
Structural Characteristics of the Pine Stands on Degraded Lands in the South-East of Romania, in the Context of Climate Changes

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
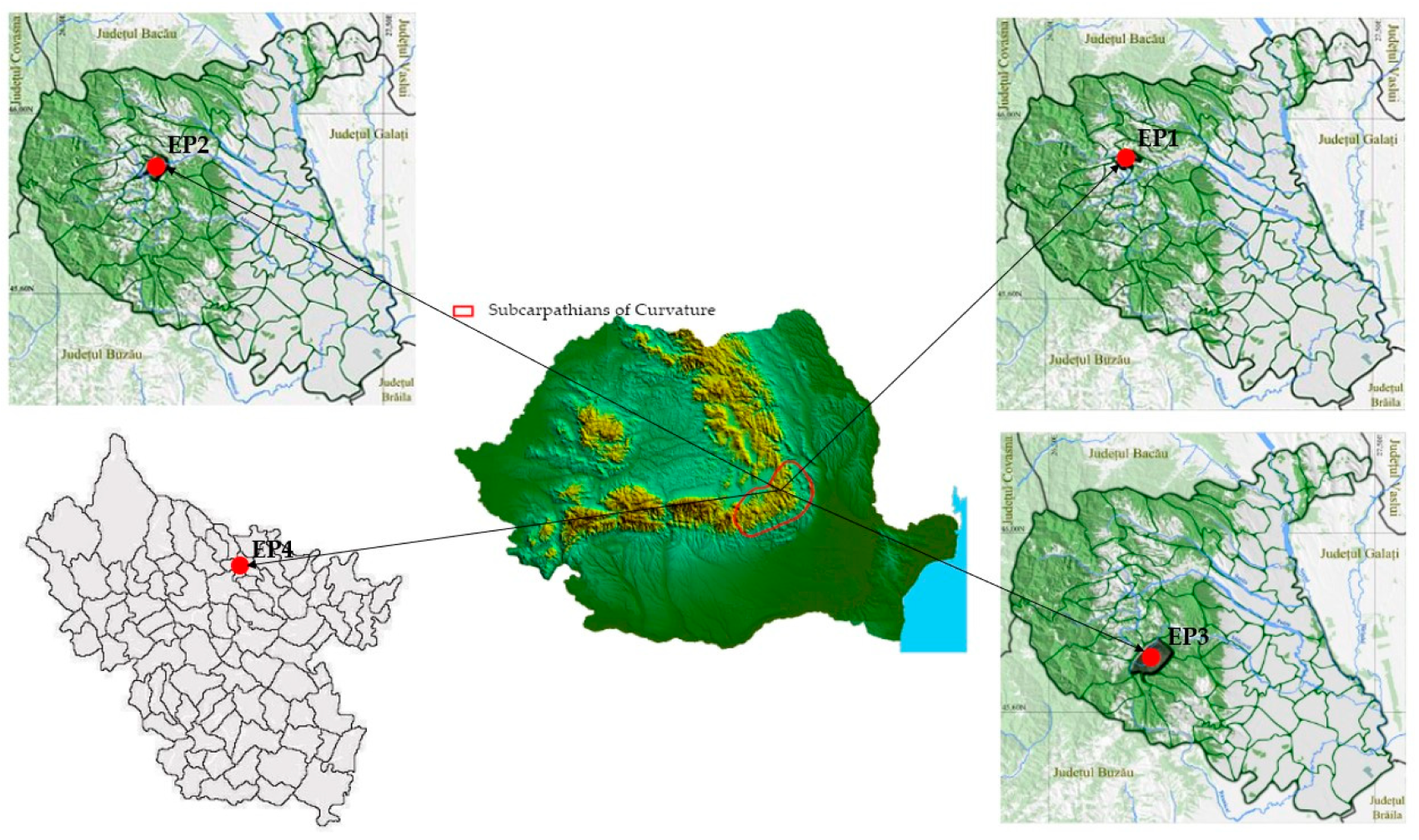
2.1. Data Collection
- (1)
- The sessile oak and common beech phytoclimatic storey (FD3): (i) Vidra Experimental Base, experimental perimeters: Caciu-Bârseşti (EP1) (45°55′19.69″ N, 26°44′44.74″ E) and Pârâul Sărat–Valea Sării (EP2) (45°52′35.57″ N, 26°47′51.97″ E); (ii) Focșani Forest District, experimental perimeter Roşoiu–Andreiaşu (EP3) (45°44′56.24″ N, 26°49′55.96″ E);
- (2)
- The internal hilly forest-steppe site (Ssd): (iii) Râmnicu-Sărat Forest District perimeter Livada–Râmnicu Sărat (EP4) (45°23′55.78″ N, 26°55′29.49″ E).
The Description of Degradation
2.2. Data Analysis
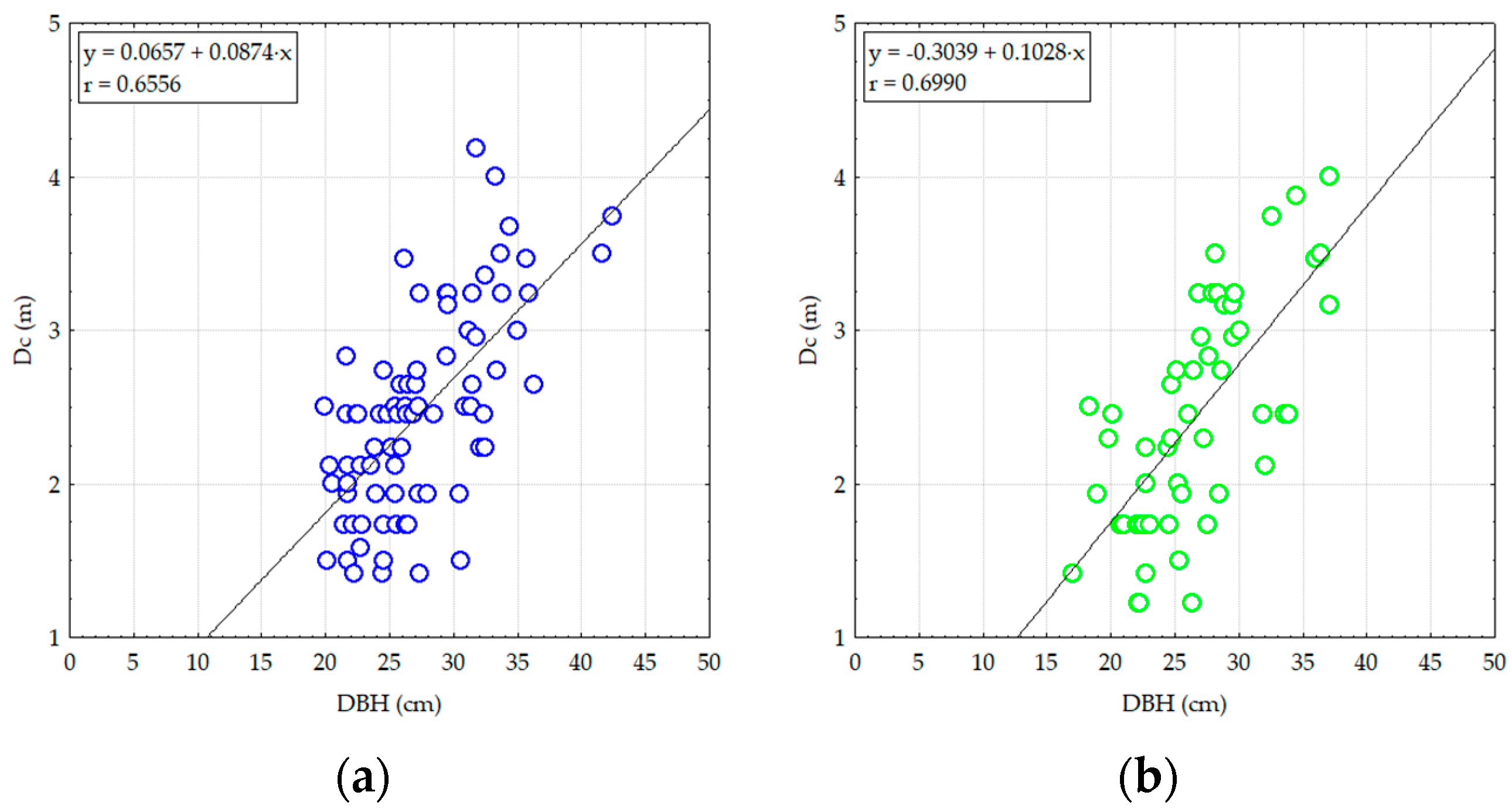
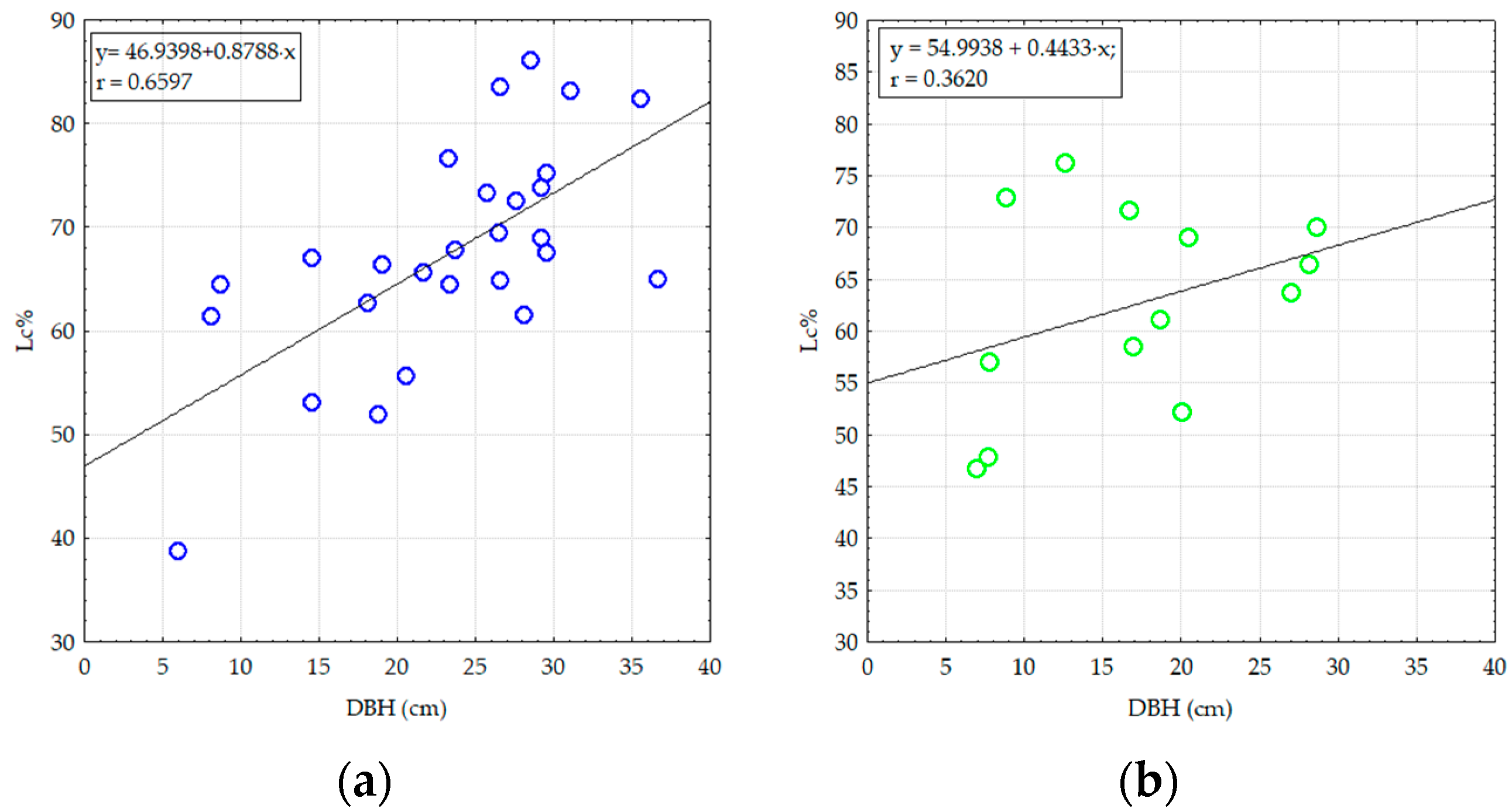
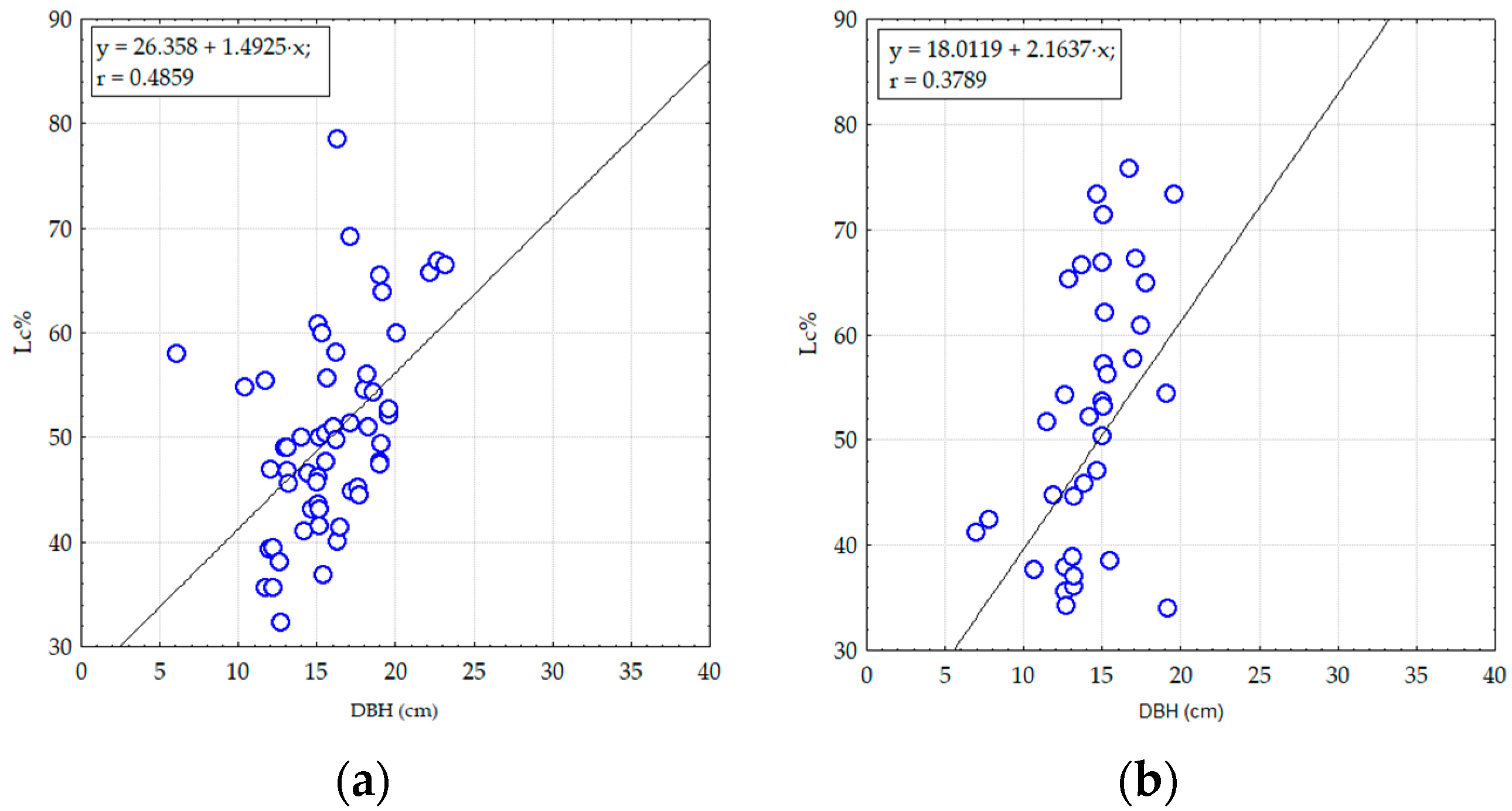
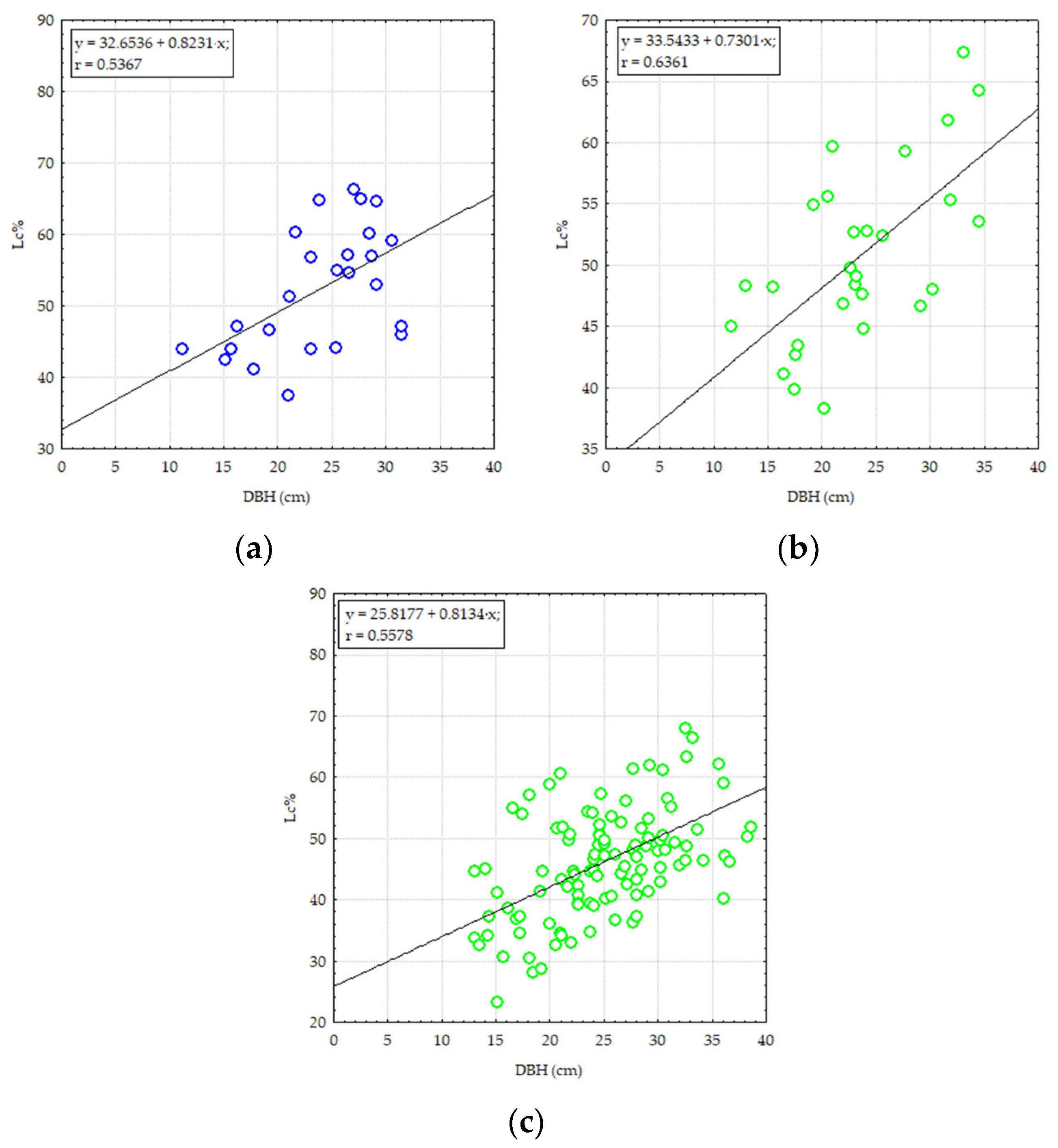
3. Results
3.1. Structural and Diversity Parameters of Pine Stands from Degraded Lands
3.2. The Effect of Damaging Factors on the Composition and Structure of Pine Stands on Degraded Lands
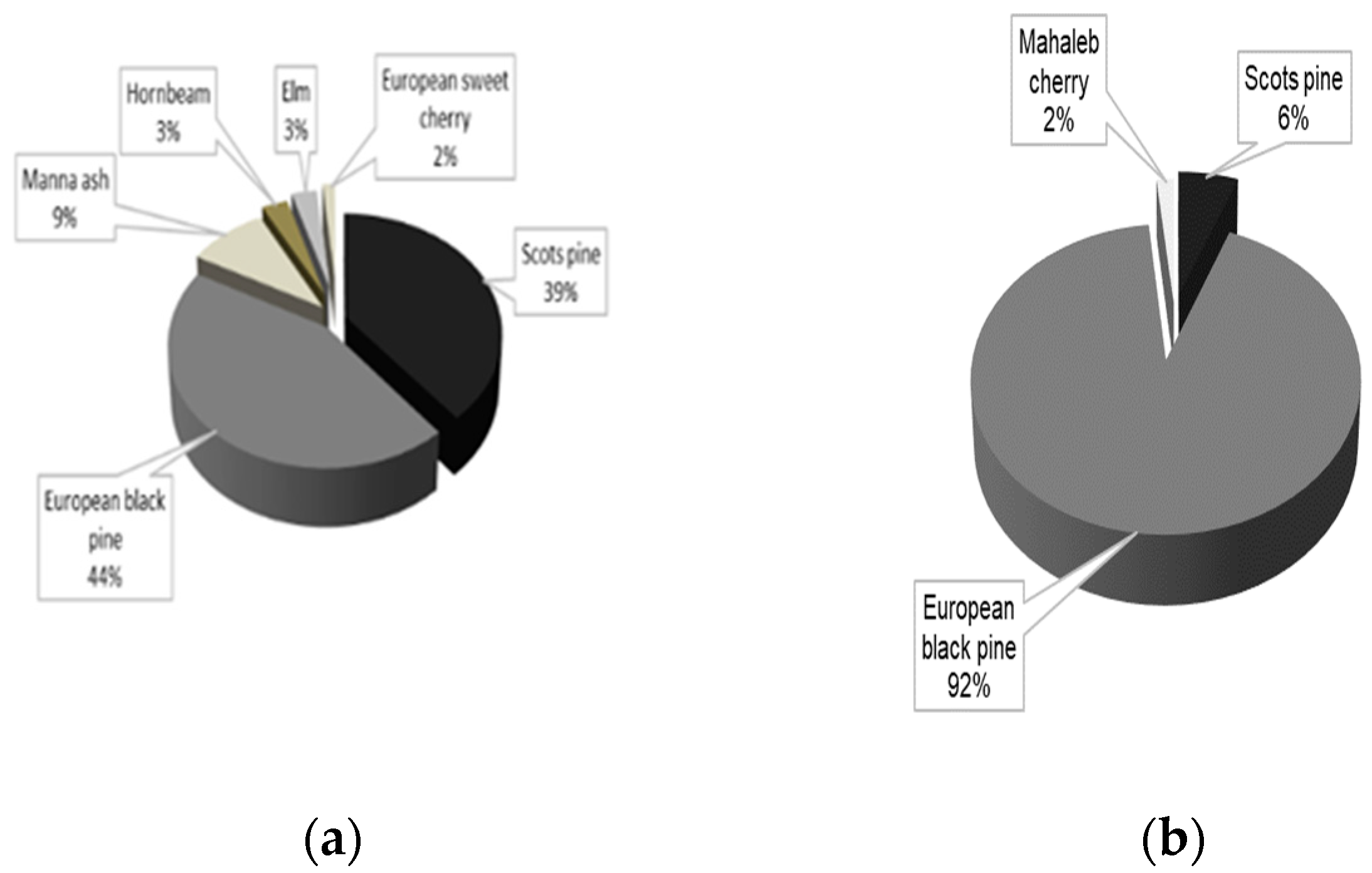
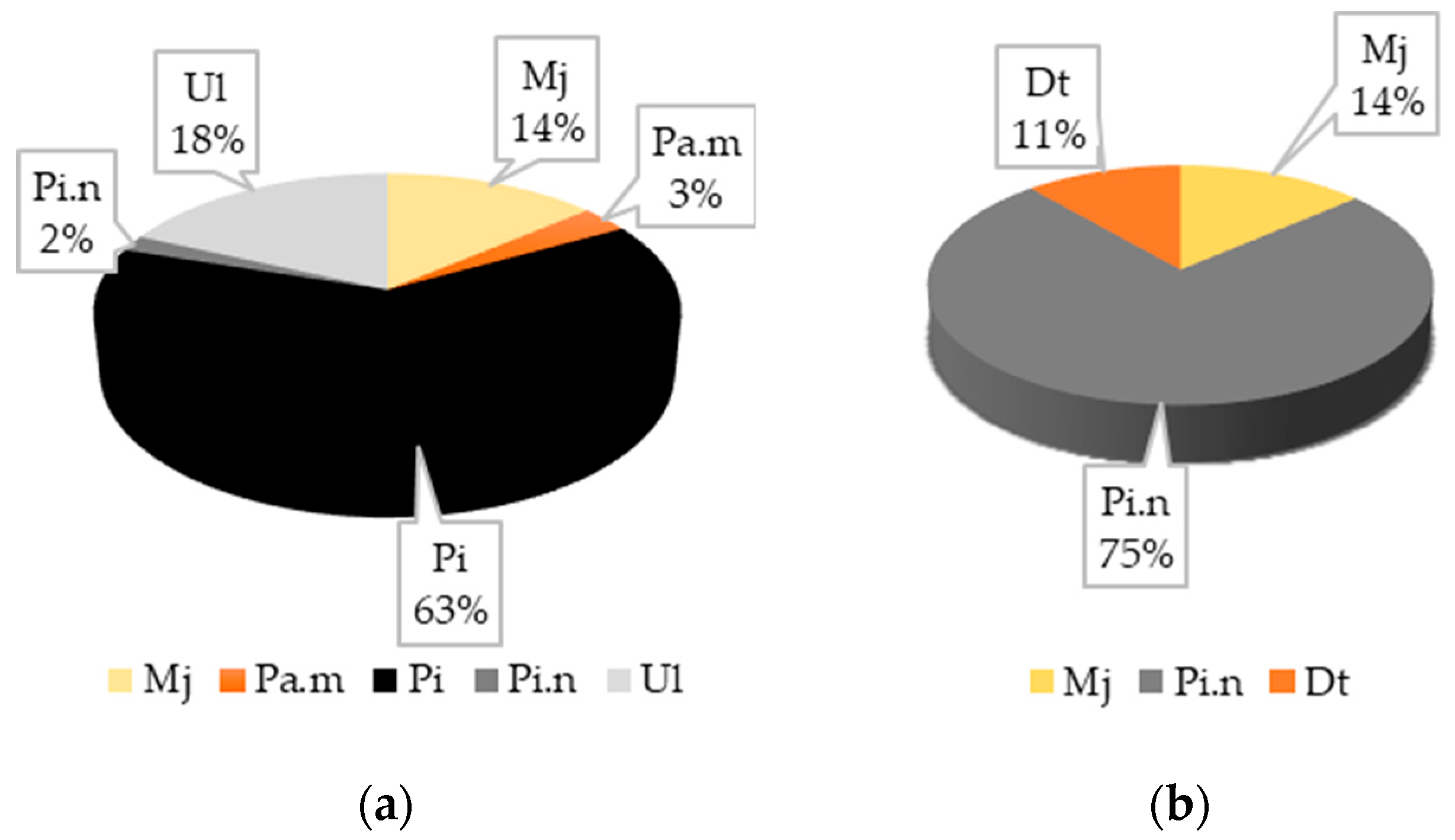
3.2.1. The Compositional Diversity of Pine Stands
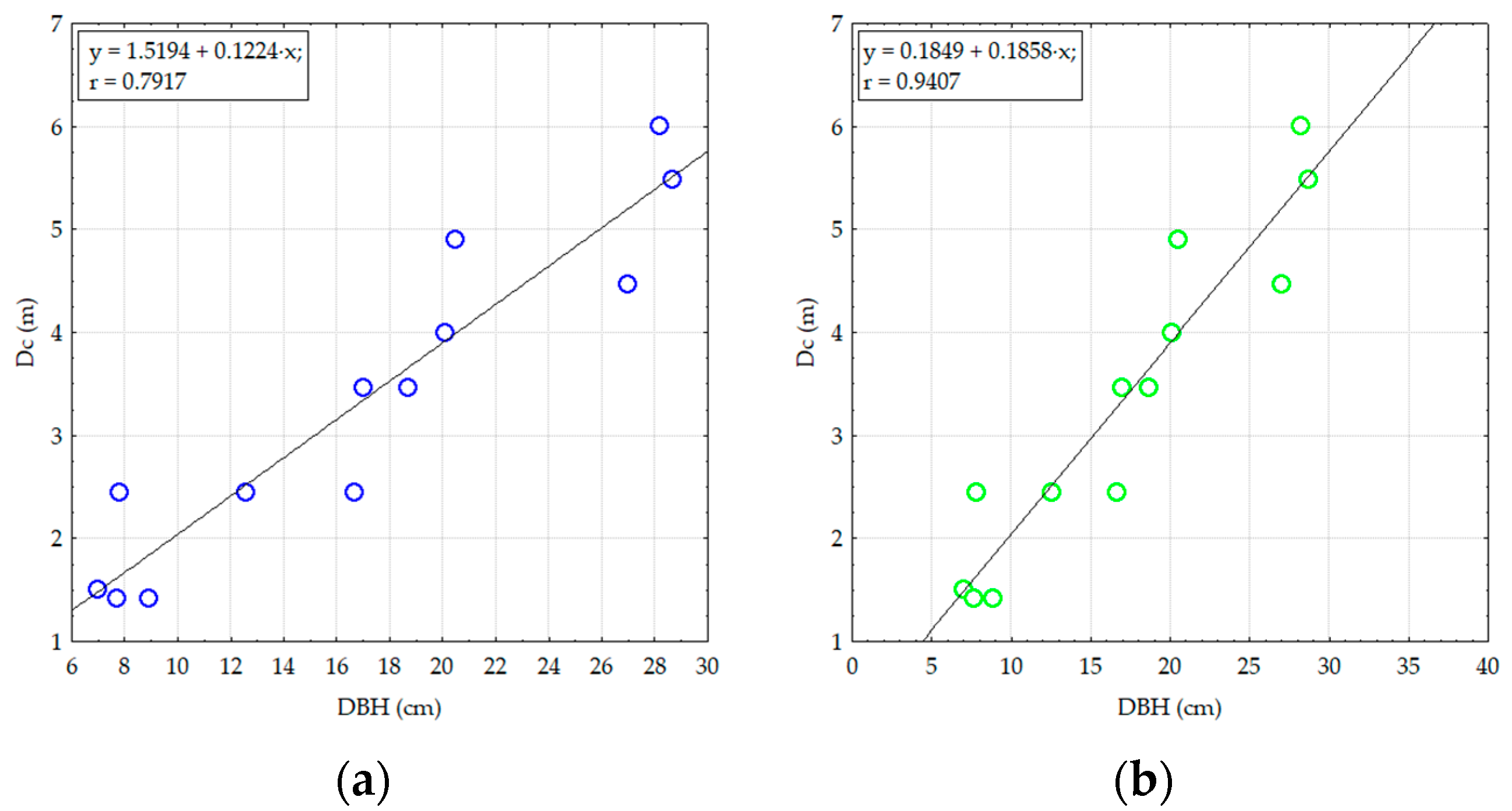
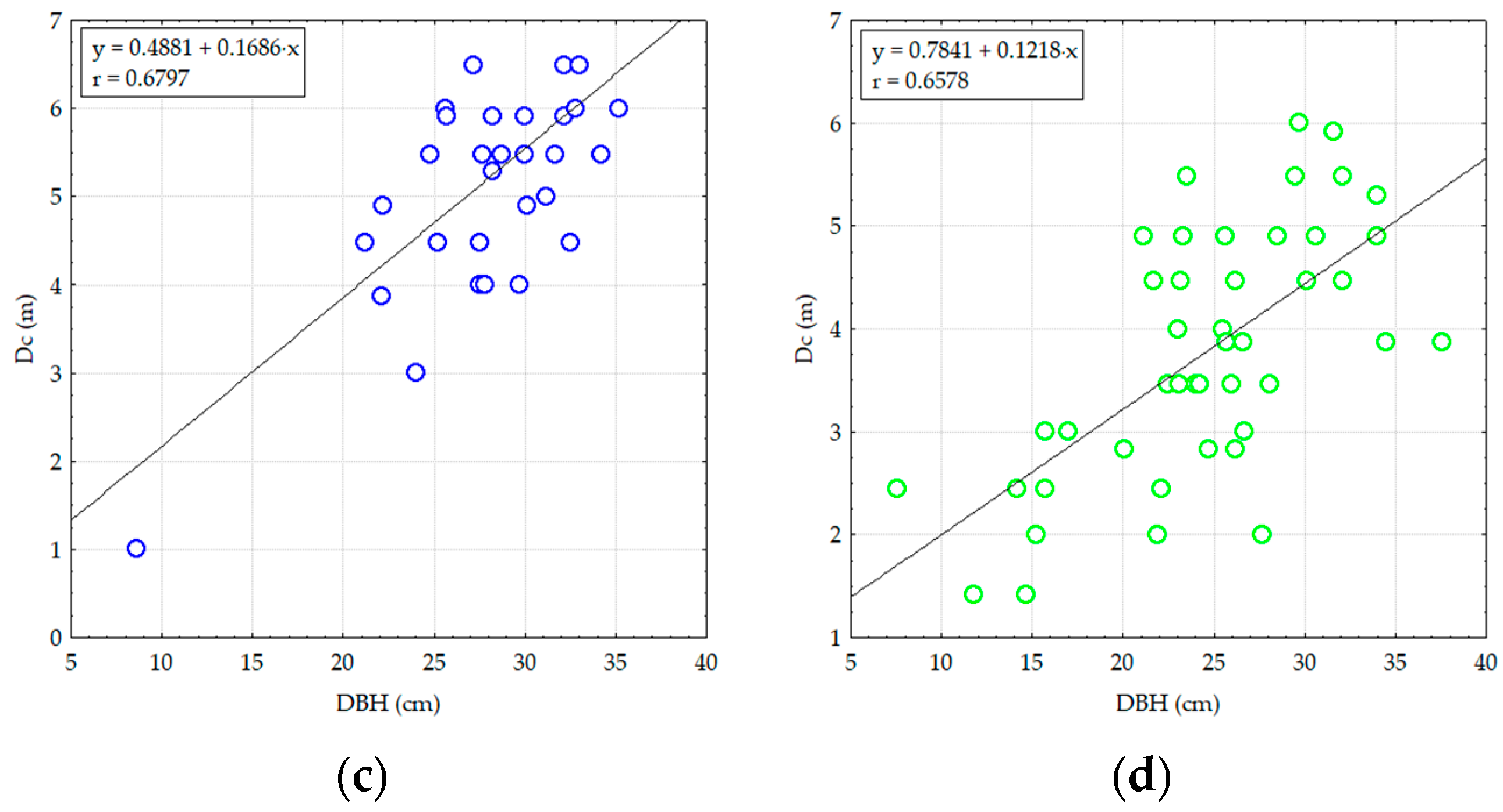
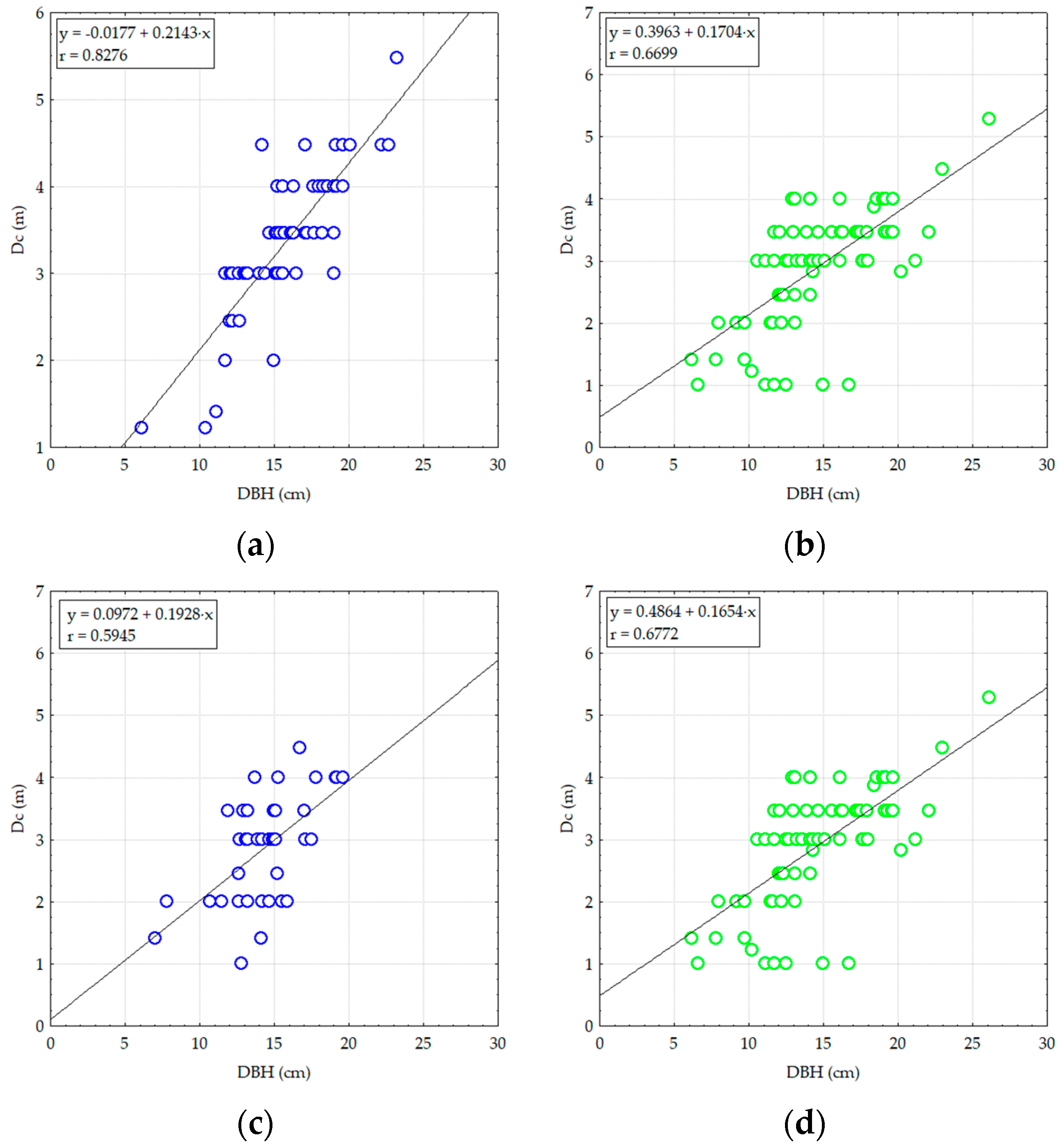
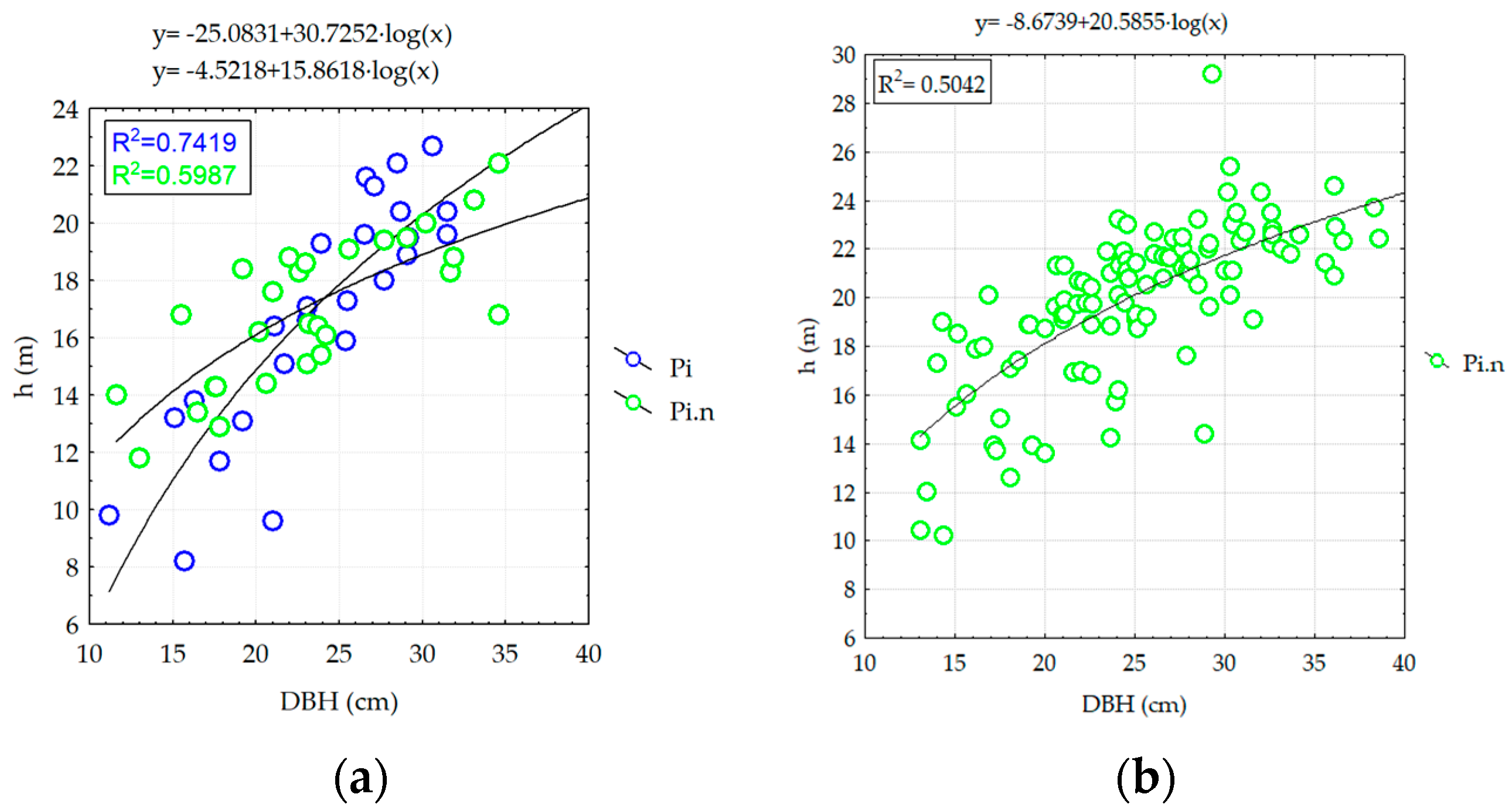
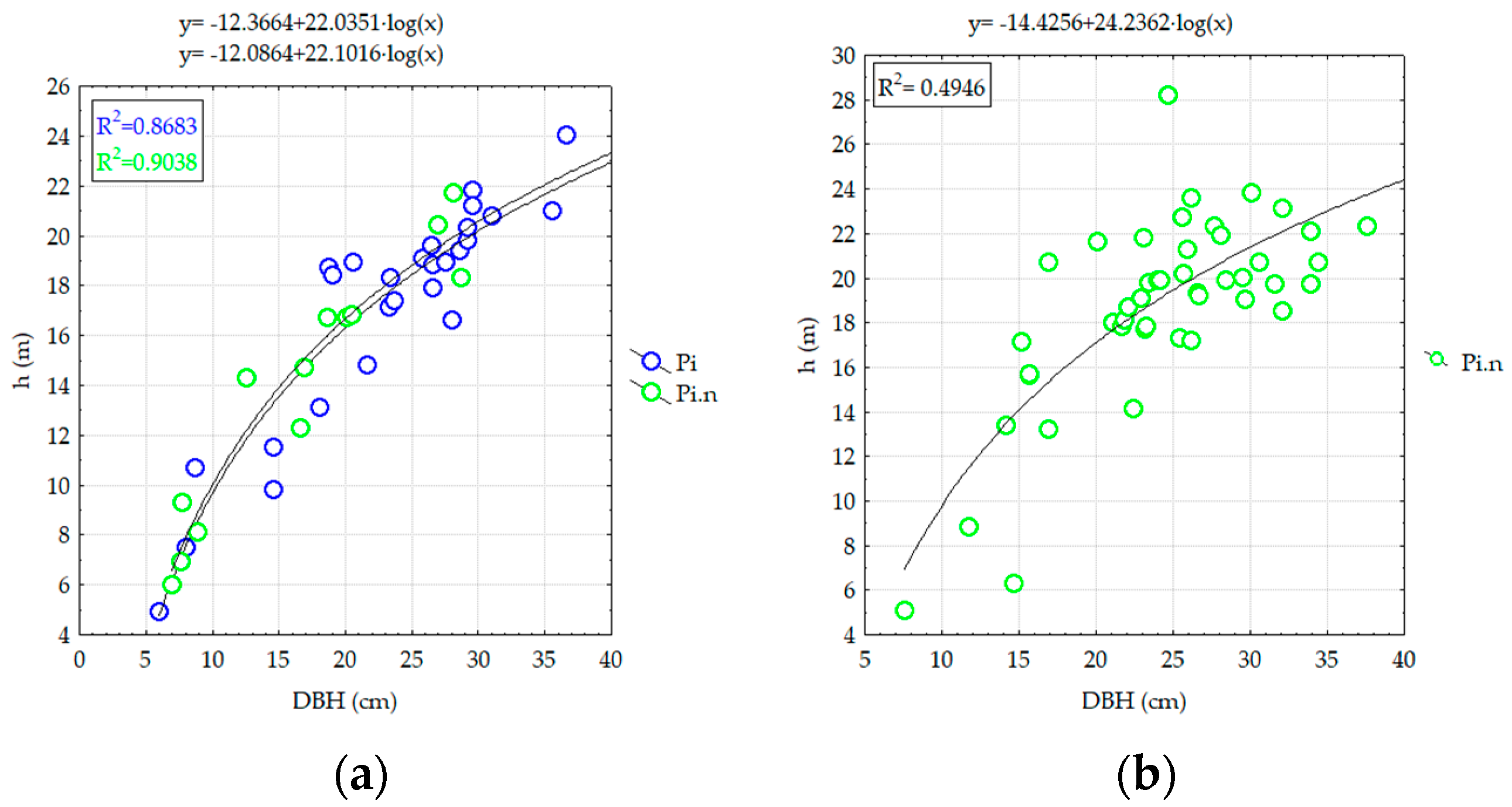
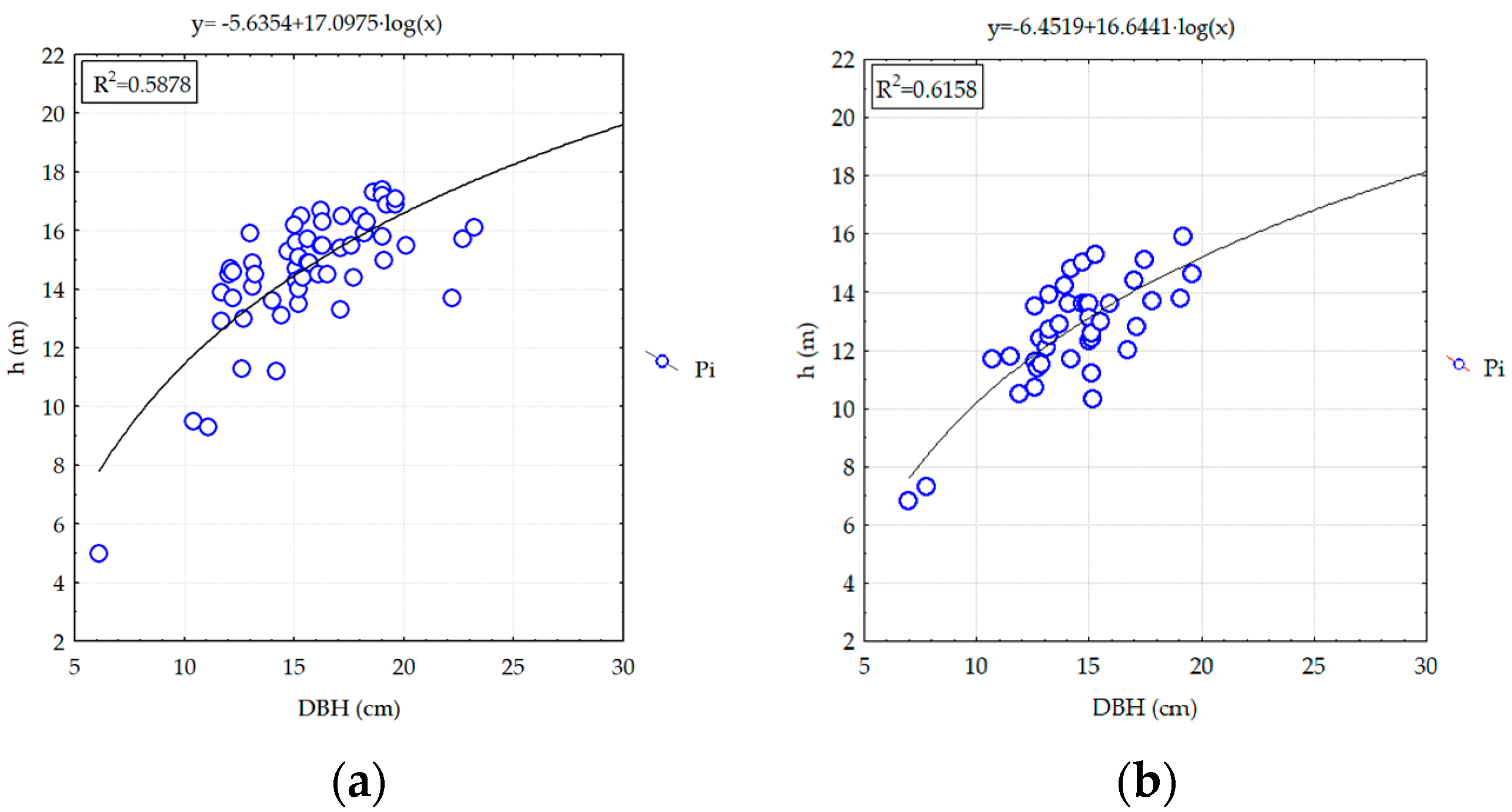
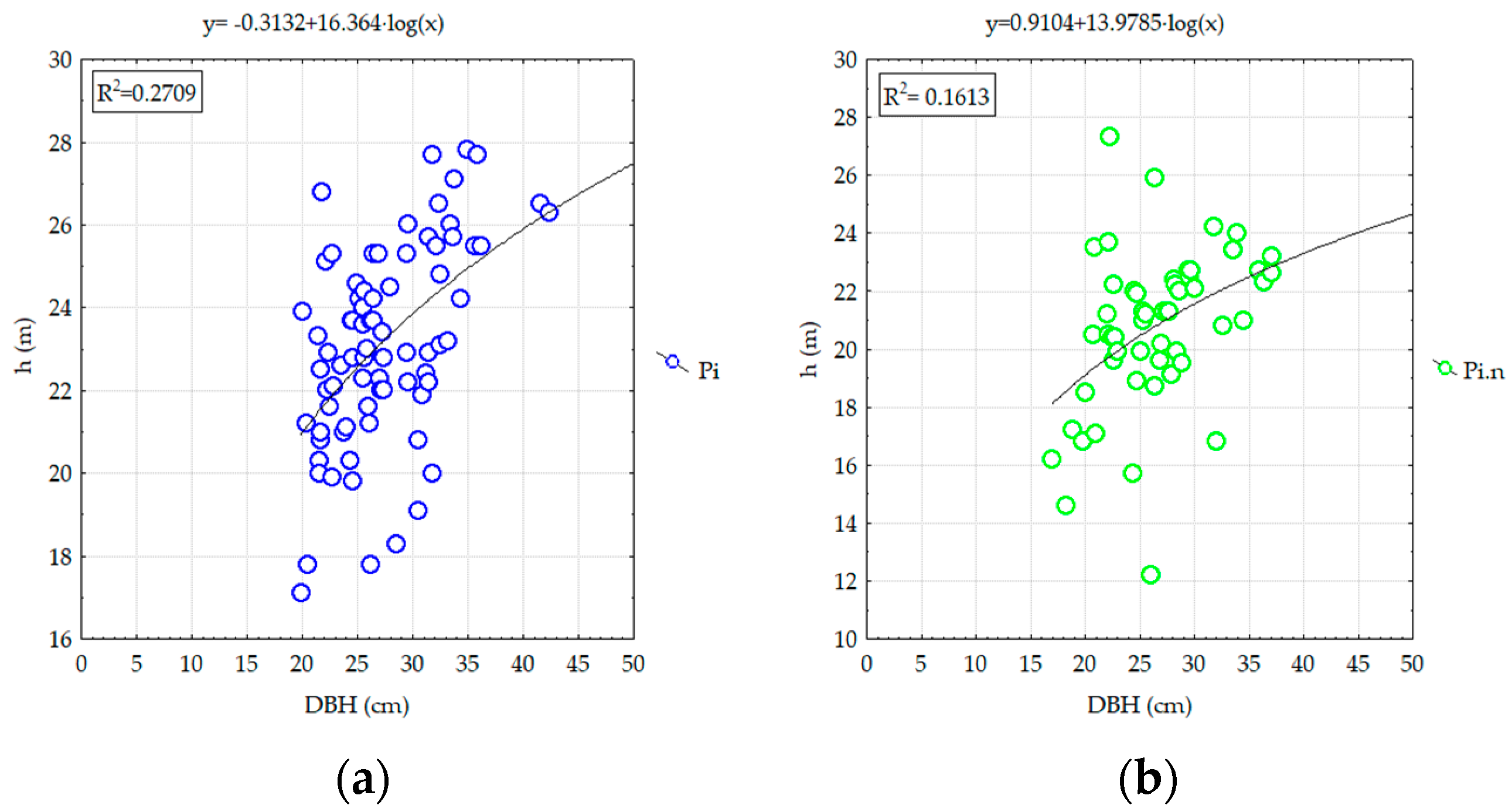
3.2.2. Analyzing the Stability of Scots Pine and Black Pine Stands
Pine Stands with Age between 55 and 65 Years (RP1 and RP9-EP2; RP4, RP9 and RP10-EP3) and over 70 Years (RP6 and RP9-EP4)
Pine Stands with Ages of 45 Years (RP5 and RP9-EP1)
4. Discussion
5. Conclusions
- The dynamic of structural parameters (diameter and height) in the forest cultures installed on degraded lands from the sessile oak–common beech phyto-climatic storey is significantly influenced by the complex of local harmful abiotic and biotic factors;
- The structural characteristics record significant differences firstly between the means of both the diameters and heights of its component species (pine and broadleaved), with the dimensions being superior for pine. Secondly, we found superior structural characteristics for Scots pine compared with black pine in identical environmental conditions;
- The evolution of stands from degraded lands goes in the direction of their diversification under a structural aspect. Stands composed of more species (mixtures of pine with broadleaved) are more stable than pure pine stands;
- In the case that we did not note the presence of species other than the ones initially used in creating the cultures, and when the influence of the abiotic factors was insignificant, the experimental distribution of trees into diameter classes followed the normal theoretical distribution and the beta theoretical frequency curve. In the case that more species were noted, the bilevel tendency of pine stands was evident through the observation of a new vegetation storey composed of local broadleaved species installed naturally;
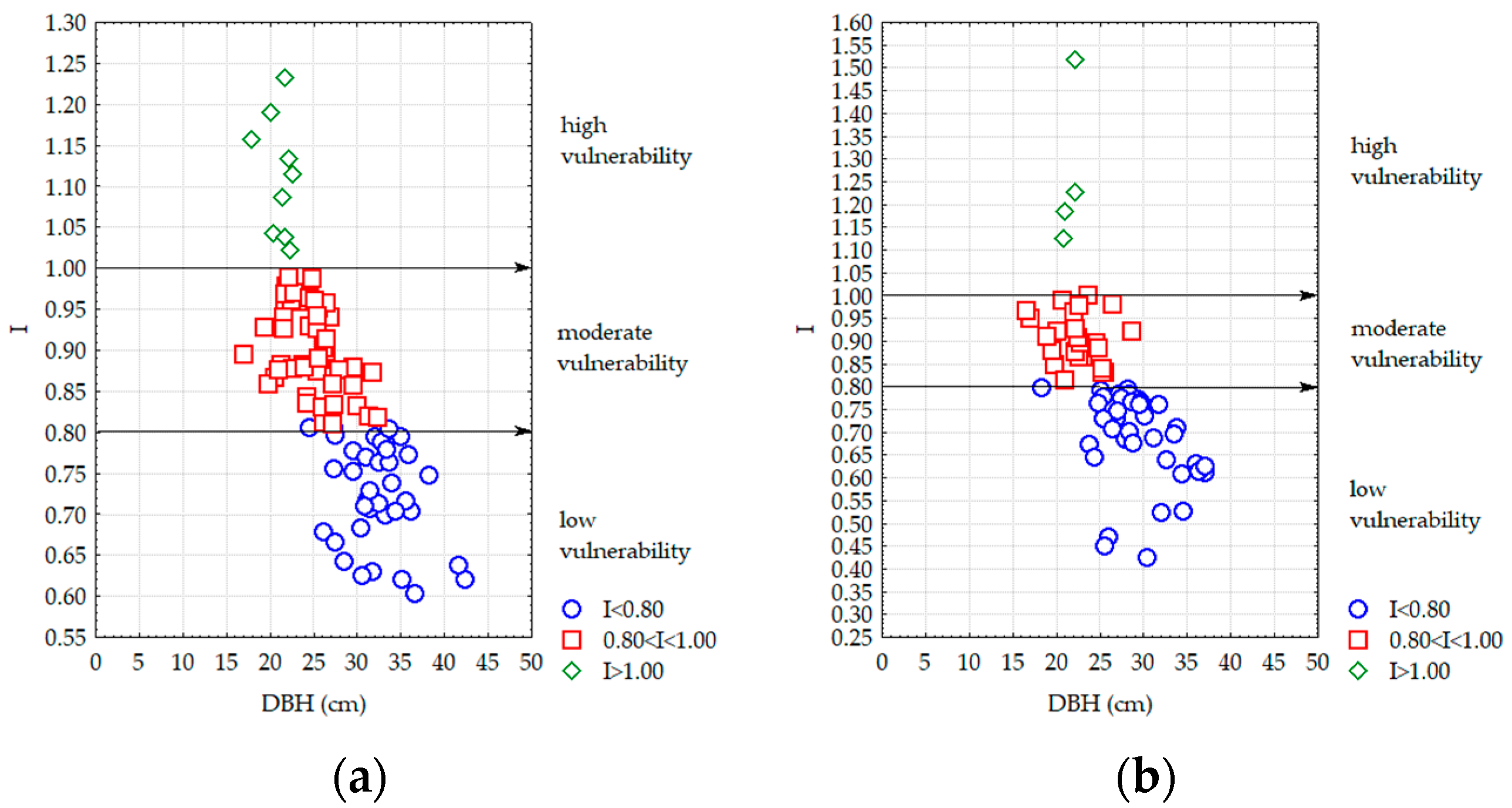
- From the perspective of stand stability, the main species (Scots pine or black pine) are situated in the moderate-vulnerability and high-vulnerability domain for young stands (40 years), with a high consistency and density. Stands with advanced ages (over 50 years) are characterized by a low vulnerability of pine species towards wind and snow, having reduced consistencies and densities;
- Much younger pine stands with diameters under 16 cm are situated in the moderate- and high-vulnerability domain due to the fact that they have a larger number of trees and have not yet been affected by wind and snow or other harmful factors;
- Stands over 50 years in age and with diameters higher than 22 cm are situated in the low-vulnerability domain because during their evolution, most of the pines from the vegetation storey were affected by snow and wind;
- Taking into account the harmful abiotic factors (wind, snow, etc.), in the moderate- and high-vulnerability domains, there are no significant differences between the means and variances of slenderness indexes. However, the statistical results showed significant differences between the means and variances at low vulnerability;
- Monitoring the evolution of pine stands by investigating the structure and diversity will lead to establishing the optimal moments for intervention with silvotechnical works in order to ensure the sustainable management of forest ecosystems from degraded lands.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dincă, L.; Achim, F. The management of forests situated on fields susceptible to landslides and erosion from the Southern Carpathians. Sci. Pap. Ser. Manag. Econ. Eng. Agric. Rural Dev. 2019, 19, 183–188. [Google Scholar]
- Janis, D.; Renate, S.; Oskars, K.; Edgars, D.; Mara, K.; Aris, J. A Financial Assessment of Windstorm Risks for Scots Pine Stands in Hemiboreal Forests. Forests 2020, 11, 566. [Google Scholar] [CrossRef]
- Silvestru-Grigore, C.V.; Dinulică, F.; Spârchez, G.; Hălălișan, A.F.; Dincă, L.; Enescu, R.; Crișan, V. The radial growth behaviour of pines (Pinus sylvestris L. and Pinus nigra Arn.) on Romanian degraded lands. Forests 2018, 9, 213. [Google Scholar] [CrossRef]
- Traci, C.; Untaru, E. Behavior and enhancement and consolidation effect of forestry plantation on degraded lands in experimental perimeters. In ICAS; 2nd series; Redacția de propagandă tehnică agricolă publishing House: Bucharest, Romania, 1986; p. 70. [Google Scholar]
- Dincă, L.; Murariu, G.; Enescu, C.M.; Achim, F.; Georgescu, L.; Murariu, A.; Timiș-Gânsac, V.; Holonec, L. Productivity differences between southern and northern slopes of Southern Carpathians (Romania) for Norway spruce, silver fir, birch and black alder. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 1070–1084. [Google Scholar] [CrossRef]
- Fedorca, A.; Popa, M.; Jurj, R.; Ionescu, G.; Ionescu, O.; Fedorca, M. Assessing the regional landscape connectivity for multispecies to coordinate on-the-ground needs for mitigating linear infrastructure impact in Brasov—Prahova region. J. Nat. Conserv. 2020, 58, 125903. [Google Scholar] [CrossRef]
- Lohmander, P.; Mohammadi, Z.; Kašpar, J.; Tahri, M.; Berčák, R.; Holuša, J.; Marušák, R. Future forest fires as functions of climate change and attack time for central Bohemian region, Czech Republic. Ann. For. Res. 2022, 65, 17–30. [Google Scholar] [CrossRef]
- Murariu, G.; Dinca, L.; Tudose, N.; Crisan, V.; Georgescu, L.; Munteanu, D.; Dragu, M.D.; Rosu, B.; Mocanu, G.D. Structural Characteristics of the Main Resinous Stands from Southern Carpathians, Romania. Forests 2021, 12, 1029. [Google Scholar] [CrossRef]
- Fedorca, A.; Fedorca, M.; Ionescu, O.; Jurj, R.; Ionescu, G.; Popa, M. Sustainable landscape planning to mitigate wildlife–vehicle collisions. Land 2021, 10, 737. [Google Scholar] [CrossRef]
- Portoghesi, L.; Tomao, A.; Bollati, S.; Mattioli, W.; Angelini, A.; Agrimi, M. Planning coastal Mediterranean stone pine (Pinus pinea L.) reforestations as a green infrastructure: Combining GIS techniques and statistical analysis to identify management options. Ann. For. Res. 2022, 65, 31–46. [Google Scholar] [CrossRef]
- Chmura, D.J.; Rożkowski, R.; Guzicka, M.; Dorobek, K. Variation in aboveground biomass carbon accumulation in Scots pine seed orchards progeny. Ann. For. Res. 2021, 64, 139–148. [Google Scholar] [CrossRef]
- Bigler, C.; Braker, O.U.; Bugmann, H.; Dobbertin, M.; Rigling, A. Drought as an Inciting Mortality Factor in Scots Pine stands of the Valais, Swizerland. Ecosystems 2006, 9, 330–343. [Google Scholar] [CrossRef]
- Ilek, A.; Nowak, M.; Błonska, E. Seasonal changes in water absorbability of some litterfall components in Scots pine stands differing in age. Ann. For. Res. 2021, 64, 149–164. [Google Scholar] [CrossRef]
- Constandache, C.; Peticilă, A.; Dincă, L.; Vasile, D. The usage of Sea Buckthorn (Hippophae rhamnoides L.) for improving Romania’s degraded lands. AgroLife Sci. J. 2016, 5, 50–58. [Google Scholar]
- Dinca, L.; Marin, M.; Vlad, R.; Murariu, G.; Drasovean, R.; Cretu, R.; Georgescu, L.; Timiș-Gânsac, V. Which are the Best Site and Stand Conditions for Silver Fir (Abies alba Mill.) Located in the Carpathian Mountains? Diversity 2022, 14, 547. [Google Scholar] [CrossRef]
- Constandache, C.; Dincă, L.; Tudor, C. The bioproductive potential of fast-growing forest species on degraded lands. Sci. Pap. Ser. E Land Reclam. Earth Obs. Surv. Environ. Eng. 2020, IX, 87–93. [Google Scholar]
- Tkach, V.; Tarnopilska, O.; Luk’yanets, V.; Musienko, S.; Kobets, O.; Rumiantsev, M.; Bondarenko, V. Density optimisation of pine plantations in the Left-Bank Steppe in Ukraine. Folia For. Pol. Ser. A—For. 2024, 66, 104–117. [Google Scholar] [CrossRef]
- Niţescu, C.; Achimescu, C. Technique of Silvicultural Crops. Tending and Management Operations of Stands; Ceres: Bucharest, Romania, 1979; p. 256. [Google Scholar]
- Sidor, C.G.; Camarero, J.J.; Popa, I.; Badea, O.; Apostol, E.N.; Vlad, R. Forest vulnerability to extreme climatic events in Romanian Scots pine forests. Sci. Total Environ. 2019, 678, 721–727. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmemann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Oberhuber, W.; Stumbock, M.; Kofler, W. Climate tree-growth relationships of Scots Pine Stands (Pinus sylvestris, L.), exposed to soil dryness. Trees 1998, 13, 19–27. [Google Scholar] [CrossRef]
- Rigling, A.; Waldner, P.O.; Forster, T.; Braker, O.U.; Pouttu, A. Ecological interpretation of tree-ring width and intraannual density fluctuations in Pinus sylvestris on dry sites in the central Alps and Siberia. Can. J. For. Res. 2001, 31, 18–31. [Google Scholar] [CrossRef]
- Rebetez, M.; Dobbertin, M. Climate change may already threaten scots pine stands in the Swiss Alps. Theor. Appl. Climatol. 2004, 79, 1–9. [Google Scholar] [CrossRef]
- Weber, P.; Bugmann, H.; Rigling, A. Radial growth responses to drought of Pinus Sylvestris and Quercus pubescens in an inner- Alpine dry valley. J. Veg. Sci. 2007, 18, 777–792. [Google Scholar]
- Pichler, P.; Oberhuber, W. Radial growth response of coniferous forest trees in inner Alpine environment to heat-wave in 2003. For. Ecol. Manag. 2007, 242, 688–699. [Google Scholar] [CrossRef]
- Petrescu, L. Guide for Tending Operations of Stands; Ceres publishing House: Bucharest, Romania, 1971. [Google Scholar]
- Petrescu, L.; Haring, P. Periodicity and Intensity of Cleanings and Thinnings in Spruce and Scots Pine Stands; Foresty Publishing: Bucharest, Romania, 1977; pp. 159–164. [Google Scholar]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of drought tolerance inmajor European temperate forest trees: Climatic drivers and levels of variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef]
- Szmyt, J.; Dobrowolska, D. Spatial diversity of forest regeneration after catastrophic wind in northeastern Poland. iForest 2015, 9, 414–421. [Google Scholar] [CrossRef]
- Schindler, D.; Kolbe, S. Assessment of the Response of a Scots Pine Tree to Effective Wind Loading. Forests 2020, 11, 145. [Google Scholar] [CrossRef]
- Leahu, I. Dendrometry; Didactical and Pedagogical Publishing House: Bucharest, Romania, 1994; p. 374. [Google Scholar]
- Constandache, C.; Tudor, C.; Popovici, L.; Vlad, R. The state and behavior of some forestry cultures installed on degraded lands in the forest-steppe site. AgroLife Sci. J. Sci. Pap. Ser. E Land Reclam. Earth Obs. Surv. Environ. Eng. 2023, XII, 215–223. [Google Scholar]
- Díaz-Yáñez, O.; Mola-Yudego, B.; González-Olabarria, J.R.; Pukkala, T. How does forest composition and structure affect the stability against wind and snow? For. Ecol. Manag. 2017, 401, 215–222. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Crecente-Campo, F.; Álvarez-Álvarez, P.; Barrio-Anta, M. Development of a stand density management diagram for radiata pine stands including assessment of stand stability. Forestry 2009, 82, 1–16. [Google Scholar] [CrossRef]
- Kamimura, K.; Gardiner, B.; Dupont, S.; Guyon, D.; Meredieu, C. Mechanistic and statistical approaches to predicting wind damage to individual maritime pine (Pinus pinaster) trees in forests. Can. J. For. Res. 2016, 46, 88–100. [Google Scholar] [CrossRef]
- Brichta, J.; Vacek, S.; Vacek, Z.; Cukor, J.; Mikeska, M.; Bílek, L.; Šimůnek, V.; Gallo, J.; Brabec, P. Importance and potential of Scots pine (L.) in 21 century. Cent. Eur. For. J. 2023, 69, 3–20. [Google Scholar]
- Vlad, R.; Constandache, C.; Dincă, L.; Tudose, N.C.; Sidor, C.G.; Popovici, L.; Ispravnic, A. Influence of climatic, site and stand characteristics on some structural parameters of scots pine (Pinus sylvestris) forests situated on degraded lands from East Romania. Range Mgmt. Agrofor. 2019, 40, 40–48. [Google Scholar]
- de Wergifosse, L. Simulating Tree Growth Response to Climate Change in Structurally-Complex Oak and Beech Stands across Europe. UCLouvain, Louvain-la-Neuve. 2021. Available online: https://dial.uclouvain.be/pr/boreal/object/boreal%3A250188/datastream/PDF_02/view (accessed on 1 March 2024).
- Dang, H.; Han, H.; Chen, S.; Li, M. A fragile soil moisture environment exacerbates the climate change-related impacts on the water use by Mongolian Scots pine (Pinus sylvestris var. mongolica) in northern China: Long-term observations. Agric. Water Manag. 2021, 251, 106857. [Google Scholar] [CrossRef]
- Belokopytova, L.V.; Zhirnova, D.F.; Krutovsky, K.V.; Mapitov, N.B.; Vaganov, E.A.; Babushkina, E.A. Species-and age-specific growth reactions to extreme droughts of the keystone tree species across forest-steppe and sub-taiga habitats of South Siberia. Forests 2022, 13, 1027. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Miller, C.L.; Parks, S.A.; Povak, N.A.; Taylor, A.H.; Higuera, P.E.; Prichard, S.J.; North, M.P.; Collins, B.M.; Hurteau, M.D.; et al. Climate, environment, and disturbance history govern resilience of western North American forests. Front. Ecol. Evol. 2019, 7, 239. [Google Scholar] [CrossRef]
- Pretzsch, H.; del Río, M.; Ammer, C.; Avdagic, A.; Barbeito, I.; Bielak, K.; Brazaitis, G.; Coll, L.; Dirnberger, G.; Drössler, L.; et al. Growth and yield of mixed versus pure stands of Scots pine (Pinus sylvestris L.) and European beech (Fagus sylvatica L.) analysed along a productivity gradient through Europe. Eur. J. For. Res. 2015, 134, 927–947. [Google Scholar] [CrossRef]
- Korhonen, K.T.; Ahola, A.; Heikkinen, J.; Henttonen, H.M.; Hotanen, J.P.; Ihalainen, A.; Melin, M.; Pitkänen, J.; Räty, M.; Sirviö, M.; et al. Forests of Finland 2014–2018 and their development 1921–2018. Silva Fenn. 2021, 55, 10662. [Google Scholar] [CrossRef]
- Arets, E.J.M.M.; Van der Meer, P.J.; Verwer, C.C.; Hengeveld, G.M.; Tolkamp, G.W.; Nabuurs, G.J.; Van Oorschot, M. Global Wood Production: Assessment of Industrial Round Wood Supply from Forest Management Systems in Different Global Regions; (Alterra-report; No. 1808); Alterra: Wageningen, The Netherlands, 2011; Available online: https://edepot.wur.nl/196265 (accessed on 27 June 2024).
- Güner, Ş.T.; Makineci, E. Determination of annual organic carbon sequestration in soil and forest floor of Scots pine forests on The Türkmen Mountain (Eskişehir, Kütahya). J. Fac. For. Istanb. Univ. 2017, 67, 109–115. [Google Scholar]
- Mäkelä, A.; Vanninen, P. Vertical structure of Scots pine crowns in different age and size classes. Trees 2001, 15, 385–392. [Google Scholar] [CrossRef]
- Pretzsch, H.; Rais, A. Wood quality in complex forests versus even-aged monocultures: Review and perspectives. Wood Sci. Technol. 2016, 50, 845–880. [Google Scholar] [CrossRef]
- Andreu, L.; Gutierrez, E.; Macias, M.; Ribas, M.; Bosch, O.; Camarero, J.J. Climate increases regional tree-growth variability in Iberian pine forests. Glob. Chang. Biol. 2007, 13, 804–815. [Google Scholar] [CrossRef]
- Pretzsch, H.; Pretzsch, H. Forest Dynamics, Growth, and Yield: A Review, Analysis of the Present State, and Perspective; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–39. [Google Scholar]
- Diaci, J.; Rozenbergar, D.; Anic, I.; Mikac, S.; Saniga, M.; Kucbel, S.; Visnjic, C.; Ballian, D. Structural dynamics and synchronous silver fir decline in mixed old-growth mountain forests in Eastern and Southeastern Europe. Forestry 2011, 84, 479–491. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Perimeter (EP) | Research Plot (RP) | * Degradation Form | Plantation Scheme (m) | Plantation Composition | Age | ** Actual Composition |
|---|---|---|---|---|---|---|
| EP1 | 5 | E3 | 1.50 × 3.00 0.50 × 3.00 | 50 Pi (Pi.n) 50 Ct.a | 45 | 55 Pi.n + 42 Pi + 3 An.a |
| 9 | E3 | 1.25 × 2.50 | 50 Pi (Pi.n) | 45 | 73 Pi.n + 26 Pi + 1 Sc | |
| 0.33 × 2.50 | 50 Ct.a | |||||
| EP2 | 1 | E3 | 1.00 × 1.00 | 70 Pi (Pi.n) 30 Mj, Ci, arb | 62 | 39 Pi + 44 Pi.n + 13 Dt |
| 9 | E1 | 1.00 × 1.00 | 60 Pi.n (Pi) 40 Vi.t, arb | 65 | 92 Pi.n + 6 Pi + 2 Vi.t | |
| EP3 | 4 | E2 | 1.00 × 1.00 | 70 Pi (Pi.n) 20 Pa 10 arb | 58 | 66 Pi + 32 Pi.n + 2 Ci |
| 9 | E3 | 1.00 × 3.00 | 50 Pi.n 50 Ct.a | 58 | 74 Pi.n + 13 Ar.t + 8 Pi + 5 Ci | |
| 10 | E3 | 1.00 × 3.00 | 50 Pi 50 Ct.a | 57 | 83 Pi + 17 Dt | |
| EP4 | 6 | E2 | 1.00 × 1.00 | Pi ± Pi.n, Ul, Lc, Mc | 75 | 63 Pi + 37 Dt |
| 9 | E1 | 1.00 × 1.00 | R1 = Pi.n R2 = Sg | 72 | 75 Pi.n + 25 Dt | |
| 12 | E1 | 2.00 × 1.00 | R1 = Pi.n + St.b. (St) R2 = Mc; R4 = Sg | 73 | 95 Pi.n + 5 Dt |
| Degree of Erosion | Eroded Land Category | Changes to Soils with Type Profile... | |||
|---|---|---|---|---|---|
| Qualifying | Symbol | A-AC-C | A(-AB)-Bv(Bt)-C | A-E-EB (E + B)-Bt-C | |
| Moderate erosion | E1 | Moderately eroded land | Up to 50% of the A horizon has been eroded | Up to 50% of the A horizon has been eroded | The A horizon has partially or totally eroded |
| High erosion | E2 | Heavily eroded land | More than 50% of the A horizon and up to 50% of the AC horizon has been eroded | More than 50% of the A horizon and up to 50% of the B horizon has been eroded | The E horizon has partially or totally eroded, and the EB or E + B horizon has partially or fully eroded. Parts of horizon B appear above |
| Very high erosion | E3 | Very heavily eroded land | The erosive action of water takes place in the lower half of the AC transition horizon or at the level of the C horizon | The erosive action takes place in the B horizon or in the C horizon | Erosion affects B horizon or C horizon |
| EP | RP | Sp | d ̅g | h ̅g | N·ha−1 | BA·ha−1 | COP | IN | IG | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 5 | Pi | 16.41 | 15.14 | 1324 | 28.08 | 3 | 0.86 | 0.82 | 2.93 | 45.35 |
| Pi.n | 15.16 | 13.71 | 1712 | 30.82 | 3 | 1.12 | 0.99 | 3.35 | 49.87 | ||
| 9 | Pi | 14.12 | 12.78 | 514 | 8.04 | 3 | 0.42 | 0.23 | 2.86 | 49.06 | |
| Pi.n | 12.60 | 10.68 | 1448 | 18.05 | 4 | 1.01 | 0.73 | 2.62 | 56.29 | ||
| 2 | 1 | Pi | 24.76 | 17.59 | 361 | 17.39 | 3 | 0.38 | 0.44 | 3.92 | 52.31 |
| Pi.n | 24.27 | 17.44 | 405 | 18.70 | 3 | 0.43 | 0.52 | 3.67 | 50.61 | ||
| 9 | Pi | 30.95 | 22.07 | 89 | 6.71 | 4 | 0.11 | 0.15 | 4.21 | 49.54 | |
| Pi.n | 26.03 | 20.40 | 1401 | 74.53 | 4 | 1.41 | 2.02 | 3.88 | 46.42 | ||
| 3 | 4 | Pi | 24.72 | 18.33 | 415 | 19.92 | 3 | 0.42 | 0.51 | 4.38 | 67.49 |
| Pi.n | 18.09 | 15.52 | 200 | 5.14 | 3 | 0.20 | 0.15 | 3.34 | 62.53 | ||
| 9 | Pi | 25.07 | 18.99 | 56 | 2.74 | 2 | 0.10 | 0.10 | 4.01 | 68.46 | |
| Pi.n | 25.20 | 19.59 | 500 | 24.92 | 2 | 0.52 | 0.6 | 3.75 | 58.05 | ||
| 10 | Pi | 28.52 | 20.29 | 388 | 24.79 | 2 | 0.45 | 0.57 | 5.22 | 57.95 | |
| 4 | 6 | Pi | 27.75 | 22.99 | 647 | 39.14 | 3 | 0.72 | 0.93 | 2.47 | 39.08 |
| 9 | Pi.n | 26.77 | 20.76 | 930 | 52.28 | 3 | 0.99 | 1.39 | 2.43 | 41.64 |
| EP-RP Species | Statistical Parameters | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Basal Diameter (cm) | Total Height (m) | |||||||||||
| Median | s | cv (%) | Min | Max | Median | s | cv (%) | Min | Max | |||
| 3–4 total | 21.70 | 23.30 | 8.07 | 37.22 | 7 | 37 | 16.34 | 17.90 | 4.64 | 28.43 | 6 | 24 |
| Pi | 24.05 | 26.15 | 7.22 | 30.02 | 8 | 37 | 17.51 | 18.75 | 3.99 | 22.81 | 8 | 24 |
| Pi.n | 17.82 | 17.85 | 7.59 | 42.63 | 8 | 29 | 14.68 | 15.70 | 4.73 | 32.23 | 7 | 22 |
| 3–9 total | 22.39 | 23.25 | 7.72 | 34.49 | 8 | 38 | 17.08 | 19.05 | 5.54 | 32.46 | 5 | 28 |
| Ar.t | 12.95 | 13.00 | 5.01 | 38.67 | 8 | 22 | 8.80 | 7.10 | 4.88 | 55.49 | 5 | 19 |
| Ci | 14.40 | 14.40 | 5.94 | 41.25 | 10 | 19 | 12.35 | 12.35 | 4.45 | 36.07 | 9 | 16 |
| Pi | 27.80 | 28.40 | 4.14 | 14.90 | 22 | 32 | 19.75 | 20.05 | 1.96 | 9.96 | 17 | 22 |
| Pi.n | 24.73 | 25.10 | 6.05 | 24.49 | 12 | 38 | 19.04 | 19.70 | 3.82 | 20.06 | 6 | 28 |
| 3–10 total | 26.72 | 27.80 | 6.15 | 23.05 | 9 | 35 | 18.79 | 20.40 | 4.97 | 26.49 | 6 | 25 |
| Pi | 28.67 | 28.45 | 3.59 | 12.55 | 21 | 35 | 20.46 | 20.55 | 2.68 | 13.10 | 13 | 25 |
| Ar.t | 15.67 | 14.90 | 2.94 | 18.76 | 13 | 20 | 8.52 | 8.30 | 1.23 | 14.47 | 7 | 10 |
| 2–1 total | 22.99 | 23.10 | 6.20 | 27.00 | 9 | 35 | 16.23 | 16.70 | 3.87 | 23.90 | 8 | 23 |
| Mj | 16.32 | 17.00 | 5.25 | 32.20 | 9 | 23 | 9.22 | 9.30 | 1.08 | 11.74 | 8 | 11 |
| Pi | 24.41 | 25.45 | 5.02 | 20.59 | 15 | 32 | 17.14 | 17.65 | 3.93 | 22.95 | 8 | 23 |
| Pi.n | 23.81 | 23.10 | 6.03 | 25.34 | 13 | 35 | 17.04 | 16.80 | 2.53 | 14.87 | 12 | 22 |
| 2–9 total | 25.65 | 25.10 | 6.33 | 24.67 | 13 | 41 | 20.06 | 20.90 | 3.23 | 16.12 | 10 | 29 |
| Pi | 31.63 | 31.40 | 1.65 | 5.24 | 30 | 35 | 22.63 | 23.25 | 2.47 | 10.95 | 18 | 25 |
| Pi.n | 25.44 | 25.10 | 6.25 | 24.56 | 13 | 41 | 19.98 | 20.80 | 3.18 | 15.95 | 10 | 29 |
| EP-RP Species | Statistical Parameters | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Basal Diameter (cm) | Total Height (m) | |||||||||||
| Median | s | cv (%) | Min | Max | Median | s | cv (%) | Min | Max | |||
| 1–5 total | 15.19 | 15.10 | 3.57 | 23.55 | 6 | 26 | 13.89 | 14.40 | 2.49 | 17.95 | 4 | 17 |
| Pi | 15.92 | 15.60 | 2.96 | 18.58 | 10 | 23 | 14.84 | 14.95 | 1.73 | 11.69 | 9 | 17 |
| Pi.n | 14.89 | 14.30 | 3.75 | 25.23 | 7 | 26 | 13.49 | 13.90 | 2.40 | 17.85 | 6 | 17 |
| An.a | 14.90 | 15.00 | 0.36 | 2.42 | 15 | 15 | 12.76 | 13.30 | 0.92 | 7.24 | 12 | 13 |
| 1–9 total | 12.64 | 12.70 | 3.21 | 25.37 | 6 | 20 | 10.80 | 10.95 | 2.43 | 22.54 | 5 | 16 |
| Pi | 14.53 | 14.70 | 2.35 | 16.19 | 8 | 20 | 12.81 | 12.85 | 1.63 | 12.72 | 7 | 16 |
| Pi.n | 11.97 | 11.80 | 3.17 | 26.50 | 7 | 19 | 10.04 | 9.85 | 2.23 | 22.28 | 5 | 15 |
| EP-RP Species | Statistical Parameters | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Basal Diameter (cm) | Total Height (m) | |||||||||||
| Median | s | cv (%) | Min | Max | Median | s | cv (%) | Min | Max | |||
| 4–6 total | 22.89 | 23.80 | 8.96 | 39.16 | 9 | 56 | 11.10 | 12.70 | 4.47 | 40.34 | 2 | 18 |
| Pi | 27.21 | 26.40 | 5.29 | 19.44 | 17 | 42 | 22.69 | 22.80 | 2.70 | 11.93 | 15 | 29 |
| Ul | 14.12 | 13.10 | 5.25 | 37.16 | 10 | 39 | 11.95 | 12.10 | 3.36 | 28.18 | 6 | 21 |
| Mj | 12.27 | 10.55 | 3.40 | 27.75 | 9 | 24 | 12.90 | 12.90 | 1.54 | 11.98 | 10 | 15 |
| Pa.m | 29.26 | 32.25 | 17.88 | 61.12 | 12 | 56 | 16.66 | 14.40 | 6.37 | 38.25 | 9 | 25 |
| 4–9 total | 21.88 | 23.67 | 8.93 | 40.82 | 5 | 37 | 17.78 | 19.60 | 5.39 | 30.35 | 4 | 27 |
| Pi.n | 26.27 | 25.42 | 5.17 | 19.69 | 17 | 37 | 20.40 | 20.90 | 2.30 | 11.32 | 15 | 24 |
| Mj | 7.68 | 7.75 | 1.90 | 15.49 | 5 | 10 | 10.85 | 11.00 | 2.59 | 23.90 | 4 | 14 |
| EP3-RP9_Pi.n vs. EP1-RP9_Pi.n | |||||||||
| Vulnerability | Mean—Group 1 | Mean—Group 2 | t-value | df | p | Std.Dev.—Group 1 | Std.Dev.—Group 2 | F-ratio—Variances | p-Variances |
| I < 0.80 | 0.67 | 0.73 | −3.45 | 79 | 0.00 * | 0.09 | 0.05 | 2.51 | 0.00 * |
| 0.80 < I < 1.00 | 0.87 | 0.88 | −0.65 | 60 | 0.51 | 0.05 | 0.05 | 1.00 | 0.95 |
| I > 1.00 | 1.11 | 1.12 | −0.18 | 21 | 0.85 | 0.07 | 0.13 | 3.23 | 0.26 |
| EP3-RP9_Pi.n vs. EP1-RP5_Pi.n | |||||||||
| Vulnerability | Mean—Group 1 | Mean—Group 2 | t-value | df | p | Std.Dev.—Group 1 | Std.Dev.—Group 2 | F-ratio—Variances | p-Variances |
| I < 0.80 | 0.67 | 0.68 | −0.29 | 50 | 0.76 | 0.09 | 0.09 | 1.01 | 0.93 |
| 0.80 < I < 1.00 | 0.87 | 0.88 | −0.65 | 55 | 0.51 | 0.05 | 0.07 | 1.58 | 0.27 |
| I > 1.00 | 1.11 | 1.16 | −0.98 | 27 | 0.33 | 0.07 | 0.11 | 2.33 | 0.42 |
| EP3-RP10_Pi vs. EP1-RP9_Pi | |||||||||
| Vulnerability | Mean—Group 1 | Mean—Group 2 | t-value | df | p | Std.Dev.—Group 1 | Std.Dev.—Group 2 | F-ratio—Variances | p-Variances |
| I < 0.80 | 0.73 | 0.68 | 1.47 | 32 | 0.15 | 0.02 | 0.07 | 6.86 | 0.02 |
| 0.80 < I < 1.00 | 0.89 | 0.87 | 1.10 | 31 | 0.27 | 0.05 | 0.03 | 2.40 | 0.33 |
| EP3-RP10_Pi vs. EP1-RP5_Pi | |||||||||
| Vulnerability | Mean—Group 1 | Mean—Group 2 | t-value | df | p | Std.Dev.—Group 1 | Std.Dev.—Group 2 | F-ratio—Variances | p-Variances |
| I < 0.80 | 0.68 | 0.73 | −1.39 | 32 | 0.17 | 0.07 | 0.06 | 1.36 | 0.74 |
| 0.80 < I < 1.00 | 0.87 | 0.91 | −1.91 | 37 | 0.06 | 0.03 | 0.04 | 1.87 | 0.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cristinel, C.; Ciprian, T.; Popovici, L.; Radu, V.; Crișan, V.; Dincă, L.C. Structural Characteristics of the Pine Stands on Degraded Lands in the South-East of Romania, in the Context of Climate Changes. Appl. Sci. 2024, 14, 8127. https://doi.org/10.3390/app14188127
Cristinel C, Ciprian T, Popovici L, Radu V, Crișan V, Dincă LC. Structural Characteristics of the Pine Stands on Degraded Lands in the South-East of Romania, in the Context of Climate Changes. Applied Sciences. 2024; 14(18):8127. https://doi.org/10.3390/app14188127
Chicago/Turabian StyleCristinel, Constandache, Tudor Ciprian, Laurențiu Popovici, Vlad Radu, Vlad Crișan, and Lucian Constantin Dincă. 2024. "Structural Characteristics of the Pine Stands on Degraded Lands in the South-East of Romania, in the Context of Climate Changes" Applied Sciences 14, no. 18: 8127. https://doi.org/10.3390/app14188127
APA StyleCristinel, C., Ciprian, T., Popovici, L., Radu, V., Crișan, V., & Dincă, L. C. (2024). Structural Characteristics of the Pine Stands on Degraded Lands in the South-East of Romania, in the Context of Climate Changes. Applied Sciences, 14(18), 8127. https://doi.org/10.3390/app14188127