Abstract
Microbial biofilms have co-evolved with grazing animals, such as gastropods, to develop mutually beneficial relationships. Although microbial biofilms demonstrate resilience and resistance to chemical exposure, pre-existing relationships can be negatively affected by chemical input. In this study, we determined how the grazer, Littorina littorea (common periwinkle sea snail), and a biological surfactant (rhamnolipid) interact on a phototrophic marine biofilm. Biofilms were cultured in 32 twenty-liter buckets at the Queen’s University Marine Laboratory in Portaferry, Northern Ireland on clay tiles that were either exposed to 150 ppm of a rhamnolipid solution or that had no chemical exposure. L. littorea were added into half of the buckets, and biofilms were developed over 14 days. Biofilms exposed to grazing alone demonstrated high tolerance to the disturbance, while those growing on rhamnolipid-exposed substrate demonstrated resistance but experienced slight declines in carbon and stoichiometric ratios. However, when exposed to both, biofilms had significant decreases in stoichiometry and declined in productivity and respiration. This is problematic, as continuing marine pollution increases the likelihood that biofilms will be exposed to combinations of stressors and disturbances. Loss of biofilm productivity within these areas could lead to the loss of an important food source and nutrient cycler within the marine ecosystem.
1. Introduction
Microorganisms, such as those making up microbial biofilms, are major drivers of biogeochemical cycling (or nutrient movement across abiotic and biotic elements) within aquatic ecosystems [1,2,3,4]. In marine environments, these biofilms are often made up of bacteria, which are important cyclers of dissolved organic matter [5,6], and diatoms, depending on their location [7]. Diatoms can break down and recycle environmental nutrients, such as nitrogen, which is necessary in typically nitrogen-limited marine environments [8,9]. Diatoms are also very important autotrophs, acting as major primary producers in marine environments [10,11].
Living within biofilms is a way that microorganisms can protect themselves against outside biotic and abiotic stressors and disturbance events, including environmental changes or contaminant exposure [12]. The surrounding extracellular polymeric substance (EPS) matrix, formed by microorganisms within the biofilm [13], limits compounds, such as antibiotics, from reaching the bacterial cells and other microorganisms within the biofilm [14,15,16]. This matrix has a high carbon content and is mainly composed of compounds such as proteins, lipids, polysaccharides, and extracellular DNA [17]. The structure of the EPS matrix itself also provides biofilms a measure of protection against predation and grazing. For example, cellulose in the EPS matrix gives biofilms resistance against the mechanical forces of predation from Caenorhabditis elegans (nematode) [18].
Biofilms have complex relationships with other aquatic organisms. For many, such as predatory bacteria [19], nematodes [20], invertebrates, and various fish taxa [21], the biofilms are an important food source. However, they also have holobiontic (where a symbiotic relationship forms between hosts and the surrounding microbiotas, creating a unique ecosystem [22]) and mutualistic relationships with other marine organisms. Holobiontic relationships have been noted among grazers where snails [23] and limpets [24] remove unproductive canopy from the biofilm. This encourages new biofilm growth and greater productivity, leading to benefits to both the grazer, which also acts as the host for microorganisms, and the surrounding biofilm itself. Research has shown that the presence of filter feeders, such as mussels and tadpoles, in aquatic environments can increase benthic primary production and increase nitrogen (N): phosphorus (P) ratios within aquatic biofilms, leading to a bottom-up effect on the food chain from consumers [25,26]. These beneficial relationships are important for local biofilm and grazer health, and any alterations in biofilm stoichiometry (determined through testing carbon (C) and N within the biofilm that indicate changes in the EPS matrix and nutrient levels) potentially indicate shifts in biofilm productivity and composition.
Although organisms such as snails can positively influence biofilm and EPS accumulation, microorganisms within a biofilm typically produce biological surfactants, such as rhamnolipids, as a way to control their EPS matrix size and structure [27,28]. Surfactants are part of a chemical class that reduces surface tension through micelle formation. Micelles occur when molecules form an aggregate where hydrophobic tails or chains are on the inside [29] and the hydrophilic moieties (or heads) of the amphiphilic compound are on the outside [30]. Biological surfactants, such as rhamnolipids, are currently being investigated for their ability to control biofilm growth industrially and medically in a more environmentally friendly way than synthetic alternatives, which have a known toxicity within the environment [31]. With a growing presence of anthropogenic chemicals within marine environments [32], marine biotas are constantly exposed to low levels of these pollutants. Surfactants specifically are being introduced in great diversity and quantity to the marine environment due to their heavy use in cargo ships, oil cleanups, and ocean drilling [32]. This can be problematic, as many biotas, such as biofilms within a holobiontic relationship, evolved to resist and be tolerant to either competition, stress, or disturbance (C-S-R survival strategies where R is for ruderal (i.e., disturbed lands)) [33,34]. When biofilms that are resistant to stressors, such as grazers, are exposed to chemical disturbances, they may lose the beneficial relationship from grazers that allowed them greater productivity and faster growth rates. Historically, research identifying potential effects from biological surfactant exposure has mostly examined how well biological surfactants compare to synthetic surfactants for biofilm control in medical and industrial environments but has expended much less effort on investigating how biological surfactants may affect desirable aquatic biofilms and biological interactions within them. With a rising trend of replacing synthetic surfactant use by biological surfactants across all areas of application, it is more important than ever to understand the risk applications that biological surfactants pose to important marine biotas, such as biofilms, as the foundation of many benthic food webs.
In this study, we investigated the individual and combined effects of grazers and biosurfactants upon marine biofilms. Using a flow-through mesocosm, we tested how the metabolic activity and structure of a biofilm changed following grazer (Littorina littorea (common periwinkle sea snail)) exposure and its growth on a substrate exposed to a biological surfactant, rhamnolipid, together and separately. We hypothesized that biofilms would have a positive relationship with grazers, and productivity would increase overall. As rhamnolipids are used for biofilm control, we expected that the tested biofilms would be unable to grow on the exposed substrate. The addition of snails and rhamnolipid influence on a biofilm would alter metabolic activity and structure, as the EPS matrix and biofilm itself will be under added pressures. We tested these hypotheses by quantifying changes in biofilm gross primary productivity (GPP), community respiration (CR), biomass, and C:N ratios.
2. Materials and Methods
2.1. Experimental Set Up
Thirty-two outdoor flow-through marine mesocosms were set up at the Queen’s University Marine Laboratory in Portaferry, Northern Ireland to identify biofilm response to exposure from both a grazer (Littorina littorea, common periwinkle sea snail) and a rhamnolipid biosurfactant (JBR 425, Jeneil Biosurfactant Company, Saukville, WI, USA). L. littorea was the grazer chosen for this experiment, as it is often used in marine toxicology experiments and can be found along the Strangford Lough coastline. Due to space limitations, only one biosurfactant was used in order to maximize the number of replicates and robustness within the study. Rhamnolipids were chosen as our biosurfactant due to their potential application as an alternative biofilm control agent to chemical surfactants. Mesocosms were contained within a shallow table receiving a constant flow of sand-filtered seawater pumped directly from the adjacent Strangford Lough. Each mesocosm was one 20 L bucket, with a hose for seawater inflow and a hole approximately 11 cm above the base for water outflow. In total, four variables were tested (control with no grazers or surfactant, grazer exposure only, surfactant exposure only, and combined) with eight replicates per variable.
L. littorea (n = 80) were collected by hand from the Strangford Lough coastline and transported in sea water to the laboratory within one hour after collection. Only snails with an observable length > 2 cm were chosen, as smaller snails may not have an observable effect on biofilm biomass over a two-week period. A 150 ppm solution of rhamnolipid was created in a large bucket. Thirty-two clay tiles (L10 × W10 × H0.7 cm) were dosed in the solution for five minutes before being allowed to thoroughly air dry on a sterile table. Afterwards, two dosed clay tiles were placed flat on the bottom of sixteen of the mesocosms within a mesh bag. The remaining sixteen mesocosms received two tiles without chemical additions within a mesh bag. Tiles were used as substrata for marine biofilm growth during the experiment. Eight mesocosms containing rhamnolipid-dosed tiles and eight mesocosms without chemically dosed tiles received snails (n = 5) within their mesh bags. Bags were tightened to prevent snail loss, and snails were initially placed directly on the tiles. Water quality parameters (pH, dissolved oxygen (DO) (mg/L) and water temperature (°C) were measured daily, and snail loss was monitored. The experiment began on 28 July 2020 and lasted a period of 14 days.
2.2. Sample Collection
On day 14 the water flow to the buckets was stopped. Snails were initially removed from the mesh bags and counted. While minimal snail losses occurred, the losses were comparable among treatments. DO measurements (mg/L) (HQ40D multi-meter probe, Hach Lange GmbH, Düsseldorf Germany) were taken from each mesocosm and recorded. To obtain respiration and primary productivity data, oxygen measurements were necessary from the mesocosms in dark (night) and light (day) environments. To simulate a nighttime environment, dark plastic bags were placed over each mesocosm to prevent light from reaching any biofilms that had developed on the clay tiles. Bags were pressed against the water line to prevent excess oxygen from entering the water as CR commenced. Incubation occurred for 65 min, after which oxygen measurements were re-measured to determine DO loss over that period of time. Mesocosms were then covered with clear plastic to allow natural light penetration but no oxygen penetration. This allowed us to measure the productivity of the biofilms alone and not measure excess oxygen input from the outside. Incubations lasted for 65 min, after which the DO measurements were re-taken to obtain GPP.
After obtaining oxygen measurements, tiles were carefully removed from their respective mesocosms. Razor blades were cleaned with 70% ethanol for bacteriostatic effect and used to scrape biofilm material into sterile, pre-weighed, 2 mL Eppendorf tubes. Obtained biofilm materials were immediately weighed and frozen at −20 °C to ensure the contents were fully frozen and solidified. The materials were then lyophilized (−85 °C (Alpha 2-4 LDplus, Martin Christ Freeze Dryers, Osterode am Harz, Germany)) and sent to the Agri-Food and Biosciences Institute (Belfast, Northern Ireland) for C (% C) and N (% N) content analyses.
The C and N content of lyophilized biofilm samples were determined by combustion elemental analysis on a Thermo Flash 2000 elemental analyzer (Thermo Fisher Scientific GmbH, Bremen, Germany). Briefly, biofilm samples were weighed and encapsulated in pre-weighed tin cups for analysis. Each sample was then combusted at 950 °C within a 10 sec stream of ultra-pure oxygen (flow rate 240 mL/min). Exothermic oxidation of the tin cup raised the combustion temperature to over 1000 °C. The gaseous carbon and nitrogen oxides from each sample were carried in a stream of helium carrier gas (140 mL/min) through a column of cobaltous oxide catalyst to remove sulfur and halogenated compounds. The purified gas then passed through a column of reduced copper, held at 680 °C, which converted the nitrous oxide gases present to elemental nitrogen and retained excess oxygen. Any water vapor present was removed from the gas mixture by passage through a magnesium perchlorate-packed adsorption filter, after which the carbon and nitrogen were separated on a chromatographic column and passed to a thermal conductivity detector. The instrument was calibrated following the manufacturer’s instructions, using the K-factor calibration methodology with acetanilide as an analytical standard. Aspartic acid (Thermo Fisher part number: 338 400 22) and a commercial soil reference material (Thermo Fisher part number 334 400 25) were used as quality control (QC) materials to check the instrument calibration, with two replicates of each QC material analyzed within each batch of 30 samples. The ratios of C to N were examined to identify changes in ecological stoichiometry [35]. Lyophilized weights were used for biomass analysis. The mass of carbon and nitrogen within a sample was then calculated as:
where S (C or N) is the mass of carbon or nitrogen within the sample, S (dry weight) is the dry weight of the sample, and P (C or N) is the percentage carbon or percentage nitrogen in the sub-sample analyzed by the elemental analyzer.
S (C or N) = S (dry weight) × (P_(C or N)/100)
2.3. Data Analysis
R version 4.0.3 [36] was used for all statistical analyses. Additional R packages were employed for normality testing (‘car’ [37]), figure creation (‘ggplot2′ [38]), and non-parametric testing (‘FSA’ [39]). All GPP and CR data met assumptions of normality and homogeneity of variance, while biomass and C:N data were not normal, and therefore, non parametric tests were used. The Shapiro–Wilk’s test was performed to measure normality, and Levene’s test for equality of variances was used to confirm normality. GPP and CR data were calculated to mg O2/cm2 biofilm, and C:N data were transformed from % to mass (mg C/N per mg biofilm) before further analyses were performed. To test for differences among the four treatments, a one-way analysis of variance (ANOVA) test was carried out. A Kruskal–Wallis test followed by a Dunn’s multiple comparison post hoc test was used to test for significant differences within both the biomass and the C:N data.
3. Results
3.1. Primary Productivity, Respiration, and Biomass
We found no significant differences between treatments (control, grazer exposure, chemical exposure, and combined) in GPP (ANOVA F = 0.46, p = 0.712) and CR (ANOVA F = 1.082, p = 0.373) (Figure 1A,B). There were non-significant decreases in GPP and CR within the rhamnolipid treatment and the combined treatment. The lyophilized biomass also had no significant differences (Kruskal-Wallis p = 0.669); however, there were slight decreases in the average control biomass compared to all other treatments (control (9.71 ± 2.07), grazer only (10.72 ± 0.64), rhamnolipid (10.32 ± 1.16), and combined (10.49 ± 0.33)).
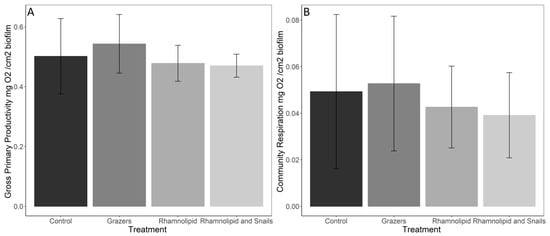
Figure 1.
Gross primary productivity (A) and community respiration (B) of marine microbial biofilms exposed to either predation by the common periwinkle snail and/or 150 ppm rhamnolipid for 14 days. N = 8 ± SD.
3.2. Biofilm Stoichiometry
Significant differences were found among the control, rhamnolipid-treated, and combination biofilms for C:N ratios by mass (Kruskal–Wallis, p = 0.0121) (Figure 2). Dunn’s post hoc test showed that these differences lay between the mesocosms containing the combination treatment and the surfactant-only treatment (Dunn p = 0.0171), and the combination treatment and the control (Dunn p = 0.0018). Both treatments with no snails had significantly lower C:N ratios than the rhamnolipid treatment with snails. Significant differences were also identified in the quantity of carbon present within the biofilms (mg C/mg biofilm) (Kruskal–Wallis p = 0.0139) (Figure 3A). The treatment containing both rhamnolipid-dosed tiles and snails had significantly less C than both non-chemically treated systems (with snails: Dunn p = 0.00126, without snails: Dunn p = 0.0456). There were no significant differences in N content within the biofilms among any treatment or control (Kruskal–Wallis p = 0.222) (Figure 3B).
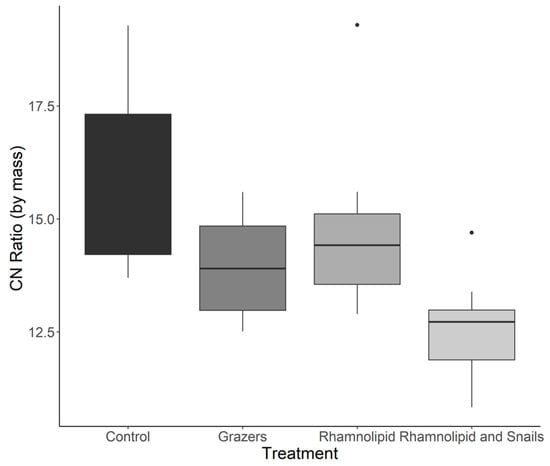
Figure 2.
Carbon: nitrogen ratios of marine microbial biofilms (n = 8) exposed to predation by either the common periwinkle snail and/or 150 ppm rhamnolipid for 14 days. Dots indicate outliers in the data for figure visualization.
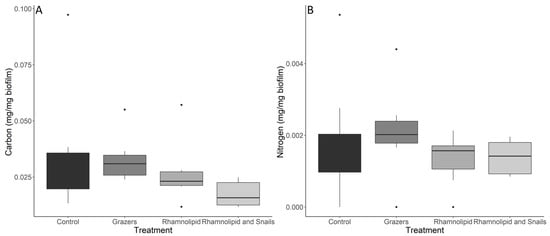
Figure 3.
Carbon (A) and nitrogen (B) quantities within marine microbial biofilms (n = 8) exposed to either predation by the common periwinkle snail and/or 150 ppm rhamnolipid. Dots indicate outliers in the data for figure visualization. All mesocosms were treated similarly and were thus equally affected by the low-level deposition of inorganic solids from the sea water flowing through the system. This deposition decreased the relative carbon and nitrogen contents within the measured biofilm, which were analyzed after a 14-day exposure.
4. Discussion
Our experiment demonstrates that marine biofilms are highly resilient to stress and disturbance events but have lower tolerances of them when experiencing both together. Productivity was not altered within our experimental timeline, but resultant stoichiometric shifts may lead to less productive biofilms over a longer timescale, affecting local food sources and biogeochemical cycling. Although past studies have focused on how biofilm metabolic activity and structure change after exposure to biosurfactants and grazers separately, no research has been performed to investigate these combined relationships.
Prior research has shown that biofilm exposure to synthetic surfactants can decrease respiratory activity temporarily, with full recovery occurring after 12 h [40]. Conversely, antibiotic exposure temporarily increased respiration for several hours for affected single-species biofilms [41]. Biomass has similarly been negatively affected by surfactant exposure over a short time period. Rhamnolipid exposure reduced single-species biofilms by 74–98% over a 4-day exposure of a 4-day-old biofilm [42]. Grazing on biofilms from known biofilm feeders has also been seen previously to significantly decrease biofilm biomass after a period of 3–4 days [43]. Our study took place over a longer time period than the previous studies, and the exposure started before the initial growth occurred. Therefore, it is likely that the lack of significant differences in GPP, CR, and biomass occurred due to the longer experimental timeline allowing the biofilm time to recover and adapt its growth to the presence of a surfactant stressor. Our GPP and CR results are also comparable to other studies examining the GPP and CR of biofilms overall, demonstrating a similar productivity level to other aquatic biofilms [44,45]. In addition, our results were comparable to another long-term study, where a 32-day exposure of rhamnolipid yielded no differences in examined GPP, CR, or biomass for exposed freshwater biofilms, where exposure to the surfactant also occurred before biofilm development began [46].
As originally hypothesized, exposed biofilms demonstrated strong tolerance and resistance to grazing effects. Our results also indicate the development of a potentially holobiontic relationship, as biomass increased and productivity remained high and similar to the control per cm2 of biofilm material examined. The formation of a beneficial relationship between biofilms and grazers in our study is supported by previous studies where similar results occurred, although we obtained our evidence over a much smaller timeframe. Limpet grazing of epilithic biofilms over the summer when growth and grazing rates peaked led to biofilm biomass growth increases of 20% later on in the season [47]. Nutrients from grazer excretion can also specifically increase microphytobenthic growth, which would then affect local biofilm productivity [48]. Nutrients obtained from grazer excretion are typically P [49] and N [50], which would explain why biofilms exposed to only grazers within our mesocosms had altered stoichiometry with decreased C:N ratios and increased N. Relatively low C and N contents in percent of biofilm dry mass were attributable to the inevitable low-level deposition of inorganic solids from the water flowing through the system. Therefore, in line with previous research, our results show an indication that marine biofilms, even after 14 days of grazing, may be able to develop beneficial relationships with grazers and demonstrate high tolerance for grazing disturbances.
Although biofilms can slightly resist chemical exposure through their EPS matrix (Pinto et al. 2020), rhamnolipids are known for their ability to break down biofilms [51], and therefore, we did not expect growth and development of biofilm material to occur on the exposed substrate at a comparable level to the control. Although not significant, loss of C within our biofilms in the rhamnolipid-treated systems led to a decreased C:N ratio. Rhamnolipids can encourage EPS material disintegration [52]. It is likely that, as the EPS matrix of a biofilm is heavily made up of lipids, polysaccharides, proteins, and extracellular DNA [17], and therefore heavy in C content, rhamnolipids affected the EPS matrix and decreased overall C within the biofilm. Due to the loss of C and altered biofilm stoichiometry within rhamnolipid-treated mesocosms, biofilms were not wholly tolerant of the chemical but demonstrated strong resistance, as productivity did not change. There is a chance that resistance developed, as biofilm exposure to rhamnolipid was diluted with the constant input of fresh seawater; however, a constant exposure study would have to be performed to confirm this possibility. Carrying out additional measurements of macromolecular compositions of the biofilm would be a useful undertaking in similar future studies.
The marine environment contains a combination of stressors and disturbances for biofilms and other biotas. As many of the stressors, such as chemical exposure, are fairly new, we hypothesized that our biofilms would not be tolerant to a combination of grazing and chemical exposure. Although productivity did not change within exposed biofilms, stoichiometry did, with a significant decrease in C:N in the presence of rhamnolipid, indicating resistance but not full tolerance. Our results support the idea that organisms will have different, and potentially negative, reactions to new combinations of stressors and disturbances within the environment. Climate change, leading to increased temperature stress, led to decreases in grazing overall from L. littorea after a two-week period [53]. Grazing habits are also based on food availability, which can be impacted by stressors, such as pollution and flow alteration [54]. As N increased in mesocosms exposed to grazing alone due to excretion, likely of ammonium by L. littorea, the lack of differences in N after dual exposure may demonstrate a lack of grazing occurring, thereby indicating damage to a previously beneficial relationship. Prior research has also shown that when the stressor is a chemical, such as atrazine, the combination of grazing and chemical pressure decreased carbon uptake and biofilm density overall, even though when examining the stressor alone, no differences were identified. Grazing pressure reduced the chances of biofilms developing new materials that are resistant to atrazine, thereby preventing recovery [55]. This is very problematic, as biofilms can develop chemical resistance overtime in areas with high chemical stress [56], but this cannot happen if the grazing of new, resistant material occurs. Although we did not perform an analysis to identify changes in chemical resistance, the significant reduction of C and C:N indicate stoichiometric, and likely structural, shifts after dual exposure. Comparing these results to those identified within our disturbance only mesocosms provides strong evidence to suggest that dual exposure has detrimental effects on biofilms that may otherwise be tolerant to individual disturbances or resistant to stressors.
Our experiment supports the idea that biofilms can develop holobiontic relationships with grazers, and that they may be sensitive to combined stress and disturbance events. Although there were decreases in the overall C:N ratios present in grazer-disturbed mesocosms, the decreases likely occurred from snail ammonium excretion increasing N within the system. As productivity was stable but biomass increased, biofilms were not negatively affected by the presence of grazers alone and experienced increases in growth. The biofilm grown on rhamnolipid-treated substrate alone also experienced minimal differences with productivity, demonstrating high resistance, but had decreases in C, likely from the negative effects of chemical exposure on the EPS matrix. Although biofilms from the rhamnolipid treatments demonstrated resistance and showed stable productivity, they were not fully tolerant, which could lead to a productivity decline after further exposure. These two events demonstrate the natural tolerance and resistance to individual stressor and disturbance events that mixed marine biofilms have. When exposed to both grazing and chemicals, biofilms were less productive overall, had large stoichiometry changes, and were grazed on less. Overall, exposure to a combination of stressors and disturbances commonly seen in the marine environment negatively impacted both biofilms and grazers that rely on biofilms as a food source. This can have negative consequences throughout the local ecosystem, as the disruption of local biofilm relationships that previously increased biofilm biomass and productivity overtime means the disruption of grazing habitat and relationships, primary productivity, and nutrient cycling.
Author Contributions
S.P.G., W.R.H., L.K., I.M.B. and J.A. were all equally involved in the design, execution, and data analysis within the study. S.P.G. wrote the first draft of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
SG is funded by an Ulster University Vice Chancellors Doctoral Research Fellowship and received additional support through the G and M Williams Fund. Analytical costs were partly supported by core funding from the AFBI Aquatic Chemistry Laboratory (BH).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available at https://doi.org/10.5281/zenodo.7316452 accessed on 11 November 2022.
Acknowledgments
The authors acknowledge the support of Simon Exley and Emma Gorman for their assistance with the mesocosms (QML) and Mark Whyte (AFBI) for his analytical expertise with the Thermo Flash 2000 elemental analyzer. We also acknowledge Jeneil Biosurfactant, Saukville, WI, USA for providing rhamnolipid samples.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Battin, T.J.; Besemer, K.; Bengtsson, M.M.; Romani, A.M.; Packmann, A.I. The ecology and biogeochemistry of stream biofilms. Nat. Rev. Microbiol. 2016, 14, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Ren, Z.; Zhang, H.; Zhang, M.; Zhang, Y.; Liu, X.; Peng, W. Influences of anthropogenic land use on microbial community structure and functional potentials of stream benthic biofilms. Sci. Rep. 2017, 7, 15117. [Google Scholar] [CrossRef] [PubMed]
- Romani, A.M.; Guasch, H.; Munoz, I.; Ruana, J.; Vilalta, E.; Schwartz, T.; Emtiazi, F.; Sabater, S. Biofilm structure and function and possible implications for riverine DOC dynamics. Mirobiol. Ecol. 2004, 47, 316–328. [Google Scholar] [CrossRef]
- Kroukamp, O.; Wolfaardt, G.M. CO₂ Production as an indicator of biofilm metabolism. Appl. Environ. Microbiol. 2008, 75, 4391–4397. [Google Scholar] [CrossRef]
- Salta, M.; Wharton, J.A.; Blache, Y.; Stokes, K.R.; Briand, J. Marine biofilms on artificial surfaces: Structure and dynamics. Environ. Microbiol. 2013, 15, 2879–2893. [Google Scholar] [CrossRef]
- Hockin, N.L.; Mock, T.; Mulholland, F.; Kopriva, S.; Malin, G. The response of diatom central carbon metabolism to nitrogen starvation is different from that of green algae and higher plants. Plant Physiol. 2012, 158, 299–312. [Google Scholar] [CrossRef]
- Moore, L.J.; Fuentes, L.; Rodgers, J.H., Jr.; Bowerman, W.W.; Yarrow, G.K.; Chao, W.Y.; Bridges, W.C., Jr. Relative toxicity of the components of the original formulation of Roundup® to five North American anurans. Ecotoxicol. Environ. Saf. 2012, 78, 128–133. [Google Scholar] [CrossRef]
- Koedooder, C.; Stock, W.; Willems, A.; Mangelinckx, S.; De Troch, M.; Vyverman, W.; Sabbe, K. Diatom-bacteria interactions modulate the composition and productivity of benthic diatom biofilms. Front. Microbiol. 2019, 10, 1255. [Google Scholar] [CrossRef]
- Jacobs, P.; Pitarch, J.; Kromkamp, J.C.; Philippart, C.J. Assessing biomass and primary production of microphytobenthos in depositional coastal systems using spectral information. PLoS ONE 2021, 16, e0246012. [Google Scholar] [CrossRef]
- Rode, D.K.H.; Singh, P.K.; Drescher, K. Multicellular and unicellular responses of microbial biofilms to stress. Biol. Chem. 2020, 401, 1365–1374. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Neu, T.R.; Wozniak, D.J. The EPS matrix: The “house of biofilm cells”. J. Bacteriol. 2007, 189, 7945–7947. [Google Scholar] [CrossRef]
- De Beer, D.; Srinivasan, R.; Stewart, P.S. Direct measurement of chlorine penetration into biofilms during disinfection. Appl. Environ. Microbiol. 1994, 60, 4339–4344. [Google Scholar] [CrossRef]
- Mah, T.F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Pinto, R.M.; Soares, F.A.; Reis, S.; Nunes, C.; Van Dijck, P. Innovative strategies toward the disassembly of the EPS matrix in bacterial biofilms. Front. Microbiol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Di Martino, P. Extracellular polymeric substances, a key element in understanding biofilm phenotype. AIMS Microbiol. 2018, 4, 274. [Google Scholar] [CrossRef]
- DePas William, H.; Syed Adnan, K.; Margarita, S.; Lee John, S.; David, W.; Vinay, S.; Györgyi, C.; Boles Blaise, R.; Chapman Matthew, R.; Nojiri, H. Biofilm formation protects Escherichia coli against killing by Caenorhabditis elegans and Myxococcus xanthus. Appl. Environ. Microbiol. 2014, 80, 7079–7087. [Google Scholar] [CrossRef]
- Wucher, B.R.; Elsayed, M.; Adelman, J.S.; Kadouri, D.E.; Nadell, C.D. Bacterial predation transforms the landscape and community assembly of biofilms. Curr. Biol. 2021, 31, 2643–2651.e3. [Google Scholar] [CrossRef]
- Chan, S.Y.; Liu, S.Y.; Seng, Z.; Chua, S.L. Biofilm matrix disrupts nematode motility and predatory behavior. ISME J. 2021, 15, 260–269. [Google Scholar] [CrossRef]
- Collins, S.M.; Kohler, T.J.; Thomas, S.A.; Fetzer, W.W.; Flecker, A.S. The importance of terrestrial subsidies in stream food webs varies along a stream size gradient. Oikos 2016, 125, 674–685. [Google Scholar] [CrossRef]
- Margulis, L. Words as battle cries: Symbiogenesis and the new field of endocytobiology. Bioscience 1990, 40, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Skov, M.W.; Volkelt-Igoe, M.; Hawkins, S.J.; Jesus, B.; Thompson, R.C.; Doncaster, C.P. Past and present grazing boosts the photo-autotrophic biomass of biofilms. Mar. Ecol. Prog. Ser. 2010, 401, 101–111. [Google Scholar] [CrossRef]
- Collins, P.C.; Hunter, W.R.; Carlsson, J.; Carlsson, J. Fortuitous insights into the ecology of a recently charted deep-sea hydrothermal vent, using snails’ feet. Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 163, 103358. [Google Scholar] [CrossRef]
- Rugenski, A.T.; Murria, C.; Whiles, M.R. Tadpoles enhance microbial activity and leaf decomposition in a neotropical headwater stream. Freshw. Biol. 2012, 57, 1904–1913. [Google Scholar] [CrossRef]
- Atkinson, C.L.; Halvorson, H.M.; Kuehn, K.A.; Winebarger, M.; Hamid, A.; Waters, M.N. Filter-feeders have differential bottom-up impacts on green and brown food webs. Oecologia 2021, 195, 187–198. [Google Scholar] [CrossRef]
- Pamp, S.J.; Tolker-Nielsen, T. Multiple roles of biosurfactants in structural biofilm development by Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2531–2539. [Google Scholar] [CrossRef]
- Chrzanowski, Ł.; Ławniczak, Ł.; Czaczyk, K. Why do microorganisms produce rhamnolipids? World J. Microbiol. Biotechnol. 2012, 28, 401–419. [Google Scholar] [CrossRef]
- Schmitz, K.S. Chapter 4: Life Science. In Physical Chemistry; Elsevier: Boston, MA, USA, 2018; pp. 755–832. [Google Scholar]
- Milovanovic, M.; Arsenijevic, A.; Milovanovic, J.; Kanjevac, T.; Arsenijevic, N. Chapter 14: Nanoparticles in Antiviral Therapy. In Antimicrobial Nanoarchitectonics; Elsevier Inc.: Amsterdam, The Netherlands, 2017; pp. 383–410. ISBN 0323527337. [Google Scholar]
- Thakur, P.; Saini, N.K.; Thakur, V.K.; Gupta, V.K.; Saini, R.V.; Saini, A.K. Rhamnolipid the glycolipid biosurfactant: Emerging trends and promising strategies in the field of biotechnology and biomedicine. Microb. Cell Factories 2021, 20, 1–15. [Google Scholar] [CrossRef]
- Tornero, V.; Hanke, G. Chemical contaminants entering the marine environment from sea-based sources: A review with a focus on European seas. Mar. Pollut. Bull. 2016, 112, 17–38. [Google Scholar] [CrossRef]
- Grime, J.P. The CSR model of primary plant strategies—Origins, implications and tests. In Plant Evolutionary Biology; Springer: Berlin/Heidelberg, Germany, 1988; pp. 371–393. [Google Scholar]
- Malik, A.A.; Martiny, J.B.; Brodie, E.L.; Martiny, A.C.; Treseder, K.K.; Allison, S.D. Defining trait-based microbial strategies with consequences for soil carbon cycling under climate change. ISME J. 2020, 14, 1–9. [Google Scholar] [CrossRef]
- Van de Waal, D.B.; Elser, J.J.; Martiny, A.C.; Sterner, R.W.; Cotner, J.B. Editorial: Progress in ecological stoichiometry. Front. Microbiol. 2018, 9, 1957. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 1 July 2022).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: New York, NY, USA, 2018. [Google Scholar]
- Wickham, H. Package ‘ggplot2′: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Volume 10, pp. 978–980. [Google Scholar]
- Ogle, D.H.; Doll, J.C.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis; R Package Version 0.9; 2022. [Google Scholar]
- Simões, M.; Simões, L.C.; Pereira, M.O.; Vieira, M.J. Sodium dodecyl sulfate allows the persistence and recovery of biofilms of Pseudomonas fluorescens formed under different hydrodynamic conditions. Biofouling 2008, 24, 35–44. [Google Scholar] [CrossRef][Green Version]
- Lin, J.; Wang, Z.; Zang, Y.; Zhang, D.; Xin, Q. Detection of respiration changes inside biofilms with microelectrodes during exposure to antibiotics. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 2019, 54, 202–207. [Google Scholar] [CrossRef]
- Quinn, G.A.; Maloy, A.P.; Banat, M.M.; Banat, I.M. A comparison of effects of broad-spectrum antibiotics and biosurfactants on established bacterial biofilms. Curr. Microbiol. 2013, 67, 614–623. [Google Scholar] [CrossRef]
- Seiler, C.; Van Velzen, E.; Neu, T.R.; Gaedke, U.; Berendonk, T.U.; Weitere, M. Grazing resistance of bacterial biofilms: A matter of predators’ feeding trait. FEMS Microbiol. Ecol. 2017, 93, fix112. [Google Scholar] [CrossRef]
- Magalhães, C.M.; Bordalo, A.A.; Wiebe, W.J. Intertidal biofilms on rocky substratum can play a major role in estuarine carbon and nutrient dynamics. Mar. Ecol. Prog. Ser. 2003, 258, 275–281. [Google Scholar] [CrossRef]
- Colls Lozano, M.; Timoner, X.; Font, C.; Sabater, S.; Acuña, V. Effects of duration, frequency, and severity of the non-flow period on stream biofilm metabolism. Ecosystems 2019, 22, 1393–1405. [Google Scholar] [CrossRef]
- Gill, S.P.; Hunter, W.R.; Coulson, L.E.; Banat, I.M.; Schelker, J. Synthetic and biological surfactant effects on freshwater biofilm community composition and metabolic activity. Appl. Microbiol. Biotechnol. 2022, 106, 6847–6859. [Google Scholar] [CrossRef]
- Thompson, R.C.; Norton, T.A.; Hawkins, S.J. Physical stress and biological control regulate the producer–consumer balance in intertidal biofilms. Ecology 2004, 85, 1372–1382. [Google Scholar] [CrossRef]
- Androuin, T.; Polerecky, L.; Decottignies, P.; Dubois, S.F.; Dupuy, C.; Hubas, C.; Jesus, B.; Le Gall, E.; Marzloff, M.P.; Carlier, A. Subtidal microphytobenthos: A secret garden stimulated by the engineer species Crepidula fornicata. Front. Mar. Sci. 2018, 5, 475. [Google Scholar] [CrossRef]
- Hillebrand, H.; Kahlert, M. Effect of grazing and nutrient supply on periphyton biomass and nutrient stoichiometry in habitats of different productivity. Limnol Oceanograph 2001, 46, 1881–1898. [Google Scholar] [CrossRef]
- Heisterkamp, I.M.; Schramm, A.; Larsen, L.H.; Svenningsen, N.B.; Lavik, G.; de Beer, D.; Stief, P. Shell biofilm-associated nitrous oxide production in marine molluscs: Processes, precursors and relative importance. Environ. Microbiol. 2013, 15, 1943–1955. [Google Scholar] [CrossRef] [PubMed]
- Nickzad, A.; Déziel, É. The involvement of rhamnolipids in microbial cell adhesion and biofilm development—An approach for control. Lett. Appl. Microbiol. 2014, 58, 447–453. [Google Scholar] [CrossRef]
- Du, M.; Xu, D.; Trinh, X.; Liu, S.; Wang, M.; Zhang, Y.; Wu, J.; Zhou, Q.; Wu, Z. EPS solubilization treatment by applying the biosurfactant rhamnolipid to reduce clogging in constructed wetlands. Bioresour. Technol. 2016, 218, 833–841. [Google Scholar] [CrossRef]
- Russell, B.D.; Connell, S.D.; Findlay, H.S.; Tait, K.; Widdicombe, S.; Mieszkowska, N. Ocean acidification and rising temperatures may increase biofilm primary productivity but decrease grazer consumption. Philos. Trans. R. Soc. 2013, 368, 20120438. [Google Scholar] [CrossRef]
- Calapez, A.R.; Elias, C.L.; Alves, A.; Almeida, S.F.P.; Brito, A.G.; Feio, M.J. Shifts in biofilms’ composition induced by flow stagnation, sewage contamination and grazing. Ecol. Indic. 2020, 111, 106006. [Google Scholar] [CrossRef]
- Muñoz, I.; Real, M.; Guasch, H.; Navarro, E.; Sabater, S. Effects of atrazine on periphyton under grazing pressure. Aquat. Toxicol. 2001, 55, 239–249. [Google Scholar] [CrossRef]
- Rosi, E.J.; Bechtold, H.A.; Snow, D.; Rojas, M.; Reisinger, A.J.; Kelly, J.J. Urban stream microbial communities show resistance to pharmaceutical exposure. Ecosphere 2018, 9, e02041. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).