabecular meshwork, its connections to the ciliary muscle, and the effect of pilocarpine on outflow facility in mice. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3727–3736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, A.F.; Brotchie, D.; Read, A.T.; Hellberg, P.; English-Wright, S.; Pang, I.-H.; Ethier, C.R.; Grierson, I. Dexamethasone alters F-actin architecture and promotes cross-linked actin network formation in human trabecular meshwork tissue. Cell Motil Cytoskelet. 2005, 60, 83–95. [Google Scholar] [CrossRef]
- Zhang, X.; Ognibene, C.M.; Clark, A.F.; Yorio, T. Dexamethasone inhibition of trabecular meshwork cell phagocytosis and its modulation by glucocorticoid receptor beta. Exp. Eye Res. 2007, 84, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, T.; Inoue, T.; Inoue-Mochita, M.; Tanihara, H. Live cell imaging of actin dynamics in dexamethasone-treated porcine trabecular meshwork cells. Exp. Eye Res. 2016, 145, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Raghunathan, V.; Stamer, W.D.; Ganapathy, P.S.; Herberg, S. Extracellular matrix stiffness and TGFβ2 regulate YAP/TAZ activity in human trabecular meshwork cells. Front. Cell Dev. Biol. 2022, 10, 844342. [Google Scholar] [CrossRef]
- Liu, Z.; Li, S.; Qian, X.; Li, L.; Zhang, H.; Liu, Z. RhoA/ROCK-YAP/TAZ axis regulates the fibrotic activity in dexamethasone-treated human trabecular meshwork cells. Front. Mol. Biosci. 2021, 8, 728932. [Google Scholar] [CrossRef]
- Uray, I.P.; Uray, K. Mechanotransduction at the plasma membrane-cytoskeleton interface. Int. J. Mol. Sci. 2021, 22, 11566. [Google Scholar] [CrossRef]
- Pegoraro, A.F.; Janmey, P.; Weitz, D.A. Mechanical Properties of the Cytoskeleton and Cells. Cold Spring Harb. Perspect. Biol. 2017, 9, a022038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; McNally, S.; Kilpatrick, J.I.; Jarvis, S.P.; O’Brien, C.J. Aging and ocular tissue stiffness in glaucoma. Surv. Ophthalmol. 2018, 63, 56–74. [Google Scholar] [CrossRef]
- McKee, C.T.; Wood, J.A.; Shah, N.M.; Fischer, M.E.; Reilly, C.M.; Murphy, C.J.; Russell, P. The effect of biophysical attributes of the ocular trabecular meshwork associated with glaucoma on the cell response to therapeutic agents. Biomaterials 2011, 32, 2417–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katta, S.; Krieg, M.; Goodman, M.B. Feeling force: Physical and physiological principles enabling sensory mechanotransduction. Annu. Rev. Cell Dev. Biol. 2015, 31, 347–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eyckmans, J.; Boudou, T.; Yu, X.; Chen, C.S. A hitchhiker’s guide to mechanobiology. Dev. Cell 2011, 21, 35–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, J.T.; Raghunathan, V.K.; Chang, Y.-R.; Murphy, C.J.; Russell, P. The intrinsic stiffness of human trabecular meshwork cells increases with senescence. Oncotarget 2015, 6, 15362–15374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tee, S.Y.; Fu, J.; Chen, C.S.; Janmey, P.A. Cell shape and substrate rigidity both regulate cell stiffness. Biophys. J. 2011, 100, L25–L27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vranka, J.A.; Staverosky, J.A.; Reddy, A.P.; Wilmarth, P.A.; David, L.L.; Acott, T.S.; Russell, P.; Raghunathan, V.K. Biomechanical rigidity and quantitative proteomics analysis of segmental regions of the trabecular meshwork at physiologic and elevated pressures. Investig. Ophthalmol. Vis. Sci. 2018, 59, 246–259. [Google Scholar] [CrossRef] [Green Version]
- Schaedel, L.; Lorenz, C.; Schepers, A.V.; Klumpp, S.; Köster, S. Vimentin intermediate filaments stabilize dynamic microtubules by direct interactions. Nat. Commun. 2021, 12, 3799. [Google Scholar] [CrossRef]
- Jiu, Y.; Peränen, J.; Schaible, N.; Cheng, F.; Eriksson, J.E.; Krishnan, R.; Lappalainen, P. Vimentin intermediate filaments control actin stress fiber assembly through GEF-H1 and RhoA. J. Cell Sci. 2017, 130, 892–902. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Yin, L.; Song, X.; Yang, H.; Ren, X.; Gong, X.; Wang, F.; Yang, L. Effects of vimentin disruption on the mechanoresponses of articular chondrocyte. Biochem. Biophys. Res. Commun. 2016, 469, 132–137. [Google Scholar] [CrossRef]
- Sharma, P.; Bolten, Z.T.; Wagner, D.R.; Hsieh, A.H. Deformability of human mesenchymal stem cells is dependent on vimentin intermediate filaments. Ann. Biomed. Eng. 2017, 45, 1365–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gladilin, E.; Gonzalez, P.; Eils, R. Dissecting the contribution of actin and vimentin intermediate filaments to mechanical phenotype of suspended cells using high-throughput deformability measurements and computational modeling. J. Biomech. 2014, 47, 2598–2605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathje, L.-S.Z.; Nordgren, N.; Pettersson, T.; Rönnlund, D.; Widengren, J.; Aspenström, P.; Gad, A.K.B. Oncogenes induce a vimentin filament collapse mediated by HDAC6 that is linked to cell stiffness. Proc. Natl. Acad. Sci. USA 2014, 111, 1515–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Loosdregt, I.A.E.W.; Weissenberger, G.; van Maris, M.P.F.H.L.; Oomens, C.W.J.; Loerakker, S.; Stassen, O.M.J.A.; Bouten, C.V.C. The mechanical contribution of vimentin to cellular stress generation. J. Biomech. Eng. 2018, 140, 061006. [Google Scholar] [CrossRef] [PubMed]
- Goldman, R.D.; Cleland, M.M.; Murthy, S.P.; Mahammad, S.; Kuczmarski, E.R. Inroads into the structure and function of intermediate filament networks. J. Struct. Biol. 2012, 177, 14–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollenbeck, P.J.; Bershadsky, A.D.; Pletjushkina, O.Y.; Tint, I.S.; Vasiliev, J.M. Intermediate filament collapse is an ATP-dependent and actin-dependent process. J. Cell Sci. 1989, 92 Pt 4, 621–631. [Google Scholar] [CrossRef]
- Danielsson, F.; Peterson, M.K.; Araújo, H.C.; Lautenschläger, F.; Gad, A.K.B. Vimentin diversity in health and disease. Cells. 2018, 7, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patteson, A.E.; Vahabikashi, A.; Pogoda, K.; Adam, S.A.; Mandal, K.; Kittisopikul, M.; Sivagurunathan, S.; Goldman, A.; Goldman, R.D.; Janmey, P.A. Vimentin protects cells against nuclear rupture and DNA damage during migration. J. Cell Biol. 2019, 218, 4079–4092. [Google Scholar] [CrossRef] [PubMed]
- Goldman, R.D.; Grin, B.; Mendez, M.G.; Kuczmarski, E.R. Intermediate filaments: Versatile building blocks of cell structure. Curr. Opin. Cell Biol. 2008, 20, 28–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).