
Anticancer Properties of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Potential Agonists 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370 in Colorectal Adenocarcinoma Cells In Vitro

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Synthesis of 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370
2.3. Cell Culture and Treatment
2.4. Resazurin Reduction Assay
2.5. LDH Cytotoxicity Assay
2.6. Measurement of Intracellular ROS Level
2.7. Caspase-3 Activity Assay
2.8. Rhodamine-B and Hoechst 33,342 Staining
2.9. qPCR Analysis of PPARγ, AhR, PXR, and NF-κB Genes
2.10. Western Blot
2.11. Statistical Analysis
2.12. Analysis of Molecular Docking Simulations
3. Results
3.1. Metabolic Activity
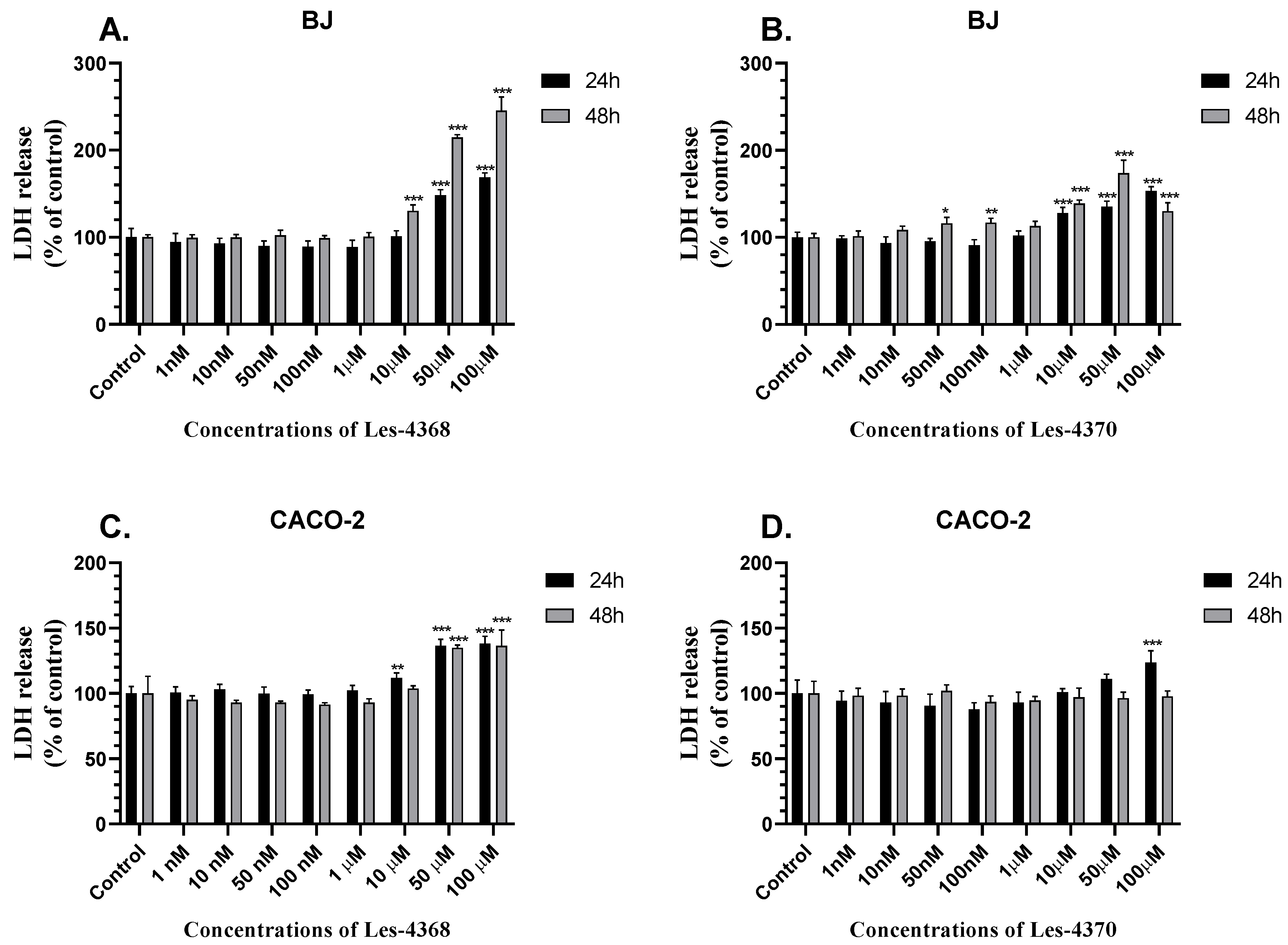
3.2. LDH Release Level
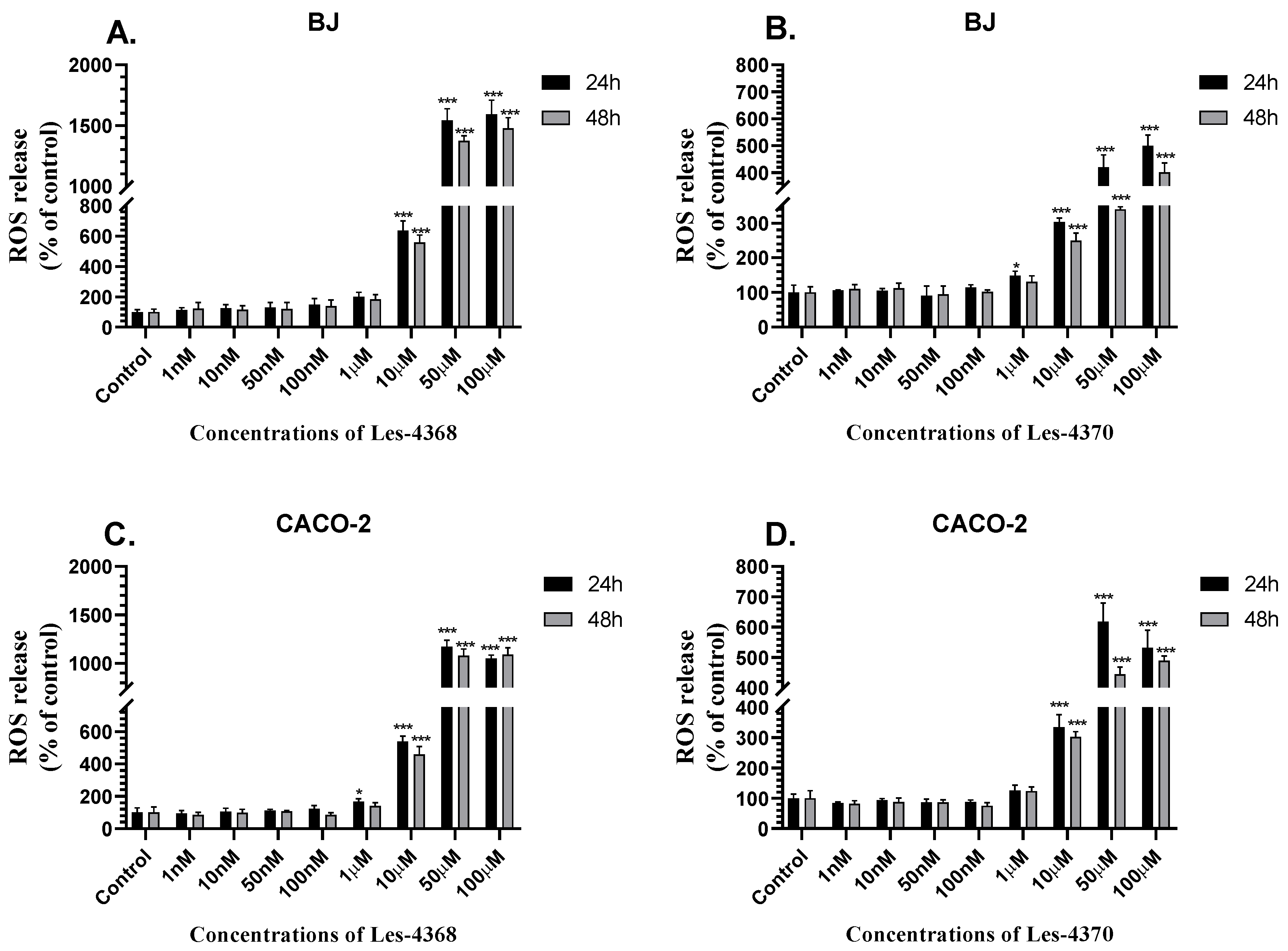
3.3. Intracellular ROS Level
3.4. Caspase-3 Activity
3.5. Cell Viability and Apoptotic Body Formation Staining
3.6. mRNA Expression of the PPARγ, AhR, PXR, and NF-κB Genes
3.7. AhR and PPARγ Protein Level
3.8. Molecular Docking Simulations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Fontana, F.; Anselmi, M.; Limonta, P. Molecular Mechanisms of Cancer Drug Resistance: Emerging Biomarkers and Promising Targets to Overcome Tumor Progression. Cancers 2022, 14, 1614. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, L.; Shi, H.; Chen, H.; Tao, J.; Shen, R.; Wang, T. Ursolic Acid Enhances the Therapeutic Effects of Oxaliplatin in Colorectal Cancer by Inhibition of Drug Resistance. Cancer Sci. 2018, 109, 94. [Google Scholar] [CrossRef]
- Debela, D.T.; Muzazu, S.G.; Heraro, K.D.; Ndalama, M.T.; Mesele, B.W.; Haile, D.C.; Kitui, S.K.; Manyazewal, T. New Approaches and Procedures for Cancer Treatment: Current Perspectives. SAGE Open Med. 2021, 9, 205031212110343. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Gomtsyan, A. Heterocycles in Drugs and Drug Discovery. Chem. Heterocycl. Compd. 2012, 48, 7–10. [Google Scholar] [CrossRef]
- Roszczenko, P.; Holota, S.; Szewczyk, O.K.; Dudchak, R.; Bielawski, K.; Bielawska, A.; Lesyk, R. 4-Thiazolidinone-Bearing Hybrid Molecules in Anticancer Drug Design. Int. J. Mol. Sci. 2022, 23, 13135. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Leja, M.L.; Kaminskyy, D.V.; Binduga, U.E.; Pinyazhko, O.R.; Lesyk, R.B.; Gmiński, J. Study of Novel Anticancer 4-Thiazolidinone Derivatives. Chem. Biol. Interact. 2017, 262, 46–56. [Google Scholar] [CrossRef]
- Bar, M.; Skóra, B.; Tabęcka-Łonczyńska, A.; Holota, S.; Khyluk, D.; Roman, O.; Lesyk, R.; Szychowski, K.A. New 4-Thiazolidinone-Based Molecules Les-2769 and Les-3266 as Possible PPARγ Modulators. Bioorg. Chem. 2022, 128, 106075. [Google Scholar] [CrossRef]
- Skóra, B.; Lewińska, A.; Kryshchyshyn-Dylevych, A.; Kaminskyy, D.; Lesyk, R.; Szychowski, K.A. Evaluation of Anticancer and Antibacterial Activity of Four 4-Thiazolidinone-Based Derivatives. Molecules 2022, 27, 894. [Google Scholar] [CrossRef] [PubMed]
- Szychowski, K.A.; Kaminskyy, D.V.; Leja, M.L.; Kryshchyshyn, A.P.; Lesyk, R.B.; Tobiasz, J.; Wnuk, M.; Pomianek, T.; Gmiński, J. Anticancer Properties of 5Z-(4-Fluorobenzylidene)-2-(4-Hydroxyphenylamino)-Thiazol-4-One. Sci. Rep. 2019, 9, 10609. [Google Scholar] [CrossRef]
- Jarrar, M.H.; Baranova, A. PPARγ Activation by Thiazolidinediones (TZDs) May Modulate Breast Carcinoma Outcome: The Importance of Interplay with TGFβ Signalling. J. Cell. Mol. Med. 2007, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- Kota, B.P.; Huang, T.H.W.; Roufogalis, B.D. An Overview on Biological Mechanisms of PPARs. Pharmacol. Res. 2005, 51, 85–94. [Google Scholar] [CrossRef]
- Augimeri, G.; Bonofiglio, D. PPARgamma: A Potential Intrinsic and Extrinsic Molecular Target for Breast Cancer Therapy. Biomedicines 2021, 9, 543. [Google Scholar] [CrossRef]
- Kryshchyshyn, A.; Atamanyuk, D.; Lesyk, R. Fused Thiopyrano[2,3-d]Thiazole Derivatives as Potential Anticancer Agents. Sci. Pharm. 2012, 80, 509–530. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Leja, M.L.; Kaminskyy, D.V.; Kryshchyshyn, A.P.; Binduga, U.E.; Pinyazhko, O.R.; Lesyk, R.B.; Tobiasz, J.; Gmiński, J. Anticancer Properties of 4-Thiazolidinone Derivatives Depend on Peroxisome Proliferator-Activated Receptor Gamma (PPARγ). Eur. J. Med. Chem. 2017, 141, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Rothhammer, V.; Quintana, F.J. The Aryl Hydrocarbon Receptor: An Environmental Sensor Integrating Immune Responses in Health and Disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, T. Aryl Hydrocarbon Receptor Connects Inflammation to Breast Cancer. Int. J. Mol. Sci. 2020, 21, 5264. [Google Scholar] [CrossRef]
- Huang, Y.; He, J.; Liang, H.; Hu, K.; Jiang, S.; Yang, L.; Mei, S.; Zhu, X.; Yu, J.; Kijlstra, A.; et al. Aryl Hydrocarbon Receptor Regulates Apoptosis and Inflammation in a Murine Model of Experimental Autoimmune Uveitis. Front. Immunol. 2018, 9, 01713. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Sherr, D.H.; Monti, S. The Role of the Aryl Hydrocarbon Receptor in Normal and Malignant B Cell Development. Semin. Immunopathol. 2013, 35, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Gazourian, L.; Quadri, S.A.; R Romieu-Mourez, R.; Sherr, D.H.; Sonenshein, G.E. The RelA NF-KappaB Subunit and the Aryl Hydrocarbon Receptor (AhR) Cooperate to Transactivate the c-Myc Promoter in Mammary Cells. Oncogene 2000, 19, 5498–5506. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Chang, H.; Tsai, W.-T.; Wu, M.-H.; Liao, Y.-S.; Chen, J.-T.; Su, J.-M. Overexpression of Aryl Hydrocarbon Receptor in Human Lung Carcinomas. Toxicol. Pathol. 2003, 31, 22–30. [Google Scholar] [CrossRef]
- Chen, J.; Röcken, C.; Klein-Hitpass, L.; Götze, T.; Leodolter, A.; Malfertheiner, P.; Ebert, M.P.A. Microarray Analysis of Gene Expression in Metastatic Gastric Cancer Cells after Incubation with the Methylation Inhibitor 5-Aza-2’-Deoxycytidine. Clin. Exp. Metastasis 2004, 21, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Snyder, M.; Kenison, J.E.; Yang, K.; Lara, B.; Lydell, E.; Bennani, K.; Novikov, O.; Federico, A.; Monti, S.; et al. How the AHR Became Important in Cancer: The Role of Chronically Active AHR in Cancer Aggression. Int. J. Mol. Sci. 2021, 22, 387. [Google Scholar] [CrossRef]
- Dou, H.; Duan, Y.; Zhang, X.; Yu, Q.; Di, Q.; Song, Y.; Li, P.; Gong, Y. Aryl Hydrocarbon Receptor (AhR) Regulates Adipocyte Differentiation by Assembling CRL4B Ubiquitin Ligase to Target PPARγ for Proteasomal Degradation. J. Biol. Chem. 2019, 294, 18504–18515. [Google Scholar] [CrossRef] [PubMed]
- Szychowski, K.A.; Skóra, B.; Kryshchyshyn-Dylevych, A.; Kaminskyy, D.; Rybczyńska-Tkaczyk, K.; Lesyk, R.; Gmiński, J. Induction of Cyp450 Enzymes by 4-Thiazolidinone-Based Derivatives in 3T3-L1 Cells in Vitro. Naunyn. Schmiedebergs. Arch. Pharmacol. 2021, 394, 915–927. [Google Scholar] [CrossRef]
- Senkiv, J.; Finiuk, N.; Kaminskyy, D.; Havrylyuk, D.; Wojtyra, M.; Kril, I.; Gzella, A.; Stoika, R.; Lesyk, R. 5-Ene-4-Thiazolidinones Induce Apoptosis in Mammalian Leukemia Cells. Eur. J. Med. Chem. 2016, 117, 33–46. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Rombel-Bryzek, A.; Dołhańczuk-Śródka, A.; Gmiński, J. Antiproliferative Effect of Elastin-Derived Peptide VGVAPG on SH-SY5Y Neuroblastoma Cells. Neurotox. Res. 2019, 36, 503–514. [Google Scholar] [CrossRef]
- Nicholson, D.W.; Ali, A.; Thornberry, N.A.; Vaillancourt, J.P.; Ding, C.K.; Gallant, M.; Gareau, Y.; Griffin, P.R.; Labelle, M.; Lazebnik, Y.A.; et al. Identification and Inhibition of the ICE/CED-3 Protease Necessary for Mammalian Apoptosis. Nature 1995, 376, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Skóra, B.; Piechowiak, T.; Szychowski, K.A. Epidermal Growth Factor-Labeled Liposomes as a Way to Target the Toxicity of Silver Nanoparticles into EGFR-Overexpressing Cancer Cells in Vitro. Toxicol. Appl. Pharmacol. 2022, 443, 116009. [Google Scholar] [CrossRef] [PubMed]
- Szychowski, K.A.; Skóra, B.; Kryshchyshyn-Dylevych, A.; Kaminskyy, D.; Khyluk, D.; Lesyk, R. 4-Thiazolidinone-Based Derivatives Rosiglitazone and Pioglitazone Affect the Expression of Antioxidant Enzymes in Different Human Cell Lines. Biomed. Pharmacother. 2021, 139, 111684. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Finiuk, N.; Kaleniuk, E.; Holota, S.; Stoika, R.; Lesyk, R.; Szychowski, K.A. Pyrrolidinedione-Thiazolidinone Hybrid Molecules with Potent Cytotoxic Effect in Squamous Cell Carcinoma SCC-15 Cells. Bioorganic Med. Chem. 2023, 92, 117442. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Yamamoto, K.; Tamura, T.; Nakamura, R.; Hosoe, S.; Matsubara, M.; Nagata, K.; Kodaira, H.; Uemori, T.; Takahashi, Y.; Suzuki, M.; et al. Development of a Novel Class of Peroxisome Proliferator-Activated Receptor (PPAR) Gamma Ligands as an Anticancer Agent with a Unique Binding Mode Based on a Non-Thiazolidinedione Scaffold. Bioorg. Med. Chem. 2019, 27, 115122. [Google Scholar] [CrossRef] [PubMed]
- Gruszczyk, J.; Grandvuillemin, L.; Lai-Kee-Him, J.; Paloni, M.; Savva, C.G.; Germain, P.; Grimaldi, M.; Boulahtouf, A.; Kwong, H.-S.; Bous, J.; et al. Cryo-EM Structure of the Agonist-Bound Hsp90-XAP2-AHR Cytosolic Complex. Nat. Commun. 2022, 13, 7010. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An Advanced Semantic Chemical Editor, Visualization, and Analysis Platform. J. Cheminform. 2012, 4, 17. [Google Scholar] [CrossRef]
- Einstein, M.; Akiyama, T.E.; Castriota, G.A.; Wang, C.F.; McKeever, B.; Mosley, R.T.; Becker, J.W.; Moller, D.E.; Meinke, P.T.; Wood, H.B.; et al. The Differential Interactions of Peroxisome Proliferator-Activated Receptor γ Ligands with Tyr473 Is a Physical Basis for Their Unique Biological Activities. Mol. Pharmacol. 2008, 73, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Finiuk, N.; Boiko, N.; Klyuchivska, O.; Kobylinska, L.; Kril, I.; Zimenkovsky, B.; Lesyk, R.; Stoika, R. 4-Thiazolidinone Derivative Les-3833 Effectively Inhibits Viability of Human Melanoma Cells through Activating Apoptotic Mechanisms. Croat. Med. J. 2017, 58, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Kim, G.Y.; Li, W.; Choi, B.T.; Kim, N.D.; Kang, H.S.; Choi, Y.H. Implication of Intracellular ROS Formation, Caspase-3 Activation and Egr-1 Induction in Platycodon D-Induced Apoptosis of U937 Human Leukemia Cells. Biomed. Pharmacother. 2009, 63, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of Apoptosis Signalling Pathways by Reactive Oxygen Species. Biochim. Biophys. Acta—Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Eskandari, E.; Eaves, C.J. Paradoxical Roles of Caspase-3 in Regulating Cell Survival, Proliferation, and Tumorigenesis. J. Cell Biol. 2022, 221, e202201159. [Google Scholar] [CrossRef] [PubMed]
- Kobylinska, L.I.; Klyuchivska, O.Y.; Grytsyna, I.I.; Finiuk, N.; Panchuk, R.R.; Starykovych, M.O.; Lehka, L.; Lesyk, R.B.; Zimenkovsky, B.S.; Stoika, R.S. Differential Pro-Apoptotic Effects of Synthetic 4-Thiazolidinone Derivative Les-3288, Doxorubicin and Temozolomide in Human Glioma U251 Cells. Croat. Med. J. 2017, 58, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Cullen, S.P.; Martin, S.J. Caspase Activation Pathways: Some Recent Progress. Cell Death Differ. 2009, 16, 935–938. [Google Scholar] [CrossRef]
- Tickner, J.; Fan, L.M.; Du, J.; Meijles, D.; Li, J.M. Nox2-Derived ROS in PPARγ Signaling and Cell-Cycle Progression of Lung Alveolar Epithelial Cells. Free Radic. Biol. Med. 2011, 51, 763–772. [Google Scholar] [CrossRef]
- Grishanova, A.Y.; Perepechaeva, M.L. Aryl Hydrocarbon Receptor in Oxidative Stress as a Double Agent and Its Biological and Therapeutic Significance. Int. J. Mol. Sci. 2022, 23, 6719. [Google Scholar] [CrossRef]
- Kosińska, K.; Skóra, B.; Holota, S.; Shepeta, Y.; Tabęcka-Łonczyńska, A.; Lesyk, R.; Szychowski, K.A. Role of 4-Thiazolidinone–Pyrazoline/Indoline Hybrids Les-4369 and Les-3467 in BJ and A549 Cell Lines. Cells 2024, 13, 1007. [Google Scholar] [CrossRef] [PubMed]
- Szychowski, K.A.; Skóra, B.; Kryshchyshyn-Dylevych, A.; Kaminskyy, D.; Tobiasz, J.; Lesyk, R.B.; Gmiński, J. 4-Thiazolidinone-Based Derivatives Do Not Affect Differentiation of Mouse Embryo Fibroblasts (3T3-L1 Cell Line) into Adipocytes. Chem. Biol. Interact. 2021, 345, 109538. [Google Scholar] [CrossRef]
- Szlachcikowska, D.; Tabęcka-Łonczyńska, A.; Holota, S.; Roman, O.; Shepeta, Y.; Lesyk, R.; Szychowski, K.A. Role of Ciminalum-4-Thiazolidinone Hybrids in Molecular NF-ΚB Dependent Pathways. Int. J. Mol. Sci. 2024, 25, 7329. [Google Scholar] [CrossRef] [PubMed]
- Neri, T.; Armani, C.; Pegoli, A.; Cordazzo, C.; Carmazzi, Y.; Brunelleschi, S.; Bardelli, C.; Breschi, M.C.; Paggiaro, P.; Celi, A. Role of NF-ΚB and PPAR-γ in Lung Inflammation Induced by Monocyte-Derived Microparticles. Eur. Respir. J. 2011, 37, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Harrison, L.E. Ciglitazone Induces Early Cellular Proliferation and NF-ΚB Transcriptional Activity in Colon Cancer Cells through P65 Phosphorylation. Int. J. Biochem. Cell Biol. 2005, 37, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-P.; Yang, X.-N.; Song, Y.; Zhou, F.; Liu, J.-J.; Hu, Y.-Q.; Chen, L.-G. Rosiglitazone Alleviates Lipopolysaccharide-Induced Inflammation in RAW264.7 Cells via Inhibition of NF-ΚB and in a PPARγ-Dependent Manner. Exp. Ther. Med. 2021, 22, 10175. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Wu, T.; Li, G.; Gu, X.; Tian, Y.; Cui, H. Insights into the Critical Role of the PXR in Preventing Carcinogenesis and Chemotherapeutic Drug Resistance. Int. J. Biol. Sci. 2022, 18, 742. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Tabb, M.M.; Nelson, E.L.; Grün, F.; Verma, S.; Sadatrafiei, A.; Lin, M.; Mallick, S.; Forman, B.M.; Thummel, K.E.; et al. Mutual Repression between Steroid and Xenobiotic Receptor and NF-KappaB Signaling Pathways Links Xenobiotic Metabolism and Inflammation. J. Clin. Investig. 2006, 116, 2280–2289. [Google Scholar] [CrossRef] [PubMed]
- Wahli, W. A Gut Feeling of the PXR, PPAR and NF-ΚB Connection. J. Intern. Med. 2008, 263, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Jasper Deuring, J.; Li, M.; Cao, W.; Chen, S.; Wang, W.; de Haar, C.; Janneke van der Woude, C.; Peppelenbosch, M. Pregnane X Receptor Activation Constrains Mucosal NF-ΚB Activity in Active Inflammatory Bowel Disease. PLoS ONE 2019, 14, e0221924. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Tested Compound | Exposure Time | IC50 ( ± SD, µM) | LD50 ( ± SD, µM) |
|---|---|---|---|---|
| BJ | Les-4368 | 24 h | 27.82 ± 6.34 | 54.50 ± 9.94 |
| 48 h | 15.46 ± 7.83 | 19.95 ± 5.38 | ||
| Les-4370 | 24 h | N/A | 87.75 ± 4.65 | |
| 48 h | N/A | 15.82 ± 1.62 | ||
| CACO-2 | Les-4368 | 24 h | N/A | N/A |
| 48 h | 29.58 ± 9.45 | N/A | ||
| Les-4370 | 24 h | N/A | N/A | |
| 48 h | N/A | N/A |
| Compounds | PPARγ (PDB: 6K0T) | AhR (PDB: 7ZUB) |
|---|---|---|
| Binding Energy ∆G kcal/mol | Binding Energy ∆G kcal/mol | |
| Les-4368 | −9.15 | −3.06 |
| Les-4370 | −8.79 | 1.49 |
| CID 136264026 | −16.15 | - |
| Indirubin | - | −12.68 |
| Rosiglitazone | −8.08 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaleniuk, E.; Holota, S.; Skóra, B.; Khylyuk, D.; Tabęcka-Łonczyńska, A.; Lesyk, R.; Szychowski, K.A. Anticancer Properties of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Potential Agonists 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370 in Colorectal Adenocarcinoma Cells In Vitro. Appl. Sci. 2024, 14, 7692. https://doi.org/10.3390/app14177692
Kaleniuk E, Holota S, Skóra B, Khylyuk D, Tabęcka-Łonczyńska A, Lesyk R, Szychowski KA. Anticancer Properties of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Potential Agonists 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370 in Colorectal Adenocarcinoma Cells In Vitro. Applied Sciences. 2024; 14(17):7692. https://doi.org/10.3390/app14177692
Chicago/Turabian StyleKaleniuk, Edyta, Serhii Holota, Bartosz Skóra, Dmytro Khylyuk, Anna Tabęcka-Łonczyńska, Roman Lesyk, and Konrad A. Szychowski. 2024. "Anticancer Properties of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Potential Agonists 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370 in Colorectal Adenocarcinoma Cells In Vitro" Applied Sciences 14, no. 17: 7692. https://doi.org/10.3390/app14177692
APA StyleKaleniuk, E., Holota, S., Skóra, B., Khylyuk, D., Tabęcka-Łonczyńska, A., Lesyk, R., & Szychowski, K. A. (2024). Anticancer Properties of Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Potential Agonists 4-Thiazolidinone-Pyrazoline Hybrids Les-4368 and Les-4370 in Colorectal Adenocarcinoma Cells In Vitro. Applied Sciences, 14(17), 7692. https://doi.org/10.3390/app14177692