Abstract
Bile acids (BAs) have been widely used on livestock in recent years and could be a good feed additive to improve the performances of animals. But the effects of bile acid on cats have not been studied. The purpose of this research was to investigate the effects of the bile acid from ducks on cats. Five healthy cats, which were two years old, were studied with a dose of the bile acid product being 50 mg/kg body weight per day. On day 0 (group A) and day 28 (group B), the fecal flora, volatile fatty acids, fat digestibility, and blood biochemical Indices of cats were studied and tested. The results showed that BAs could probably modulate the fat metabolism and gut microflora of cats and could improve the digestibility of different nutritional ingredients. The cholesterol of blood was greatly decreased and the cholesterol and activity of amylase were improved by 20.9% (p < 0.05) and 21.1% (p < 0.05), respectively. Fat digestibility was improved by 5.68% (p < 0.05) and the body weight on day 28 was decreased without significance (p > 0.05). The abundance of the Bifidobacterium on day 28 was greatly increased (p < 0.05) after using BAs product. The research showed that duck BAs could modulate fat metabolism and increase probiotics of the gut in cats and could probably be a good feed additive to reduce weight and improve gut health for cats in the future.
1. Introduction
Bile acids (BAs) have been widely used on livestock in recent years and could be a good feeding supplement for animals. Previous studies showed that bile acids (BAs) are a kind of important metabolites in the fat metabolism in human beings and animals [1]. Many kinds of BAs were detected in the gallbladder of animals, including tauroursodeoxycholic acid, taurocholic acid, taurochenodeoxycholic acid, taurodeoxycholic acid, taurolithocholic acid, cholic acid, and glycochenodeoxycholic acid [2]; in which taurocholic acid, taurochenodeoxycholic acid and taurodeoxycholic acid were dominant. BAs can emulsify foods, facilitate the digestion of fats and be important as signals in the fat metabolism process [2]. Previous research had shown that BAs were related with many diseases of human beings, such as cancers [3], inflammation, and some chronic liver diseases [4].
BAs of different sources could not only improve the performances and nutrient digestibility of animals, reduce intestinal permeability, and reduce the heat stress [5,6,7] but BAs have been proved to also have a close relationship with the gut microbiome, which was reported before [8,9,10]. Also, it has been shown that BAs could be an important signal in many metabolic pathways and modulate many diseases, like irritable bowel syndrome [9,11], liver diseases [10,12,13], some chronic illnesses [14], obesity [15], etc. However, there are just a few studies of duck bile acid extract on pets, especially cats. BAs are vital to the health of cats. They act as digestive juices that facilitate the breakdown and absorption of lipids. Additionally, they aid in the excretion of waste substances such as cholesterol and bilirubin. In doing so, the liver is able to perform its essential functions of maintaining nutrition and safeguarding the body. Also, as duck bile acid extract is easy to acquire and the cost is appropriate for its application, we will choose duck BAs for further studies.
Therefore, this study aims to evaluate the role of the BAs extract, isolated from the duck, in healthy cats for the purpose of detecting fecal microbiota, short chain fatty acids (SCFAs), fat digestibility and blood biochemical Indices.
2. Materials and Methods
2.1. Experimental Animals
Five cats of about two years old (each about 3 kg in weight, female, healthy, British shorthair) were used in the test, were self-breeding by our test base, and they were numbered 8, 9, 13, 17, 24, and marked by ourselves. The cat housing in the experiment consisted of five stainless steel cages (162 × 68 × 189 cm) and the temperature of the room was 25 ± 2 °C. Each cage was furnished with litter boxes and food bowls. Fresh water was available all the time. Cats were fed once every day at the same time.
A within-subject study design was used in this test with each cat representing its own control to exclude the factors of between-subject variation. The same method was conducted by Butterwick et al. [16]. The test was implemented for 28 days. Before the test, the body weight and the fat digestibility of each cat was tested, and the daily food requirement of cats was calculated according to the weight. The feces and the blood were also collected before the test. The concentration of exogenous duck bile acid extract (Hunan Yungang Biotechnology Co., Ltd., Changde, China) was about 50 mg/kg body weight, which was added to the food of each cat every day. During the test each cat was guaranteed to eat all the food completely. The collection of stool and blood samples was performed for the detection of subsequent Indices. On day 28, the feces and blood samples were collected. The feces on day 0 and day 28 were collected from each section of fresh feces and stored at −80 °C. At the same time, the blood samples on day 0 and day 28 were collected for the biochemical test. All the treated cats did not use any antibiotics and probiotics during the test (Figure 1).

Figure 1.
Experimental design and scheme of the animal treatments n = 5.
2.2. Blood Biochemical Test
In this test, the collected non-anticoagulant blood was harvested by centrifugation at 1200× g for 15 min at 4 °C and the serum was separated using the blood biochemical test. A biochemical analyzer (MNCHIP, Tianjin, China) was used to test biochemical parameters of the supernatant of the blood, including the total protein (TP), albumin (ALB), total bilirubin (TBIL), total bile acid (TBA), triglyceride (TG), cholesterol (CHOL), globulin (GLO), and amylase activity (AMY).
2.3. Digestibility of Fat of the Feed
For the determination of fat digestibility, the feces of each cat were collected for 3 days (day 25–27) continuously. In order to prevent interference with the test, cat hair was taken out of the excrement. The food and feces samples were separately analyzed for crude fat using the routine procedures (AOAC, 1984).
The formula of the digestibility of fat was as follows:
The Digestibiliy of Fat = ((fat intake of feed − fat in the feces)/fat intake of feed) × 100%
2.4. The Test of SCFAs of the Feces
The SCFAs of the feces samples on day 0 and day 28 were tested using ion chromatography (Metrohm, Herisau, Switzerland). A total of four samples of SCFAs were detected. An amount of 0.5 g of the feces sample was collected and dissolved in 200 µL of ZnSO4 (144 g/L) and 100 µL of K4Fe(CN)6 3H2O, and then the solution was fixed to 5 mL. The solution was centrifuged at 5180× g for 10 min and 1 mL of the supernatant with 4 mL water was mixed and tested for SCFAs.
2.5. Extraction of Fecal DNA
Total DNA was extracted from fecal samples without culturing. E.Z.N.A Mag-Bind Soil DNA Kit (Omega, M5635-02, San Antonio, TX, USA) and Quibit dsDNA HS (Thermo, Waltham, MA, USA) was used to test the concentration of DNA samples. The extracted DNA samples were stored in a refrigerator at −70 °C and used for the next PCR amplification.
2.6. PCR Amplification
The sequence of the forward primer was CCTACGGGNGGCWGCAG and the sequence of the reverse primer was GACTACHVGGGTATCTAATCC. PCR was amplified twice. The first PCR reaction conditions were as follows: 94 °C, 3 min, 5 cycles at 94 °C, 30 s, 45 °C 20 s, 65 °C 30 s; 20 cycles at 94 °C 20 s, 55 °C 20 s, 72 °C, 30 s, 72 °C, 5 min. The conditions for the second time were as follows: 95 °C, 3 min, 5 cycles at 94 °C, 20 s, 55 °C, 22 s, 72 °C, 30 s; 72 °C 5 min.
PCR products were purified and quantified by the QIA quick gel extraction kit (QIAGEN, Hilden, Germany) and Quant-iT PicoGreen dsDNA assay kit (Life Technologies, Carlsbad, CA, USA), respectively. The extracted DNA samples were sequenced for 16S rDNA in Sangon Biotech Co., Ltd., Shanghai, China. The α and β diversities of the bacterial community were calculated on the free online analysis platform (www.sangon.com) based on Chao 1 and the unweighted UniFrac distance. The main differential abundant genera were selected by the linear discriminant analysis effect size (LEfSe) method. Furthermore, the predicted microbial metabolic functions were conducted by PICRUST2 (https://huttenhower.sph.harvard.edu/galaxy (accessed on 25 March 2022)). The heatmap package of R (Version 3.3.1) and Python (Version 2.7) was applied to generate heatmaps.
2.7. Data Analysis
The alpha diversity indices were identified in terms of OTU richness. All alpha diversity indices were calculated with Mothur software (Version 3.8.31). Beta diversity was used to evaluate the differences in the microbiome among samples and was normally combined with dimensional reduction methods, such as principal coordinate analysis (PCoA), to obtain visual representations. A differential comparison was used to identify features with significantly different abundances between groups using STAMP (Version 2.1.3) and LefSe (Version 1.1.0). Correlation coefficients and p-values between communities/OTUs were calculated using SparCC (Version 1.1.0).
Data of blood, SCFAs and performance (fat digestibility and body weight) were analyzed with one-way ANOVA followed by Tukey’s multiple range test. Data were expressed as the mean ± SE by Tukey’s multiple range test and significance was expressed when the p was less than 0.05. These related statistical analyses were performed by SPSS 25.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Fat Digestibility and Body Weight
From Table 1, it could be concluded that body weight tended to have a decreasing trend (p > 0.05) and that the fat digestibility greatly improved by 5.68% (p < 0.05).

Table 1.
Effect on growth, fat digestibility and SCFAs in feline feces.
3.2. SCFAs Concentration
The result of four SCFAs of the feces samples on day 0 and day 28 are shown in Table 1. The concentration of butyrate acid on day 28 was greatly enhanced by 110.6% compared to day 0 (p < 0.05) while the acetic acid and propionic acid were also increased without significance (p > 0.05).
3.3. Blood Biochemical Indices
The blood biochemical Indices included total protein (TP), albumin (ALB), total bilirubin (TBIL), globulin (GLO), total bile acid (TBA), amylase activity (AMY) triglyceride (TG), and cholesterol (CHOL). These Indices were tested using the biochemical analyzer (MNCHIP Technologies Co., Ltd., Tianjin, China). The cholesterol on Day 28 (Group B) was decreased by 20.9% compared to Day 0 (Group A) (p < 0.05), and the total bilirubin was improved by 48.2% within a healthy range (p < 0.05). The activity of amylase was also improved by 21.2% within a healthy range (p < 0.05) (Table 2).
Table 2.
Alterations in blood biochemical indices in cats.
3.4. Fecal Microbiota Analysis
The fecal samples of the cats were sequenced. Rank abundance curves can reflect the abundance of microbiota community. The data revealed the rarefaction curve, as shown in Figure 2a, which meant the sample size was sufficient to prove the richness of the microbiota community. The alpha diversity of the microbial community was measured using Chao1 Indices, Shannon Indices, ACE Indices and Simpson Indices, and there were no significant differences between group A and group B (p > 0.05) (Figure 2c). Beta diversity evaluates differences in the microbiome among samples and was normally combined with dimensional reduction methods, such as principal coordinate analysis (PCoA), to obtain visual representations (Figure 2d).
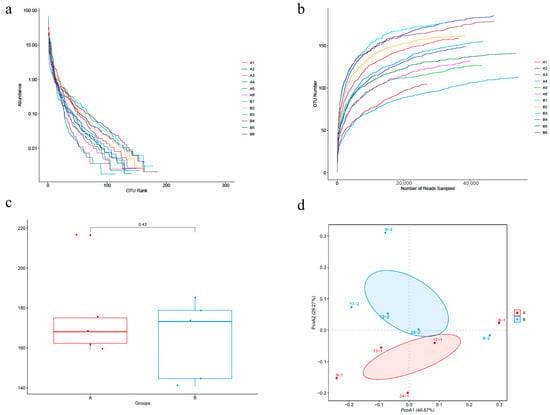
Figure 2.
BAs alters gut microbiota richness and diversity in cats. (a) Rank abundance curves. (b) α-diversity indices dilution curves. (c) Chao 1 indices were used to analyze α-diversity of feline fecal microbiota in group A and B. (d) β-diversity of fecal microflora was determined by principal coordinate analysis (PCoA).
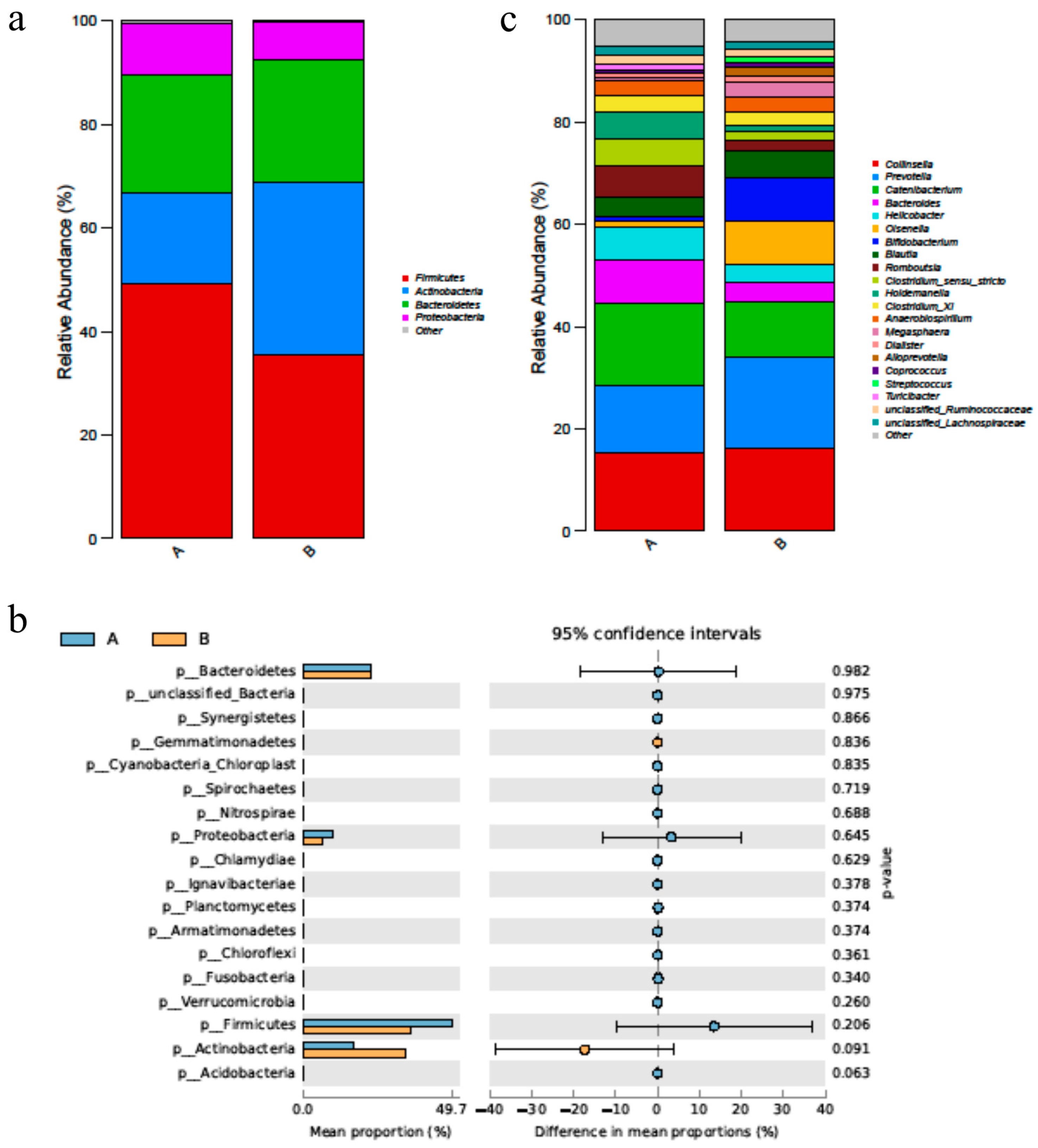
After supplementation of duck bile acid extract, the abundance of Actinobacteria was increased greatly, and the abundance of Firmicutes and Proteobacteria were decreased (p > 0.05) (Figure 3a,b). At the genus level, the abundance of Bifidobacterium, Lactobacillus and Megasphaera, the core probiotics in the gut, was improved after using the bile acid product (Figure 3c).
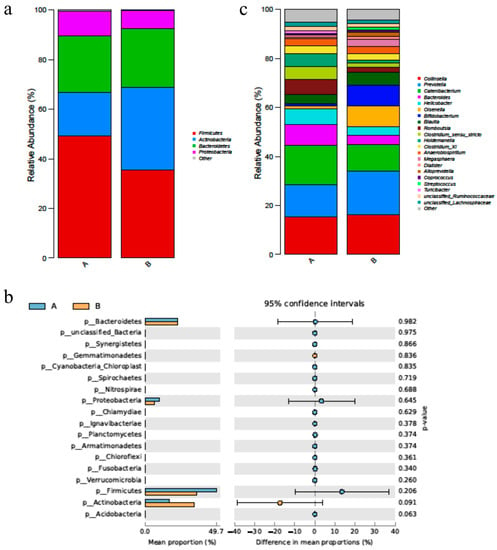
Figure 3.
BAs alters the diversified species of fecal microbiota in cat. (a) The figure shows the foundation of the phylum level of changes in the fecal microbiota, especially dominant phylum, such as Firmicutes and Bacteroides. (b) The figure showed the p value of the phylum level of fecal microbiota of group A and B. (c) The figure showed the genus level of the changes in the fecal microbiota. n = 5. Letters represented the taxonomy of the bacteria: p, phylum.
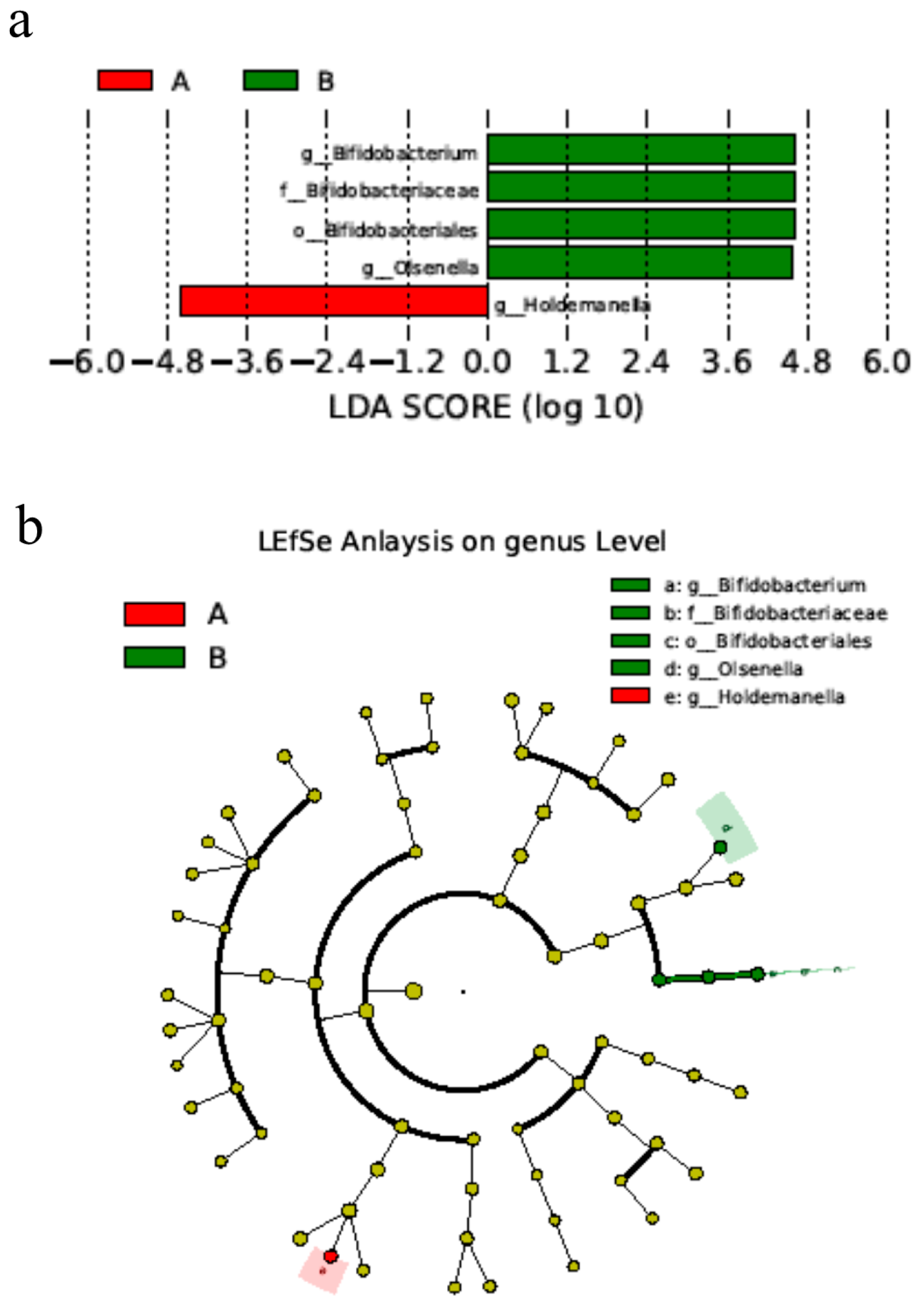
Differences in microbiota were also determined using LEfSe. Significance in species was observed between day 0 (Group A) and day 28 (Group B). The abundance of genus Bifidobacterium spp. on day 28 was higher after using Bas, and the genus Holdemanella was more abundant on day 0 than day 28 (Figure 4) (p < 0.05).
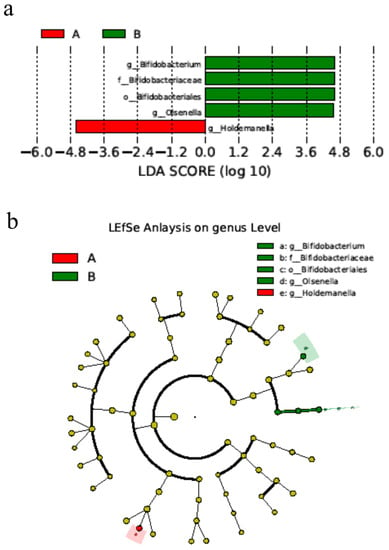
Figure 4.
BAs alters the microbial differences in bacterial communities in cats. (a) Bacterial classification between day 0 and day 28 of cats. (b) Cladogram from the linear discriminant analysis of the effect size on day 0 and day 28. n = 5. Notes: Small letters represented the taxonomy of the bacteria: o, order; f, family; g, genus.
4. Discussion
After BAs enters the digestive tract of cats orally, it is transformed into secondary bile acids (SBAs) by interacting with various flora colonized in the small intestine, and finally it is passively diffused and reabsorbed by intestinal epithelial cells into the intestinal-hepatic circulation or excreted into feces [9,17].
The research showed that on the phylum level, exogenous BAs could increase the abundance of the Actinobacteria, and decrease the abundance of Firmicutes, which may be different from the results observed before [18]. The reason for this was probably because the metabolic pathway of BAs in felines was different from humans and rats. For cats, BAs were exclusively conjugated with taurine, while the taurine in humans and rats was separated. BAs could also be conjugated with glycine; however, the main bile acids in cats also differ from humans [4]. On the genus level, the abundance of Bifidobacterium spp., the beneficial bacteria in the gut [19] was greatly increased which could indicate that BAs could improve gut health by modulating the gut microbiota. The mechanism was probably that Bifidobacterium spp. could produce the bile salt hydrolase and was bile-metabolizing. This result was also proved by former studies [20]. The research also showed that BAs could inhibit the Helicobacter pylori of the cats, which coincided with the former studies [21,22].
The protein and fat content were high in the food of cats and because of the lack of exercise the obesity problem often happens in cats; thus, this obesity problem may present clinical hyperlipidemia and hypercholesteremia problems. In this research, the cholesterol was significantly decreased after using BAs, which coincided with former studies [23], and showed that BAs could be a good supplement to modulate fat metabolism. The mechanism for the reduction of cholesterol could be related with the result of the increase of the abundance of Bifidobacterium spp. as the former study showed that Bifidobacterium spp. lowers cholesterol activity [24,25]. The mechanism for lowering cholesterol by Bifidobacterium spp. was not very clear until now [24]. The research also found that, after adding BAs, fat digestibility was improved greatly (p < 0.05), which could reduce strain on the liver for cats. The same results were proved on poultry [26].
Amylase was very important for cats, and it could help felines to digest the starch in the food to provide energy for the body, but their saliva could not produce amylase like in humans; thus, the production of amylase could probably be reflected by the function of the pancreas in cats [27]. In this research, the amylase activity was increased greatly after using BAs, which meant that BAs could improve the pancreas function indirectly. Similar results were also found in aquatilia hydrocoles [28].
SCFAs were very important in modulating life activities and material metabolism which could supply energy for intestinal epithelial cells and be beneficial to the health of humans and animals [29]. The changes of the SCFAs were related with the gut’s microflora [30]. Four SCFAs, including acetic acid, propionic acid, butyric acid, and isovaleric acid were studied in this research. And among these SCFAs, the butyric acid was improved after using BAs (p < 0.05) and acetic acid and propionic acid had the trend of increasing (p > 0.05), which was probably related to the increase of the species producing SCFAs, such as Bifidobacteria. Similar results were also showed in vivo and in vitro in former studies [30,31], and the mechanism worked as the Bifidobacteria could modulate the receptor GPR43 of SCFAs, which was already proved before [30].
The research focused solely on the result, and more research about the mechanism will be needed in future. Also, this research proved the positive effect of the BAs; however, the treatments were limited. In the future, various concentrations need to be studied to make sure the best concentration of BAs. Also, taking measurements in the middle of the test (e.g., day 14) should be considered in future tests.
5. Conclusions
Duck BAs can promote cat fat metabolism and cause changes in cat intestinal microflora and affect cat blood biochemical Indices and fecal odor content. All these Indices indicate the potential use of duck bile acid extract in cat clinical practice. These could probably supply a good way to reduce weight and improve gut health for cats in future.
Author Contributions
Conceptualization, B.H.; methodology, B.H. and P.Z.; software, B.H. and Z.W.; validation, B.H., P.Z., Z.W., H.T., X.W., J.L. and J.W.; formal analysis, B.H.; investigation, B.H.; resources, B.H.; data curation, B.H. and S.L.; writing—original draft preparation, B.H.; writing—review and editing, B.H.; visualization, B.H.; supervision, B.H. and J.W.; project administration, B.H. and P.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The Agricultural Science and Technology Innovation Program (ASTIP).
Institutional Review Board Statement
The animal experiment was performed according to the Animal Care and Use Committee of the Institute of Feed Research of Chinese Academy of Agricultural Sciences (CAAS) and approved by the Laboratory Animal Ethical Committee and its inspection of the Institute of Feed Research of CAAS (IFR-CAAS-20220101).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Washizu, T.; Ikenaga, H.; Washizu, M.; Ishida, T.; Tomoda, I.; Kaneko, J.J. Bile acid composition of dog and cat gall-bladder bile. Jpn. J. Vet. Sci. 1990, 52, 423–425. [Google Scholar] [CrossRef] [PubMed]
- Duszka, K. Versatile Triad Alliance: Bile Acid, Taurine and Microbiota. Cells 2022, 11, 2337. [Google Scholar] [CrossRef]
- Ocvirk, S.; O’Keefe, S.J. Influence of Bile Acids on Colorectal Cancer Risk: Potential Mechanisms Mediated by Diet-Gut Microbiota Interactions. Curr. Nutr. Rep. 2017, 6, 315–322. [Google Scholar] [CrossRef]
- Liu, Y.; Kang, W.; Liu, S.; Liu, J.; Chen, X.; Gan, F.; Huang, K. Gut microbiota–bile acid–intestinal Farnesoid X receptor signaling axis orchestrates cadmium-induced liver injury. Sci. Total. Environ. 2022, 849, 157861. [Google Scholar] [CrossRef]
- Alzawqari, M.H.; Moghaddam, H.N.; Kermanshahi, H.; Raji, A.R. The effect of desiccated ox bile supplementation on performance, fat digestibility, gut morphology and blood chemistry of broiler chickens fed tallow diets. J. Appl. Anim. Res. 2011, 39, 169–174. [Google Scholar] [CrossRef]
- de Diego-Cabero, N.; Mereu, A.; Menoyo, D.; Holst, J.J.; Ipharraguerre, I.R. Bile acid mediated effects on gut integrity and performance of early-weaned piglets. BMC Veter. Res. 2015, 11, 111. [Google Scholar] [CrossRef]
- Lai, W.Q.; Huang, W.G.; Dong, B.; Cao, A.; Zhang, W.; Li, J.; Wu, H.; Zhang, L. Effects of dietary supplemental bile acids on performance, carcass characteristics, serum lipid metabolites and intestinal enzyme activities of broiler chickens. Poult. Sci. 2018, 97, 196–202. [Google Scholar] [CrossRef]
- Mori, H.; Svegliati Baroni, G.; Marzioni, M.; Di Nicola, F.; Santori, P.; Maroni, L.; Scarpellini, E. Farnesoid X Receptor, Bile Acid Metabolism, and Gut Microbiota. Metabolites 2022, 12, 647. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.W.; Rezaie, A.; Pimentel, M. Bile Acid and Gut Microbiota in Irritable Bowel Syndrome. J. Neurogastroenterol. Motil. 2022, 28, 549–561. [Google Scholar] [CrossRef]
- Wu, L.; Feng, J.; Li, J.; Yu, Q.; Ji, J.; Wu, J.; Dai, W.; Guo, C. The gut microbiome-bile acid axis in hepatocarcinogenesis. Biomed. Pharmacother. 2020, 133, 111036. [Google Scholar] [CrossRef]
- Gu, Y.; Li, L.; Yang, M.; Liu, T.; Song, X.; Qin, X.; Xu, X.; Liu, J.; Wang, B.; Cao, H. Bile acid–gut microbiota crosstalk in irritable bowel syndrome. Crit. Rev. Microbiol. 2022, 49, 350–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-L.; Li, Z.-J.; Gou, H.-Z.; Song, X.-J.; Zhang, L. The gut microbiota–bile acid axis: A potential therapeutic target for liver fibrosis. Front. Cells Infect. Microbiol. 2022, 12, 945368. [Google Scholar] [CrossRef] [PubMed]
- Simbrunner, B.; Trauner, M.; Reiberger, T. Review article: Therapeutic aspects of bile acid signalling in the gut-liver axis. Aliment. Pharmacol. Ther. 2021, 54, 1243–1262. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zhuo, L.-B.; He, Y.; Fu, Y.; Shen, L.; Xu, F.; Gou, W.; Miao, Z.; Shuai, M.; Liang, Y.; et al. The gut microbiota-bile acid axis links the positive association between chronic insomnia and cardiometabolic diseases. Nat. Commun. 2022, 13, 3002. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Huang, F.; Zhao, L.; Zhang, Y.; Yang, W.; Wang, S.; Li, M.; Han, X.; Ge, K.; Qu, C.; et al. A dysregulated bile acid-gut microbiota axis contributes to obesity susceptibility. EBioMedicine 2020, 55, 102766. [Google Scholar] [CrossRef]
- Marshall-Jones, Z.V.; Baillon, M.-L.A.; Croft, J.M.; Butterwick, R.F. Effects of Lactobacillus acidophilus DSM13241 as a probiotic in healthy adult cats. Am. J. Veter. Res. 2006, 67, 1005–1012. [Google Scholar] [CrossRef]
- Anna, V.G.; Susan, A.J.; Gerard, M.; Aurelijus, B.; Eoin, S.; Cryan, J.F. Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. Ebiomedicine 2017, 24, 166–178. [Google Scholar]
- Islam, K.S.; Fukiya, S.; Hagio, M.; Fujii, N.; Ishizuka, S.; Ooka, T.; Ogura, Y.; Hayashi, T.; Yokota, A. Bile Acid Is a Host Factor That Regulates the Composition of the Cecal Microbiota in Rats. Gastroenterology 2011, 141, 1773–1781. [Google Scholar] [CrossRef]
- Hidalgo-Cantabrana, C.; Delgado, S.; Ruiz, L.; Ruas-Madiedo, P.; Sánchez, B.; Margolles, A. Bifidobacteria and Their Health-Promoting Effects. Microbiol. Spectr. 2017, 5, 73–98. [Google Scholar] [CrossRef]
- Klaver, F.A.; van der Meer, R. The assumed assimilation of cholesterol by Lactobacilli and Bifidobacterium bifidum is due to their bile salt-deconjugating activity. Appl. Environ. Microbiol. 1993, 59, 1120–1124. [Google Scholar] [CrossRef]
- Okoli, A.S.; Raftery, M.J.; Mendz, G.L. Effects of human and porcine bile on the proteome of Helicobacter hepaticus. Proteome Sci. 2012, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Itoh, M.; Wada, K.; Tan, S.; Kitano, Y.; Kai, J.; Makino, I. Antibacterial action of bile acids against Helicobacter pylori and changes in its ultrastructural morphology: Effect of unconjugated dihydroxy bile acid. J. Gastroenterol. 1999, 34, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Burrin, D.; Stoll, B.; Moore, D. Digestive physiology of the pig symposium: Intestinal bile acid sensing is linked to key endocrine and metabolic signaling pathways. J. Anim. Sci. 2013, 91, 1991–2000. [Google Scholar] [CrossRef]
- Fernandez-Calderon, M.C.; Sanchez-Moro, M.D.H.; Rincon, E.O. In vitro Cholesterol Assimilation by Bifidobacterium animalis subsp. lactis (BPL1) Probiotic Bacteria Under Intestinal Conditions. Endocr. Metab. Immune Disord. Drug Targets 2022, 22, 433–439. [Google Scholar] [CrossRef]
- Zanotti, I.; Turroni, F.; Piemontese, A.; Mancabelli, L.; Milani, C.; Viappiani, A.; Ventura, M. Evidence for cholesterol-lowering activity by Bifidobacterium bifidum PRL2010 through gut microbiota modulation. Appl. Microbiol. Biotechnol. 2015, 99, 6813–6829. [Google Scholar] [CrossRef]
- Geng, S.; Zhang, Y.; Cao, A.; Liu, Y.; Di, Y.; Li, J.; Lou, Q.; Zhang, L. Effects of Fat Type and Exogenous Bile Acids on Growth Performance, Nutrient Digestibility, Lipid Metabolism and Breast Muscle Fatty Acid Composition in Broiler Chickens. Animals 2022, 12, 1258. [Google Scholar] [CrossRef]
- Mao, Y.; Fang, L.; Ai, L.; Li, C.; Wang, Z.; Wu, J.; Li, F. An in vivo study of the effects on serum glucose, amylase and histopathology of the feline pancreatic tissue treated by focused ultrasound. PLoS ONE 2014, 9, e88815. [Google Scholar] [CrossRef]
- Li, Y.; Wang, S.; Hu, Y.; Cheng, J.; Cheng, X.; Cheng, P.; Cui, Z. Dietary bile acid supplementation reveals beneficial effects on intestinal healthy status of tongue sole (Cynoglossus semiliaevis). Fish Shellfish Immunol. 2021, 116, 52–60. [Google Scholar] [CrossRef]
- Sasaki, D.; Sasaki, K.; Abe, A.; Ozeki, M.; Kondo, A. Effects of partially hydrolyzed guar gums of different molecular weights on a human intestinal in vitro fermentation model. J. Biosci. Bioeng. 2023, 136, 67–73. [Google Scholar] [CrossRef]
- Horiuchi, H.; Kamikado, K.; Aoki, R.; Suganuma, N.; Nishijima, T.; Nakatani, A.; Kimura, I. Bifidobacterium animalis subsp. lactis GCL2505 modulates host energy metabolism via the short-chain fatty acid receptor GPR43. Sci. Rep. 2020, 10, 4158. [Google Scholar] [CrossRef]
- Zampa, A.; Silvi, S.; Fabiani, R.; Morozzi, G.; Orpianesi, C.; Cresci, A. Effects of different digestible carbohydrates on bile acid metabolism and SCFA production by human gut micro-flora grown in an in vitro semi-continuous culture. Anaerobe 2004, 10, 19–26. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).