Plant-Derived Flavonoids as AMPK Activators: Unveiling Their Potential in Type 2 Diabetes Management through Mechanistic Insights, Docking Studies, and Pharmacokinetics

Abstract
1. Introduction
2. The Structure and Activation of AMPK
3. The Role of AMPK in T2DM
3.1. The Role of AMPK in Insulin Secretion from Pancreatic Beta Cells
3.2. The Role of AMPK in Skeletal Muscle
4. Plant-Derived Flavonoids That Induce AMPK Activation
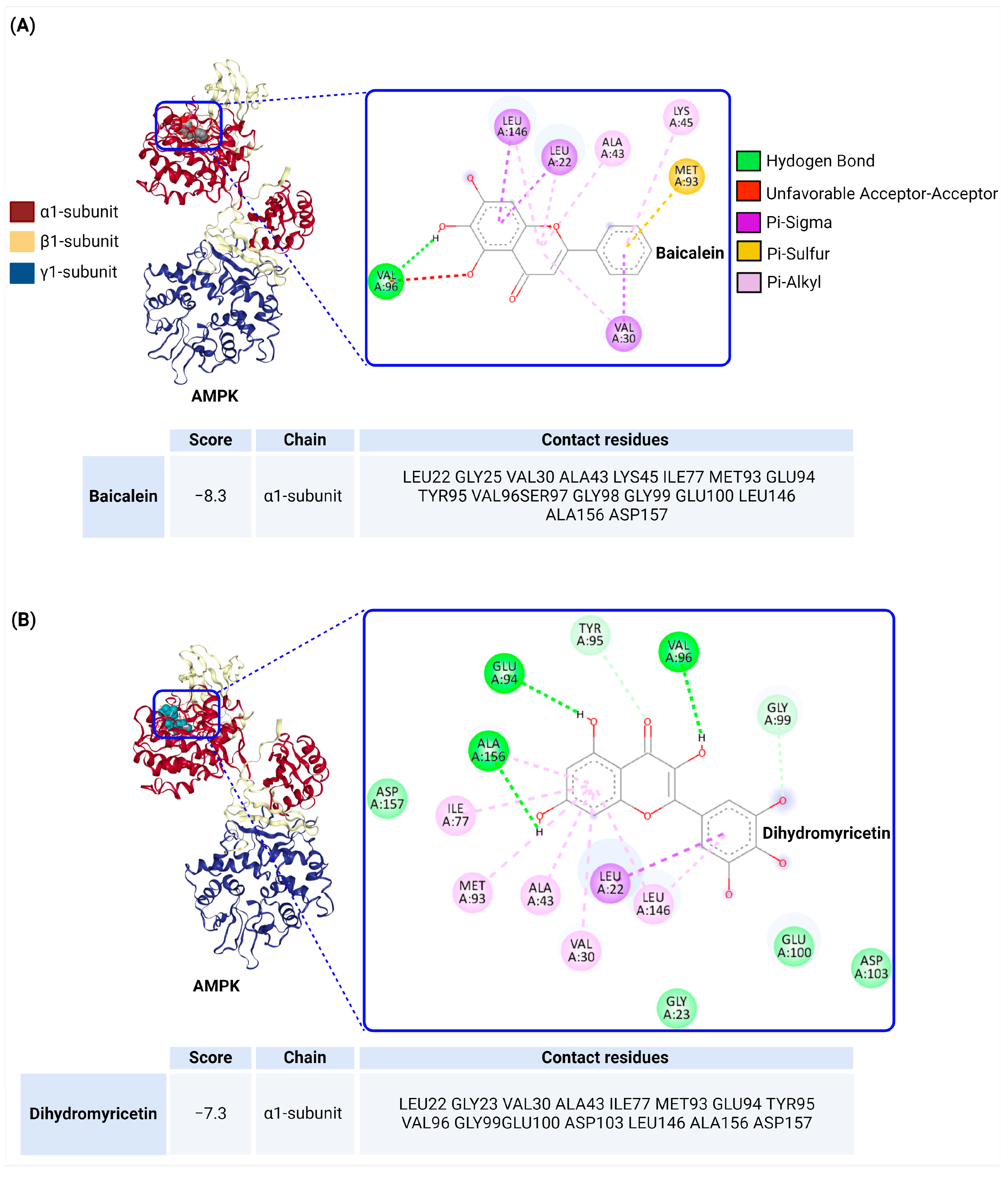
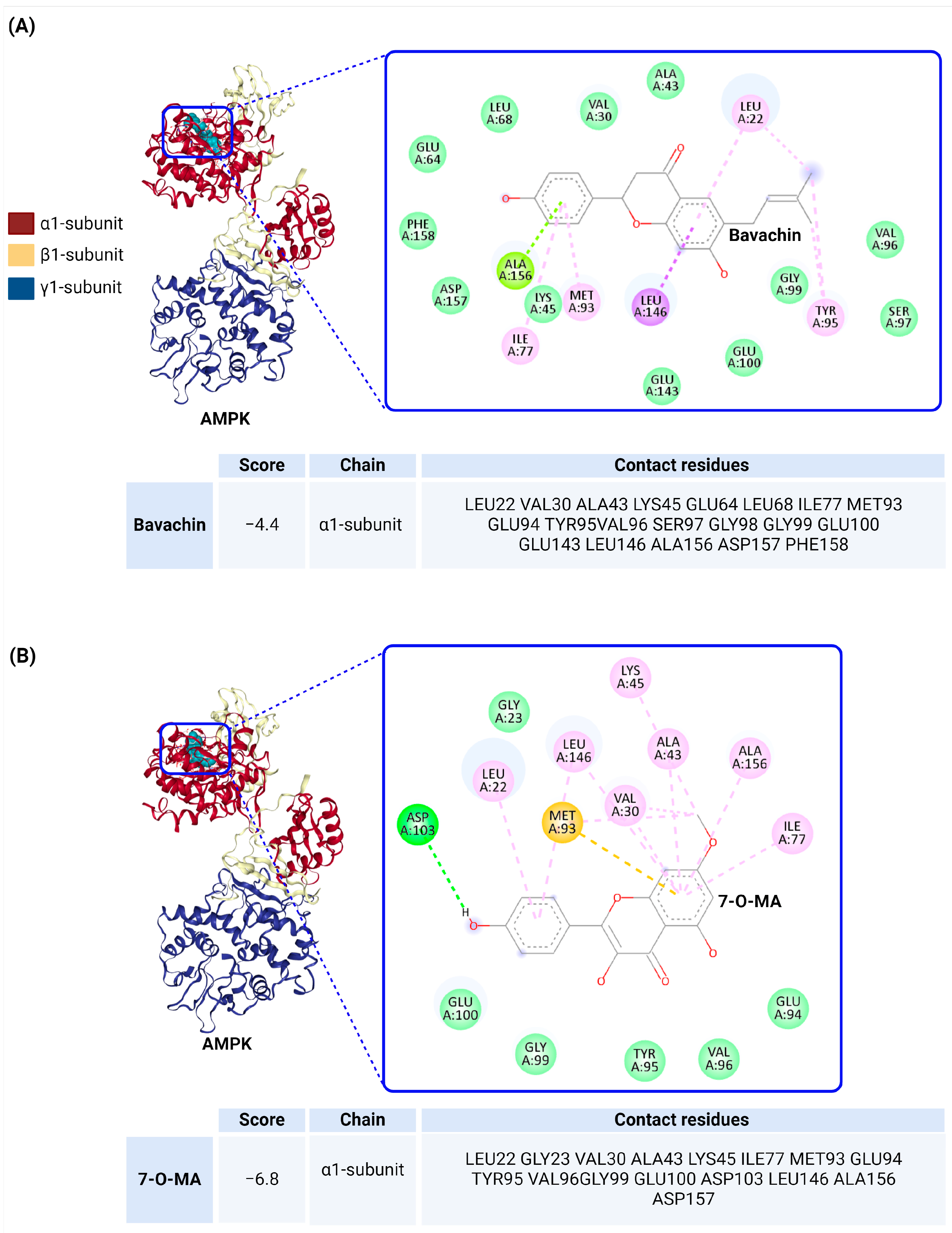
5. Docking Analysis of AMPK with Flavonoids
6. Comprehensive ADME and Drug-Likeness Evaluation of Derrone Using SwissADME
7. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Artasensi, A.; Pedretti, A.; Vistoli, G.; Fumagalli, L. Type 2 Diabetes Mellitus: A Review of Multi-Target Drugs. Molecules 2020, 25, 1987. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Gourgari, E.; Wilhelm, E.E.; Hassanzadeh, H.; Aroda, V.R.; Shoulson, I. A comprehensive review of the FDA-approved labels of diabetes drugs: Indications, safety, and emerging cardiovascular safety data. J. Diabetes Complicat. 2017, 31, 1719–1727. [Google Scholar] [CrossRef] [PubMed]
- Dahlén, A.D.; Dashi, G.; Maslov, I.; Attwood, M.M.; Jonsson, J.; Trukhan, V.; Schiöth, H.B. Trends in Antidiabetic Drug Discovery: FDA Approved Drugs, New Drugs in Clinical Trials and Global Sales. Front. Pharmacol. 2021, 12, 807548. [Google Scholar] [CrossRef]
- Szymczak-Pajor, I.; Wenclewska, S.; Śliwińska, A. Metabolic Action of Metformin. Pharmaceuticals 2022, 15, 810. [Google Scholar] [CrossRef]
- Goel, S.; Singh, R.; Singh, V.; Singh, H.; Kumari, P.; Chopra, H.; Sharma, R.; Nepovimova, E.; Valis, M.; Kuca, K.; et al. Metformin: Activation of 5′ AMP-activated protein kinase and its emerging potential beyond anti-hyperglycemic action. Front. Genet. 2022, 13, 1022739. [Google Scholar] [CrossRef]
- Salminen, A.; Hyttinen, J.M.; Kaarniranta, K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: Impact on healthspan and lifespan. J. Mol. Med. 2011, 89, 667–676. [Google Scholar] [CrossRef]
- Chen, X.; Li, X.; Zhang, W.; He, J.; Xu, B.; Lei, B.; Wang, Z.; Cates, C.; Rousselle, T.; Li, J. Activation of AMPK inhibits inflammatory response during hypoxia and reoxygenation through modulating JNK-mediated NF-κB pathway. Metabolism 2018, 83, 256–270. [Google Scholar] [CrossRef]
- Clark, H.; Carling, D.; Saggerson, D. Covalent activation of heart AMP-activated protein kinase in response to physiological concentrations of long-chain fatty acids. Eur. J. Biochem. 2004, 271, 2215–2224. [Google Scholar] [CrossRef]
- Suchankova, G.; Tekle, M.; Saha, A.K.; Ruderman, N.B.; Clarke, S.D.; Gettys, T.W. Dietary polyunsaturated fatty acids enhance hepatic AMP-activated protein kinase activity in rats. Biochem. Biophys. Res. Commun. 2005, 326, 851–858. [Google Scholar] [CrossRef]
- Kelley, D.E.; Simoneau, J.A. Impaired free fatty acid utilization by skeletal muscle in non-insulin-dependent diabetes mellitus. J. Clin. Investig. 1994, 94, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Dong, M.; Guo, N.; Tian, J.; Lei, P.; Wang, S.; Yang, Y.; Shi, Y. Flavonoids improve type 2 diabetes mellitus and its complications: A review. Front. Nutr. 2023, 10, 1192131. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem Kalhoro, M.; Hussain Kalhoro, D.; Adebowale, T.O.; Usman Mazhar, M.; Rehman, Z.U.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Future J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Francini, F.; Schinella, G.R.; Ríos, J.L. Activation of AMPK by Medicinal Plants and Natural Products: Its Role in Type 2 Diabetes Mellitus. Mini Rev. Med. Chem. 2019, 19, 880–901. [Google Scholar] [CrossRef]
- Thomas, I.; Gregg, B. Metformin; a review of its history and future: From lilac to longevity. Pediatr. Diabetes 2017, 18, 10–16. [Google Scholar] [CrossRef]
- Grace, M.H.; Ribnicky, D.M.; Kuhn, P.; Poulev, A.; Logendra, S.; Yousef, G.G.; Raskin, I.; Lila, M.A. Hypoglycemic activity of a novel anthocyanin-rich formulation from lowbush blueberry, Vaccinium angustifolium Aiton. Phytomedicine 2009, 16, 406–415. [Google Scholar] [CrossRef]
- Babu, P.V.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef]
- Zhang, F.; Feng, J.; Zhang, J.; Kang, X.; Qian, D. Quercetin modulates AMPK/SIRT1/NF-κB signaling to inhibit inflammatory/oxidative stress responses in diabetic high fat diet-induced atherosclerosis in the rat carotid artery. Exp. Ther. Med. 2020, 20, 280. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, W.; Guo, X.; Liu, J.; Li, S.; Zhang, H.; Fan, B. Genistein protects against hyperglycemia and fatty liver disease in diet-induced prediabetes mice via activating hepatic insulin signaling pathway. Front. Nutr. 2022, 9, 1072044. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Ma, J.; Sun, J.; Cheng, C.; Feng, Z.; Jiang, H.; Yang, W. Flavonoid-Rich Extract of Paulownia fortunei Flowers Attenuates Diet-Induced Hyperlipidemia, Hepatic Steatosis and Insulin Resistance in Obesity Mice by AMPK Pathway. Nutrients 2017, 9, 959. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Shaw, R.J. AMPK: Guardian of metabolism and mitochondrial homeostasis. Nat. Rev. Mol. Cell Biol. 2018, 19, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, G.R.; Hardie, D.G. New insights into activation and function of the AMPK. Nat. Rev. Mol. Cell Biol. 2023, 24, 255–272. [Google Scholar] [CrossRef]
- Yan, Y.; Zhou, X.E.; Xu, H.E.; Melcher, K. Structure and Physiological Regulation of AMPK. Int. J. Mol. Sci. 2018, 19, 3534. [Google Scholar] [CrossRef] [PubMed]
- Birk, J.B.; Wojtaszewski, J.F. Predominant alpha2/beta2/gamma3 AMPK activation during exercise in human skeletal muscle. J. Physiol. 2006, 577 Pt 3, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Yang, G.; Kim, Y.; Kim, J.; Ha, J. AMPK activators: Mechanisms of action and physiological activities. Exp. Mol. Med. 2016, 48, e224. [Google Scholar] [CrossRef]
- Xiao, B.; Sanders, M.J.; Underwood, E.; Heath, R.; Mayer, F.V.; Carmena, D.; Jing, C.; Walker, P.A.; Eccleston, J.F.; Haire, L.F.; et al. Structure of mammalian AMPK and its regulation by ADP. Nature 2011, 472, 230–233. [Google Scholar] [CrossRef]
- Gowans, G.J.; Hawley, S.A.; Ross, F.A.; Hardie, D.G. AMP is a true physiological regulator of AMP-activated protein kinase by both allosteric activation and enhancing net phosphorylation. Cell Metab. 2013, 18, 556–566. [Google Scholar] [CrossRef]
- Woods, A.; Dickerson, K.; Heath, R.; Hong, S.P.; Momcilovic, M.; Johnstone, S.R.; Carlson, M.; Carling, D. Ca2+/calmodulin-dependent protein kinase kinase-beta acts upstream of AMP-activated protein kinase in mammalian cells. Cell Metab. 2005, 2, 21–33. [Google Scholar] [CrossRef]
- Ciccarese, F.; Zulato, E.; Indraccolo, S. LKB1/AMPK Pathway and Drug Response in Cancer: A Therapeutic Perspective. Oxid. Med. Cell. Longev. 2019, 2019, 8730816. [Google Scholar] [CrossRef] [PubMed]
- Hawley, S.A.; Selbert, M.A.; Goldstein, E.G.; Edelman, A.M.; Carling, D.; Hardie, D.G. 5′-AMP activates the AMP-activated protein kinase cascade, and Ca2+/calmodulin activates the calmodulin-dependent protein kinase I cascade, via three independent mechanisms. J. Biol. Chem. 1995, 270, 27186–27191. [Google Scholar] [CrossRef]
- Davies, S.P.; Helps, N.R.; Cohen, P.T.; Hardie, D.G. 5′-AMP inhibits dephosphorylation, as well as promoting phosphorylation, of the AMP-activated protein kinase. Studies using bacterially expressed human protein phosphatase-2C alpha and native bovine protein phosphatase-2AC. FEBS Lett. 1995, 377, 421–425. [Google Scholar] [PubMed]
- Oakhill, J.S.; Steel, R.; Chen, Z.P.; Scott, J.W.; Ling, N.; Tam, S.; Kemp, B.E. AMPK is a direct adenylate charge-regulated protein kinase. Science 2011, 332, 1433–1435. [Google Scholar] [CrossRef]
- Leclerc, I.; Woltersdorf, W.W.; da Silva Xavier, G.; Rowe, R.L.; Cross, S.E.; Korbutt, G.S.; Rajotte, R.V.; Smith, R.; Rutter, G.A. Metformin, but not leptin, regulates AMP-activated protein kinase in pancreatic islets: Impact on glucose-stimulated insulin secretion. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E1023–E1031. [Google Scholar] [CrossRef]
- Sun, Y.; Ren, M.; Gao, G.Q.; Gong, B.; Xin, W.; Guo, H.; Zhang, X.J.; Gao, L.; Zhao, J.J. Chronic palmitate exposure inhibits AMPKalpha and decreases glucose-stimulated insulin secretion from beta-cells: Modulation by fenofibrate. Acta Pharmacol. Sin. 2008, 29, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Henquin, J.C. The dual control of insulin secretion by glucose involves triggering and amplifying pathways in β-cells. Diabetes Res. Clin. Pract. 2011, 93 (Suppl. 1), S27–S31. [Google Scholar] [CrossRef]
- Prentki, M.; Matschinsky, F.M.; Madiraju, S.R. Metabolic signaling in fuel-induced insulin secretion. Cell Metab. 2013, 18, 162–185. [Google Scholar] [CrossRef] [PubMed]
- Akkan, A.G.; Malaisse, W.J. Insulinotropic action of AICA riboside. I. Insulin release by isolated islets and the perfused pancreas. Diabetes Res. 1994, 25, 13–23. [Google Scholar]
- Malaisse, W.J.; Conget, I.; Sener, A.; Rorsman, P. Insulinotropic action of AICA riboside. II. Secretory, metabolic and cationic aspects. Diabetes Res. 1994, 25, 25–37. [Google Scholar]
- Pepin, É.; Al-Mass, A.; Attané, C.; Zhang, K.; Lamontagne, J.; Lussier, R.; Madiraju, S.R.; Joly, E.; Ruderman, N.B.; Sladek, R.; et al. Pancreatic β-Cell Dysfunction in Diet-Induced Obese Mice: Roles of AMP-Kinase, Protein Kinase Cε, Mitochondrial and Cholesterol Metabolism, and Alterations in Gene Expression. PLoS ONE 2016, 11, e0153017. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhou, L.; Li, G.; Luo, T.; Gu, Y.; Qian, L.; Fu, X.; Li, F.; Li, J.; Luo, M. Palmitate activates AMP-activated protein kinase and regulates insulin secretion from beta cells. Biochem. Biophys. Res. Commun. 2007, 352, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; You, Y.H.; Ham, D.S.; Yang, H.K.; Yoon, K.H. The Paradoxical Effects of AMPK on Insulin Gene Expression and Glucose-Induced Insulin Secretion. J. Cell Biochem. 2016, 117, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Prentki, M.; Joly, E.; El-Assaad, W.; Roduit, R. Malonyl-CoA signaling, lipid partitioning, and glucolipotoxicity: Role in beta-cell adaptation and failure in the etiology of diabetes. Diabetes 2002, 51 (Suppl. 3), S405–S413. [Google Scholar] [CrossRef] [PubMed]
- Herman, R.; Kravos, N.A.; Jensterle, M.; Janež, A.; Dolžan, V. Metformin and Insulin Resistance: A Review of the Underlying Mechanisms behind Changes in GLUT4-Mediated Glucose Transport. Int. J. Mol. Sci. 2022, 23, 1264. [Google Scholar] [CrossRef]
- Shamshoum, H.; Vlavcheski, F.; MacPherson, R.E.K.; Tsiani, E. Rosemary extract activates AMPK, inhibits mTOR and attenuates the high glucose and high insulin-induced muscle cell insulin resistance. Appl. Physiol. Nutr. Metab. 2021, 46, 819–827. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, Z.; Zhai, W.; Bi, Y.; Yin, Y.; Zhang, W. Expression and purification of asprosin in Pichia pastoris and investigation of its increase glucose uptake activity in skeletal muscle through activation of AMPK. Enzyme Microb. Technol. 2021, 144, 109737. [Google Scholar] [CrossRef]
- Prabha, B.; Sini, S.; Sherin, D.R.; Neethu, S.; Rameshkumar, K.B.; Manojkumar, T.K.; Jayamurthy, P.; Radhakrishnan, K.V. Promalabaricone B from Myristica fatua Houtt. seeds demonstrate antidiabetic potential by modulating glucose uptake via the upregulation of AMPK in L6 myotubes. Nat. Prod. Res. 2021, 35, 867–872. [Google Scholar] [CrossRef]
- Natsume, N.; Yonezawa, T.; Saito, Y.; Woo, J.T.; Teruya, T. Prenylflavonoids from fruit of Macaranga tanarius promote glucose uptake via AMPK activation in L6 myotubes. J. Nat. Med. 2021, 75, 813–823. [Google Scholar] [CrossRef]
- Lee, H.A.; Cho, J.H.; Afinanisa, Q.; An, G.H.; Han, J.G.; Kang, H.J.; Choi, S.H.; Seong, H.A. Ganoderma lucidum Extract Reduces Insulin Resistance by Enhancing AMPK Activation in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 3338. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Beecher, G.R. Overview of dietary flavonoids: Nomenclature, occurrence and intake. J. Nutr. 2003, 133, 3248s–3254s. [Google Scholar] [CrossRef] [PubMed]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, flavones, flavanones, and human health: Epidemiological evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef]
- Arts, I.C.; Hollman, P.C. Polyphenols and disease risk in epidemiologic studies. Am. J. Clin. Nutr. 2005, 81 (Suppl. 1), 317s–325s. [Google Scholar] [CrossRef]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, T.; Zhou, Y.; Zeng, X.; Ran, L.; Zhang, Q.; Zhu, J.; Mi, M. Dihydromyricetin improves skeletal muscle insulin sensitivity by inducing autophagy via the AMPK-PGC-1α-Sirt3 signaling pathway. Endocrine 2015, 50, 378–389. [Google Scholar] [CrossRef]
- Lee, H.; Li, H.; Noh, M.; Ryu, J.H. Bavachin from Psoralea corylifolia Improves Insulin-Dependent Glucose Uptake through Insulin Signaling and AMPK Activation in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2016, 17, 527. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Lee, J.J.; Kim, I.S.; Kim, Y.; Park, J.S.; Myung, C.S. 7-O-methylaromadendrin stimulates glucose uptake and improves insulin resistance in vitro. Biol. Pharm. Bull. 2010, 33, 1494–1499. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, F.; Wang, Y.; Du, Z.; Liu, D.; Guo, H.; Shen, J.; Peng, H. CaMKKβ is involved in AMP-activated protein kinase activation by baicalin in LKB1 deficient cell lines. PLoS ONE 2012, 7, e47900. [Google Scholar] [CrossRef]
- Yang, L.L.; Xiao, N.; Liu, J.; Liu, K.; Liu, B.; Li, P.; Qi, L.W. Differential regulation of baicalin and scutellarin on AMPK and Akt in promoting adipose cell glucose disposal. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 598–606. [Google Scholar] [CrossRef]
- Pu, P.; Wang, X.A.; Salim, M.; Zhu, L.H.; Wang, L.; Chen, K.J.; Xiao, J.F.; Deng, W.; Shi, H.W.; Jiang, H.; et al. Baicalein, a natural product, selectively activating AMPKα(2) and ameliorates metabolic disorder in diet-induced mice. Mol. Cell Endocrinol. 2012, 362, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Dhanya, R.; Arya, A.D.; Nisha, P.; Jayamurthy, P. Quercetin, a Lead Compound against Type 2 Diabetes Ameliorates Glucose Uptake via AMPK Pathway in Skeletal Muscle Cell Line. Front. Pharmacol. 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.T.; Park, I.J.; Shin, J.I.; Lee, Y.K.; Lee, S.K.; Baik, H.W.; Ha, J.; Park, O.J. Genistein, EGCG, and capsaicin inhibit adipocyte differentiation process via activating AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2005, 338, 694–699. [Google Scholar] [CrossRef]
- Lee, M.S.; Kim, C.H.; Hoang, D.M.; Kim, B.Y.; Sohn, C.B.; Kim, M.R.; Ahn, J.S. Genistein-derivatives from Tetracera scandens stimulate glucose-uptake in L6 myotubes. Biol. Pharm. Bull. 2009, 32, 504–508. [Google Scholar] [CrossRef]
- Son, M.J.; Minakawa, M.; Miura, Y.; Yagasaki, K. Aspalathin improves hyperglycemia and glucose intolerance in obese diabetic ob/ob mice. Eur. J. Nutr. 2013, 52, 1607–1619. [Google Scholar] [CrossRef]
- Owen, M.R.; Doran, E.; Halestrap, A.P. Evidence that metformin exerts its anti-diabetic effects through inhibition of complex 1 of the mitochondrial respiratory chain. Biochem. J. 2000, 348 Pt 3, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Fryer, L.G.; Parbu-Patel, A.; Carling, D. The Anti-diabetic drugs rosiglitazone and metformin stimulate AMP-activated protein kinase through distinct signaling pathways. J. Biol. Chem. 2002, 277, 25226–25232. [Google Scholar] [CrossRef]
- Corton, J.M.; Gillespie, J.G.; Hawley, S.A.; Hardie, D.G. 5-aminoimidazole-4-carboxamide ribonucleoside. A specific method for activating AMP-activated protein kinase in intact cells? Eur. J. Biochem. 1995, 229, 558–565. [Google Scholar] [CrossRef]
- Sullivan, J.E.; Brocklehurst, K.J.; Marley, A.E.; Carey, F.; Carling, D.; Beri, R.K. Inhibition of lipolysis and lipogenesis in isolated rat adipocytes with AICAR, a cell-permeable activator of AMP-activated protein kinase. FEBS Lett. 1994, 353, 33–36. [Google Scholar] [CrossRef]
- Scott, J.W.; van Denderen, B.J.; Jorgensen, S.B.; Honeyman, J.E.; Steinberg, G.R.; Oakhill, J.S.; Iseli, T.J.; Koay, A.; Gooley, P.R.; Stapleton, D.; et al. Thienopyridone drugs are selective activators of AMP-activated protein kinase beta1-containing complexes. Chem. Biol. 2008, 15, 1220–1230. [Google Scholar] [CrossRef]
- Moreno, D.; Knecht, E.; Viollet, B.; Sanz, P. A769662, a novel activator of AMP-activated protein kinase, inhibits non-proteolytic components of the 26S proteasome by an AMPK-independent mechanism. FEBS Lett. 2008, 582, 2650–2654. [Google Scholar] [CrossRef] [PubMed]
- Hawley, S.A.; Fullerton, M.D.; Ross, F.A.; Schertzer, J.D.; Chevtzoff, C.; Walker, K.J.; Peggie, M.W.; Zibrova, D.; Green, K.A.; Mustard, K.J.; et al. The ancient drug salicylate directly activates AMP-activated protein kinase. Science 2012, 336, 918–922. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.W.; Foretz, M.; Bultot, L.; Fullerton, M.D.; Deak, M.; Ross, F.A.; Hawley, S.A.; Shpiro, N.; Viollet, B.; Barron, D.; et al. Mechanism of action of compound-13: An α1-selective small molecule activator of AMPK. Chem. Biol. 2014, 21, 866–879. [Google Scholar] [CrossRef]
- Xiao, B.; Sanders, M.J.; Carmena, D.; Bright, N.J.; Haire, L.F.; Underwood, E.; Patel, B.R.; Heath, R.B.; Walker, P.A.; Hallen, S.; et al. Structural basis of AMPK regulation by small molecule activators. Nat. Commun. 2013, 4, 3017. [Google Scholar] [CrossRef]
- Arunkumar, E.; Anuradha, C.V. Genistein promotes insulin action through adenosine monophosphate-activated protein kinase activation and p70 ribosomal protein S6 kinase 1 inhibition in the skeletal muscle of mice fed a high energy diet. Nutr. Res. 2012, 32, 617–625. [Google Scholar] [CrossRef]
- Akinfenwa, A.O.; Abdul, N.S.; Marnewick, J.L.; Hussein, A.A. Protective Effects of Linearthin and Other Chalcone Derivatives from Aspalathus linearis (Rooibos) against UVB Induced Oxidative Stress and Toxicity in Human Skin Cells. Plants 2021, 10, 1936. [Google Scholar] [CrossRef]
- Thilakarathna, S.H.; Rupasinghe, H.P. Flavonoid bioavailability and attempts for bioavailability enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef] [PubMed]
- Křížková, J.; Burdová, K.; Stiborová, M.; Křen, V.; Hodek, P. The effects of selected flavonoids on cytochromes P450 in rat liver and small intestine. Interdiscip. Toxicol. 2009, 2, 201–204. [Google Scholar] [CrossRef]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. Int. J. Pharm. 2019, 570, 118642. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, J.; Xu, Y.; Zhou, N.; Peng, J.; Xiong, Z.; Liu, X.; Luo, X.; Luo, C.; Chen, K.; et al. In silico ADME/T modelling for rational drug design. Q. Rev. Biophys. 2015, 48, 488–515. [Google Scholar] [CrossRef]
- Grossman, I. ADME pharmacogenetics: Current practices and future outlook. Expert Opin. Drug Metab. Toxicol. 2009, 5, 449–462. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, S.; Mishra, S.; Ojha, S.; Upadhyay, P. ADME Prediction, Structure-activity Relationship of Boswellic Acid Scaffold for the Aspect of Anticancer & Anti-inflammatory Potency. Anticancer. Agents Med. Chem. 2023, 23, 1499–1505. [Google Scholar] [PubMed]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef]
- Kabała-Dzik, A.; Rzepecka-Stojko, A.; Kubina, R.; Iriti, M.; Wojtyczka, R.D.; Buszman, E.; Stojko, J. Flavonoids, bioactive components of propolis, exhibit cytotoxic activity and induce cell cycle arrest and apoptosis in human breast cancer cells MDA-MB-231 and MCF-7—A comparative study. Cell Mol. Biol. 2018, 64, 1–10. [Google Scholar] [CrossRef]
- Murata, M.; Midorikawa, K.; Koh, M.; Umezawa, K.; Kawanishi, S. Genistein and daidzein induce cell proliferation and their metabolites cause oxidative DNA damage in relation to isoflavone-induced cancer of estrogen-sensitive organs. Biochemistry 2004, 43, 2569–2577. [Google Scholar] [CrossRef] [PubMed]
- James, K.D.; Forester, S.C.; Lambert, J.D. Dietary pretreatment with green tea polyphenol, (-)-epigallocatechin-3-gallate reduces the bioavailability and hepatotoxicity of subsequent oral bolus doses of (-)-epigallocatechin-3-gallate. Food Chem. Toxicol. 2015, 76, 103–108. [Google Scholar]
- Lambert, J.D.; Kennett, M.J.; Sang, S.; Reuhl, K.R.; Ju, J.; Yang, C.S. Hepatotoxicity of high oral dose (-)-epigallocatechin-3-gallate in mice. Food Chem. Toxicol. 2010, 48, 409–416. [Google Scholar] [CrossRef]
- Matthies, A.; Loh, G.; Blaut, M.; Braune, A. Daidzein and genistein are converted to equol and 5-hydroxy-equol by human intestinal Slackia isoflavoniconvertens in gnotobiotic rats. J. Nutr. 2012, 142, 40–46. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Treatment Concentration | Model (Cell or Animal) | Change in ADP/AMP Ratio | Results | Ref. |
|---|---|---|---|---|---|
| Dihydromyricetin | 50 mg/kg/day | In vivo (skeletal muscle of mice) | N.T | Increased insulin sensitivity, glucose uptake, and autophagy in skeletal muscle via the activation of the AMPK-PGC-1α-Sirt3 signaling pathway. | [56] |
| Bavachin | Up to 10 µM | 3T3-L1 adipocytes | N.T | Increased glucose uptake via GLUT4 translocation, activated the Akt and AMPK pathways, and enhanced insulin signaling. | [57] |
| 7-O-MA | Up to 10 µM | HepG2 cells, 3T3-L1 adipocytes | N.T | Enhanced glucose uptake, activated the AMPK and PI3K pathways, and improved insulin resistance. | [58] |
| Baicalin | Up to 5 µM | HeLa cells, A549 cells, HepG2 cells | No | Activated AMPK via a CaMKKβ-dependent pathway, increased intracellular calcium, and reduced lipid accumulation. | [59] |
| Baicalein | 400 mg/kg/day | High-fat-diet-induced mice, primary hepatocytes | N.T | Improved metabolic syndrome via the activation of AMPKα2, inhibited inflammation and insulin resistance, and promoted lipid oxidation through various pathways. | [61] |
| Quercetin | Up to 100 µM | L6 myotubes (skeletal muscle cells) | Yes (Increase) [49] | Enhanced glucose uptake via the AMPK pathway and p38 MAPK, independent of insulin signaling; involved changes in mitochondrial potential and intracellular calcium. | [62] |
| Genistein | 20–200 μM | 3T3-L1 adipocytes (cells) | Yes (Increase) [53] | Inhibited adipocyte differentiation and induced apoptosis in mature adipocytes via the rapid activation of AMPK. | [63] |
| Derrone | Up to 25 µM | L6 myotubes (cells) | N.T | Enhanced glucose uptake, activated AMPK, increased GLUT4 and GLUT1 expression, and inhibited PTP1B activity without muscle cell toxicity. | [64] |
| Alpinumisoflavone | Up to 25 µM | L6 myotubes (cells) | N.T | Enhanced glucose uptake, activated AMPK, upregulated GLUT4 and GLUT1 expression, and inhibited PTP1B activity. | [64] |
| Aspalathin | 0.1% (in diet) | ob/ob mice, L6 myotubes | Yes (Increase) [54] | Increased glucose uptake, promoted AMPK phosphorylation, enhanced GLUT4 translocation, decreased fasting blood glucose levels, and improved glucose intolerance. | [65] |
| Description | Value/Range (If Applicable) | Baicalein | Dihydromyricetin | |
|---|---|---|---|---|
| Formula | C15H10O5 | C15H12O8 | ||
| Molecular weight | Helps determine drug absorption and bioavailability. | Typically <500 g/mol (Lipinski rule) | 270.24 g/mol | 320.25 g/mol |
| Num. H-bond acceptors | Important for solubility and interaction with biological molecules. | 0–10 (Lipinski rule) | 5 | 8 |
| Num. H-bond donors | Important for solubility and interaction with biological molecules. | 0–5 (Lipinski rule) | 3 | 6 |
| TPSA | Influences drug absorption, permeability, and ability to cross biological membranes. | <140 Å2 (Lipinski rule) | 90.90 Å2 | 147.68 Å2 |
| Consensus Log Po/w | Reflects the balance between water and fat solubility, crucial for absorption and distribution. | Optimal range: 0–5 (Lipinski rule) | 2.24 | 0.17 |
| GI absorption | Determines how well the compound is absorbed in the gastrointestinal tract. | High, Medium, or Low (qualitative) | High | Low |
| BBB-permeant | Critical for evaluating whether the compound can cross the blood–brain barrier. | Yes/No (qualitative) | No | No |
| CYP3A4 inhibitor | Key enzyme involved in drug metabolism, which can affect drug–drug interactions. | Yes/No (qualitative) | Yes | No |
| Lipinski | Widely used rule of thumb for predicting oral bioavailability. | Follows four of Lipinski’s rules: MW < 500, H-bond acceptors < 10, H-bond donors < 5, Log Po/w < 5 | Yes; 0 violation | Yes; 1 violation: NHorOH > 5 |
| Bioavailability Score | Assesses the compound’s potential to be absorbed and used by the body. | 0.55–1 (high bioavailability) | 0.55 | 0.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, D.O. Plant-Derived Flavonoids as AMPK Activators: Unveiling Their Potential in Type 2 Diabetes Management through Mechanistic Insights, Docking Studies, and Pharmacokinetics. Appl. Sci. 2024, 14, 8607. https://doi.org/10.3390/app14198607
Moon DO. Plant-Derived Flavonoids as AMPK Activators: Unveiling Their Potential in Type 2 Diabetes Management through Mechanistic Insights, Docking Studies, and Pharmacokinetics. Applied Sciences. 2024; 14(19):8607. https://doi.org/10.3390/app14198607
Chicago/Turabian StyleMoon, Dong Oh. 2024. "Plant-Derived Flavonoids as AMPK Activators: Unveiling Their Potential in Type 2 Diabetes Management through Mechanistic Insights, Docking Studies, and Pharmacokinetics" Applied Sciences 14, no. 19: 8607. https://doi.org/10.3390/app14198607
APA StyleMoon, D. O. (2024). Plant-Derived Flavonoids as AMPK Activators: Unveiling Their Potential in Type 2 Diabetes Management through Mechanistic Insights, Docking Studies, and Pharmacokinetics. Applied Sciences, 14(19), 8607. https://doi.org/10.3390/app14198607