


Phytoplankton as CO2 Sinks: Redirecting the Carbon Cycle




Abstract
:1. Introduction
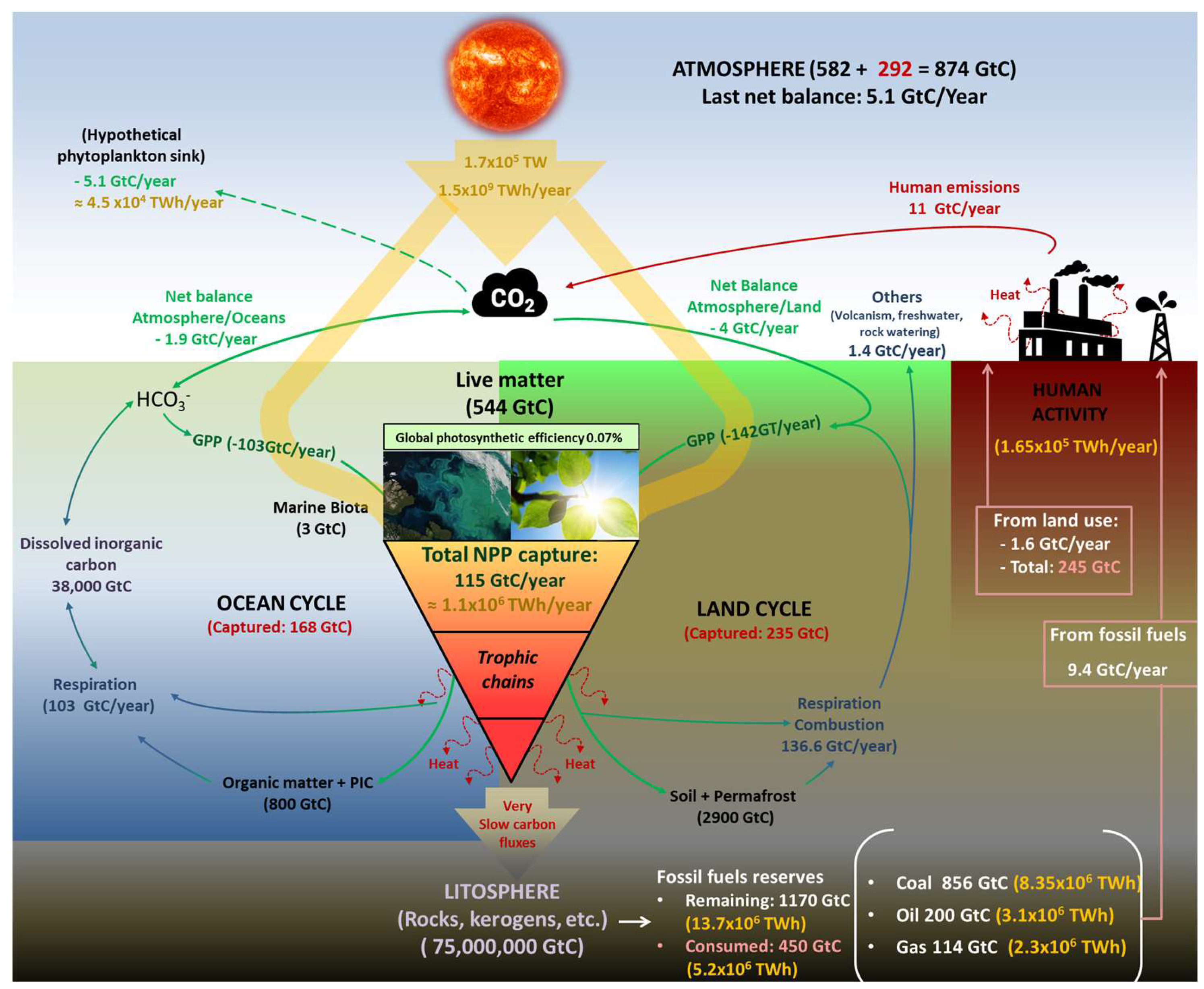
2. Photosynthesis and Global Balances of Matter and Energy
2.1. Energy Inputs in Perspective
2.2. Carbon Cycle and Civilization
2.3. Photosynthetic Efficiency
3. Phytoplankton Culture
3.1. A Brief History
3.2. Photobioreactors (PBRs)
3.2.1. Open Ponds/Artificial Lakes
3.2.2. Closed PBRs
3.2.3. Indoor/Outdoor PBRs
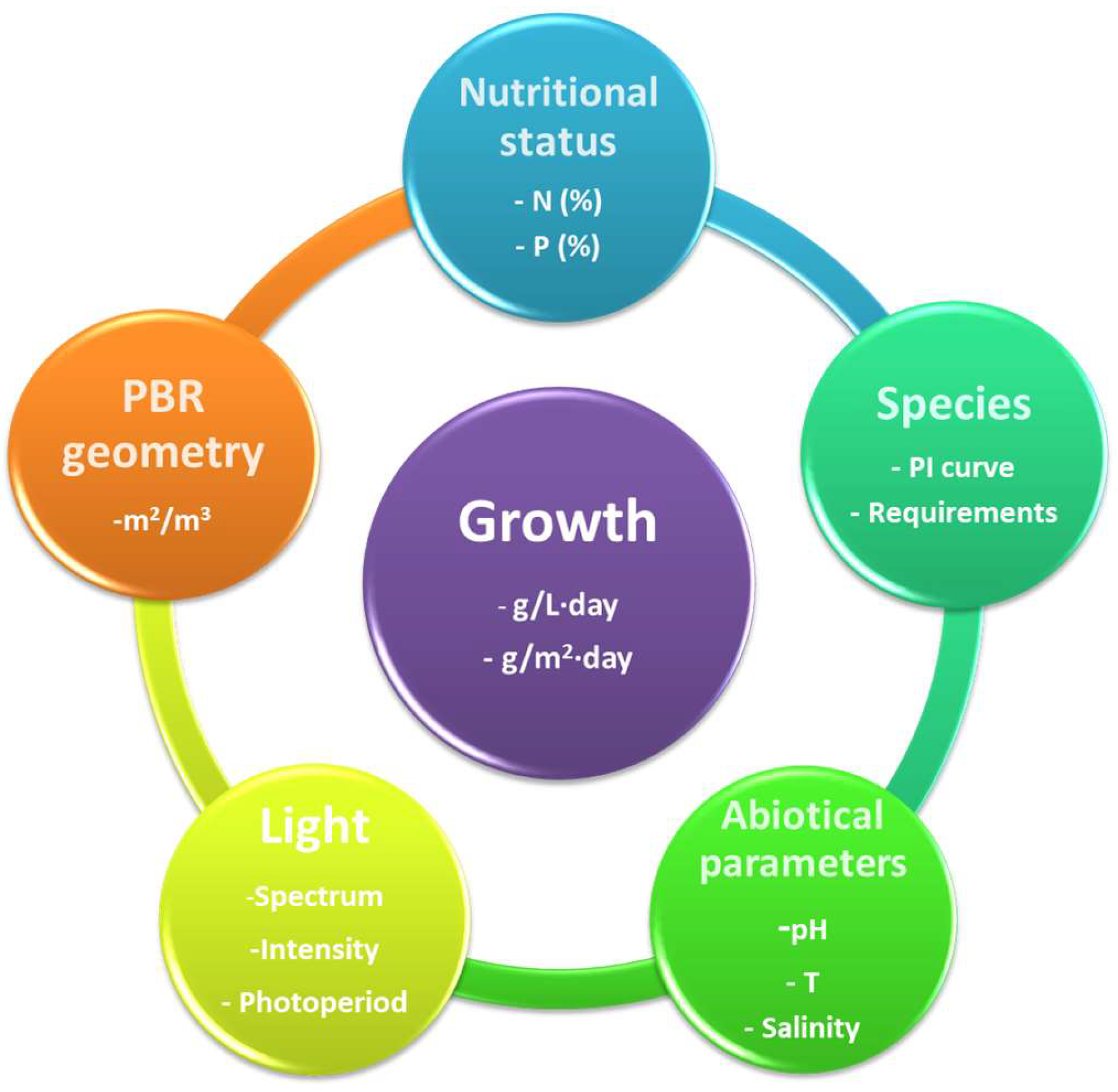
3.3. Key Parameters for the Growth
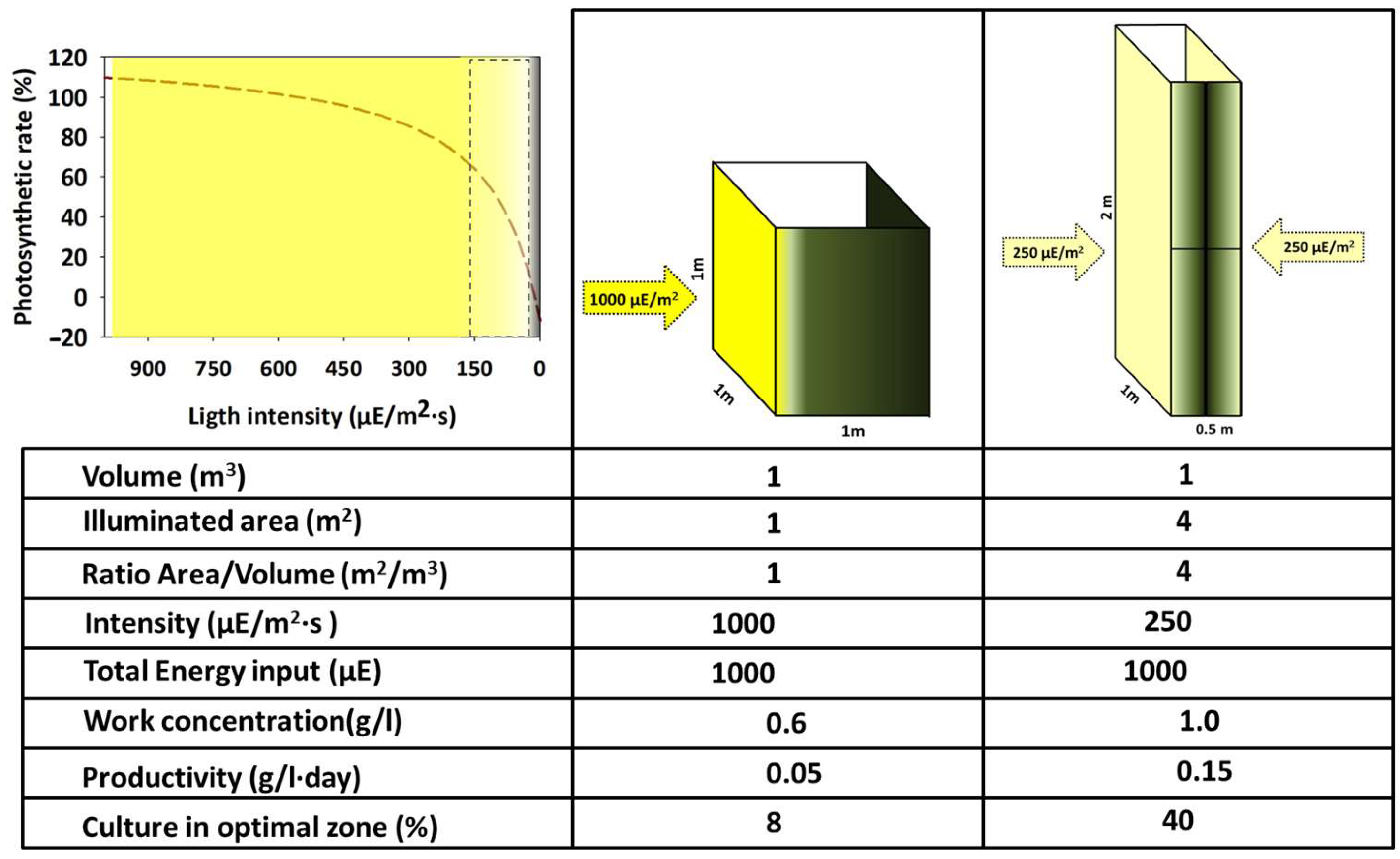
3.3.1. Light
3.3.2. Other Abiotic Parameters
- (a)
- Temperature
- (b)
- Medium pH
- -
- In the absence of any gas exchange, protons are gradually consumed as they are incorporated into new organic compounds through the Calvin cycle, increasing pH. This increase can also cause the precipitation of carbonates, negatively impacting photosynthesis since e pH values are outside the optimal range and a lack of precursors accumulates.
- -
- When the culture medium is aerated with air (0.04% CO2), the pH value will reach an equilibrium that mainly depends on the water alkalinity. The maximum pH value under these conditions is usually approximately 8.3, beyond which carbonate starts to precipitate. When the photosynthetic demand for CO2 is less than the atmospheric CO2 solubilisation rate, the pH stays stable and functions as a buffer system. However, if the CO2 demand exceeds the atmospheric delivery rate due to the increment in cellular concentration and/or light intensity, the initial scenario of proton depletion will be gradually reached.
- -
- In some culture systems, the medium is aerated with CO2-enriched air. The final pH value, after reaching equilibrium, decreases linearly concerning the concentration of this gas in the air. At a CO2 concentration between 0.5% and 2.0%, the pH remains strongly buffered between 7.0 and 8.5 (depending on the alkalinity of the medium), which is suitable for the growth of most species. At the photosynthetic level, this strategy ensures an ample supply of CO2, even in high cell concentrations or light intensities. Hence, CO2 is never depleted, so the pH remains constantly buffered, and the photosynthesis rate is never limited.
- (c)
- Major ions and trace metals:
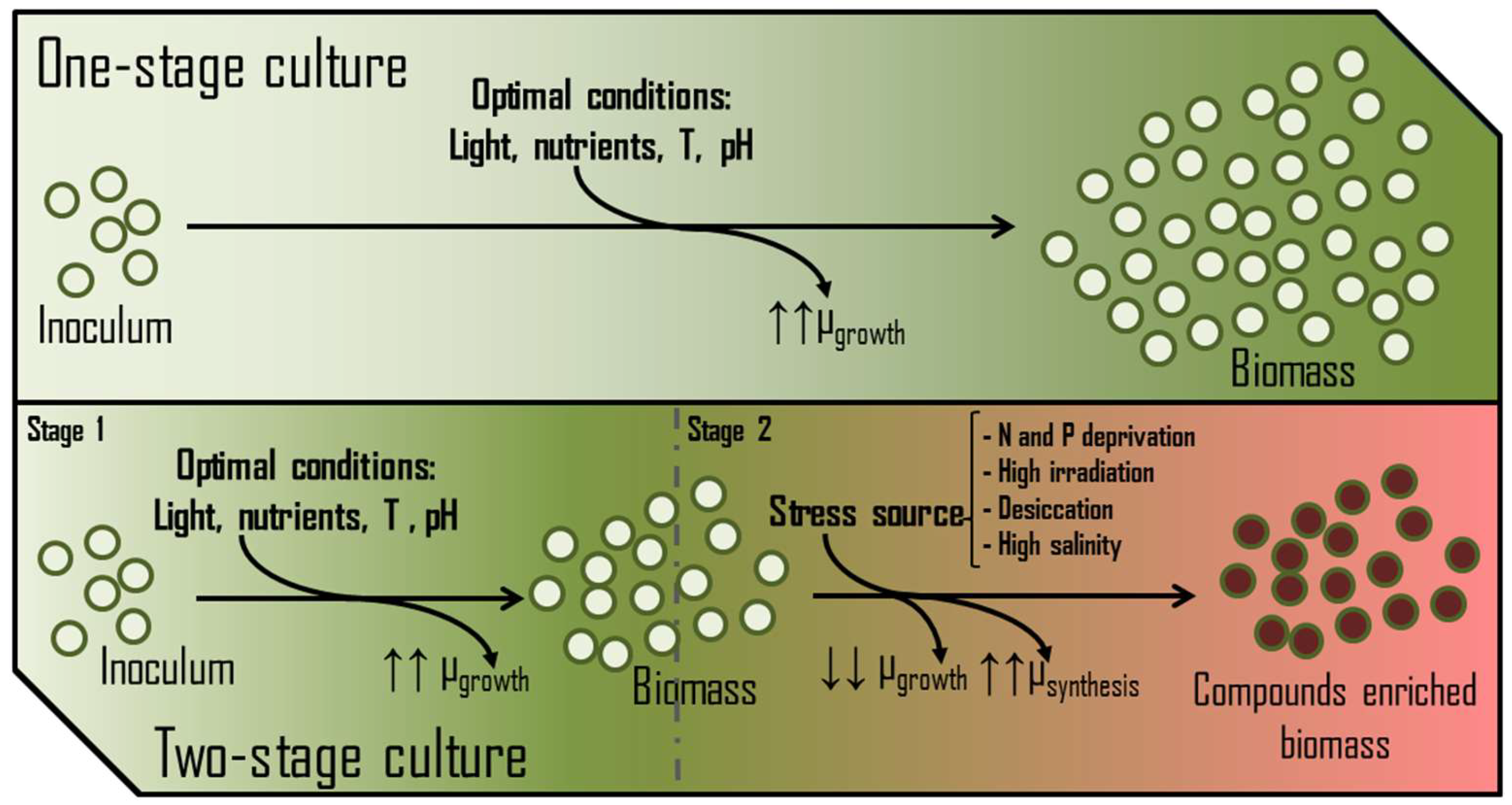
3.3.3. Nutritional Status
3.3.4. Culture Mixing
3.4. Estimations of Growth
3.5. Potential Numbers of Production
4. Mitigating Fossil Energy Dependence by Enhancing Photosynthesis
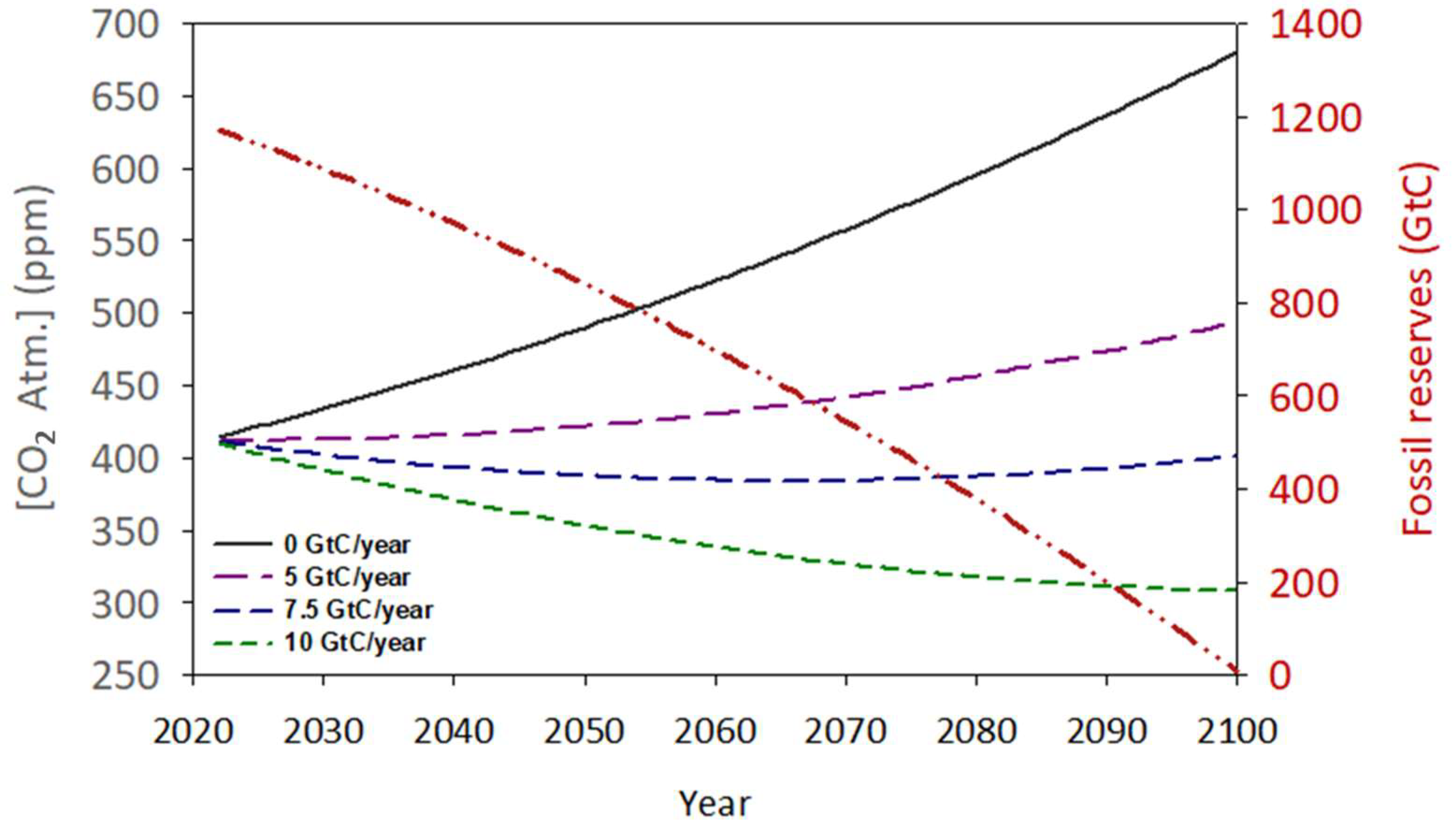
4.1. Fossil Fuel Reserves and Potential Evolution
4.2. Biological Capture of CO2
4.3. A Matter about Figures and Politics
4.4. Technical Challenges
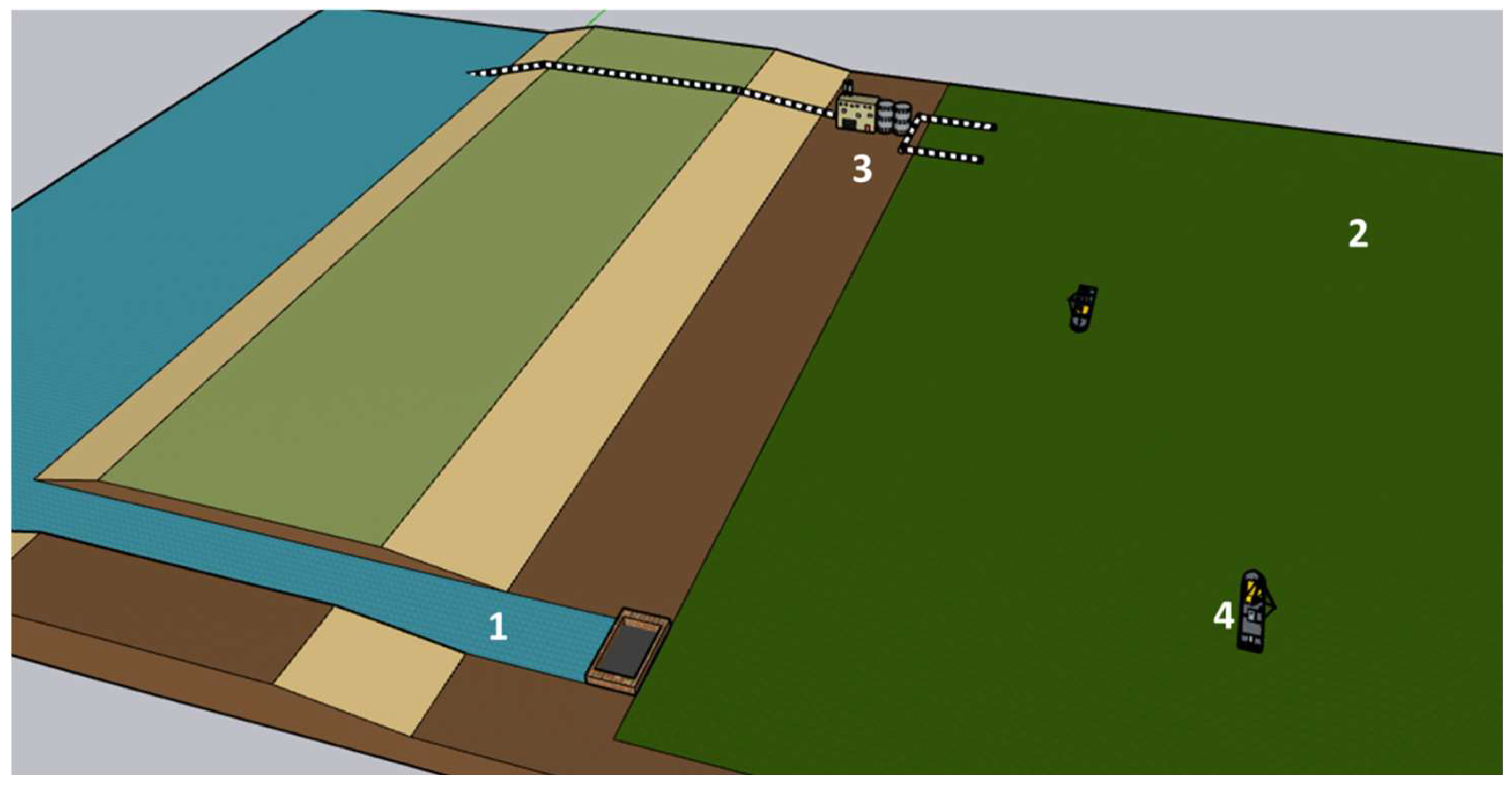
4.4.1. PBR Features
4.4.2. Water Use
4.4.3. Species and Growth Medium
4.4.4. Mixing System
4.4.5. Temperature Control
4.4.6. Sources of Nutrients
4.4.7. Biomass Management
- (a)
- Carbon lock: This option involves closing the biomass by burying it or sealing it in airtight bunkers to prevent its spontaneous decomposition from releasing CO2 and CH4 into the atmosphere [101]. This way, the captured carbon is permanently removed from the carbon cycle and detoxifies the atmosphere. However, the economic viability of this option is limited as it does not generate added value to the final product of the technology and becomes waste instead.
- (b)
- Energy source: As discussed, 13 Gt of biomass per year can store approximately 6.2 × 104 TWh, which can be economically reused through processes such as hydrothermal liquefaction [102,103] or anaerobic digestion [104] to obtain high-density energy carbon (hydrocarbons and methane, respectively) and a liquor or effluent rich in macronutrients. This effluent can be incorporated into PBRs [95,105], allowing for a closed cycle to reuse these elements in growth. It is necessary to highlight that this use would return part of the fixed CO2 to the atmosphere.
- (c)
- Animal Feed: Over 6 Gt/year of forage and grain are consumed to sustain livestock worldwide, producing 340 Mt of meat and 800 Mt of milk, among other products [106]. To achieve these numbers, about 40% of the world’s cultivated land is used [107] and is not available for cultivating human food. Phytoplankton is a suitable food source for complementing and sometimes substituting livestock feed for ruminants and monogastric [108]. Microalgae and cyanobacteria are rich in vegetable proteins and high in omega-3 fatty acids, antioxidants, and other compounds, making them not only a source of macronutrients but also a source of compounds that improve the health and vitality of farmed animals [109,110,111].
- (d)
- Human Feed: Similarly, the obtained biomass could be an alternative source of nutrients and healthy compounds for human consumption. Phytoplankton is rich in essential amino acids and has a high content of polyunsaturated fatty acids and other beneficial compounds for human health [112]. Currently, 230 Mt of dry protein is consumed annually in our society [113]. To reach this amount, heavy dependence on animal protein is necessary, with environmental and animal welfare consequences. Using phytoplankton as a source of protein could reduce the dependence on animal protein and contribute to a more sustainable and healthy food system.
5. Discussion
- (a)
- A specialised scientific–technical sector should be implemented to design and propose suitable regions for establishing these CO2 sinks would be necessary. This sector should standardise the protocol for defining a region as suitable within a specific legal framework;
- (b)
- Measures that establish a profitable sector from the culture and harvest of phytoplankton and lead to the production of an attractive and valuable product for the market should be promoted;
- (c)
- Measures to develop a profitable industry for processing the large amount of biomass produced should be promoted, which can generate added value and demand for the product in such a way that transcends to other sectors, such as energy, human or animal food, or stored as fixed carbon;
- (d)
- International organizations and laws should be created to manage the use of fixed carbon and prevent its new release into the atmosphere.
6. Conclusions
- Since the Industrial Revolution, 292 GtC have accumulated in the atmosphere, equivalent to 1072 Gt of CO2, and this number continues to increase annually at a growing rate of 5.1 GtC per year (18.75 Gt of CO2). This phenomenon is altering several of the planet’s physicochemical parameters and cycles, such as temperature and the pH of large bodies of water, whose balance is vital for the stability of the vast majority of global ecosystems.
- The measures proposed by our civilization through various policies and international treaties have been slow and ineffective, focusing only on reducing emissions without implementing technologies to neutralize the excess CO2. The window for intervention with a guarantee of success is closing.
- The mass cultivation of phytoplankton is presumed to be a mature technology capable of utilising and accelerating the natural cycles of CO2 capture. Based on the literature reviewed in this work, an average CO2 capture value of 34 g/m2·day, equivalent to a biomass production rate of 17 g/m2·day, is proposed for cultivated areas.
- From this value, it can be theorised that to neutralise the 18.75 Gt/year of CO2 that accumulates in the atmosphere, an area of 2.1 × 106 km2 would need to be cultivated, equivalent to 0.41% of the planet’s total surface or 1.41% of the land surface. These figures would imply a 6.5% increase in the global NPP value.
- This work also analyses the challenges that civilization would face in implementing phytoplankton cultivation technology on a global scale. It suggests a horizontal PBR model with a water depth of 0.5 m placed in coastal areas, completely isolated from other natural environments and ecosystems. A low level of control is suggested for these partially natural areas, involving only recurrent inoculations with nitrogen-fixing species, aeration, and control of key nutrients (N, P, and Fe). Proper management of the generated biomass, whether through blocking or re-oxidation, would be crucial for maintaining the carbon cycle in a balanced state suitable for the stability of the biosphere.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Summary for Policymakers. In Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Front Matter; Cambridge University Press: Cambridge, UK, 2023; pp. 3–32. [Google Scholar] [CrossRef]
- Head, M.J.; Steffen, W.; Fagerlind, D.; Waters, C.N.; Poirier, C.; Syvitski, J.; Zalasiewicz, J.A.; Barnosky, A.D.; Cearreta, A.; Jeandel, C.; et al. The Great Acceleration is real and provides a quantitative basis for the proposed Anthropocene Series/Epoch. Episodes 2022, 45, 359–376. [Google Scholar] [CrossRef]
- Nag, B.; Makaranga, A.; Kareya, M.S.; Nesamma, A.A.; Jutur, P.P. Photosynthetic Cell Factories, a New Paradigm for Carbon Dioxide (CO2) Valorization. In Green Sustainable Process for Chemical and Environmental Engineering and Science; Inamuddin, A.M.A., Isloor, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2023; pp. 463–480. [Google Scholar] [CrossRef]
- Rapf, R.; Vaida, V. Sunlight as an Energetic Driver in the Synthesis of Molecules Necessary for Life. Phys. Chem. Chem. Phys. 2016, 18, 20067–20084. [Google Scholar] [CrossRef] [PubMed]
- Ruban, A. Evolution under the sun: Optimizing light harvesting in photosynthesis. J. Exp. Bot. 2015, 66, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.P.; Wood, A.P. The Chemolithotrophic Prokaryotes. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 753–793. [Google Scholar] [CrossRef]
- Hammarström, L. Overview: Capturing the sun for energy production. Ambio 2012, 41, 103–1007. [Google Scholar] [CrossRef]
- Stocker, B.D.; Yu, Z.; Massa, C.; Joos, F. Holocene peatland and ice-core data constraints on the timing and magnitude of CO2 emissions from past land use. Proc. Natl. Acad. Sci. USA 2017, 114, 1492–1497. [Google Scholar] [CrossRef]
- Moriarty, P.; Honnery, D. Global bioenergy: Problems and prospects. Int. J. Glob. Energy Issues 2007, 27, 231–249. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Global Carbon and Other Biogeochemical Cycles and Feedbacks. In Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2023; pp. 673–816. [Google Scholar] [CrossRef]
- Friedlingstein, P. Global carbon budget 2022. Earth Syst. Sci. Data 2022, 14, 4811–4900. [Google Scholar] [CrossRef]
- Quetin, G.R.; Famiglietti, C.A.; Dadap, N.C.; Bloom, A.A.; Bowman, K.W.; Diffenbaugh, N.S.; Liu, J.; Trugman, A.T.; Konings, A.G. Attributing Past Carbon Fluxes to CO2 and Climate Change: Respiration Response to CO2 Fertilization Shifts Regional Distribution of the Carbon Sink. Glob. Biogeochem. Cycles 2023, 37, e2022GB007478. [Google Scholar] [CrossRef]
- Barber, J. Photosynthetic energy conversion: Natural and artificial. Chem. Soc. Rev. 2009, 38, 185–196. [Google Scholar] [CrossRef]
- BP. Statistical Review of World Energy 2022. 71th Ed. Available online: https://www.bp.com/content/dam/bp/business-sites/en/global/corporate/pdfs/energy-economics/statistical-review/bp-stats-review-2022-full-report.pdf (accessed on 13 January 2023).
- Chatterjee, A.; Gierach, M.; Sutton, A.; Feely, R.; Crisp, D.; Eldering, A.; Gunson, M.; O’Dell, C.; Stephens, B.; Schimel, D. Influence of El Niño on atmospheric CO2 over the tropical Pacific Ocean: Findings from NASA’s OCO-2 mission. Science 2017, 358, eaam5776. [Google Scholar] [CrossRef]
- Khatiwala, S.; Primeau, F.; Hall, T. Reconstruction of the history of anthropogenic CO2 concentrations in the ocean. Nature 2009, 462, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Dupont, S.; Pörtner, H.O. A snapshot of ocean acidification research. Mar. Biol. 2013, 160, 1765–1771. [Google Scholar] [CrossRef]
- Ballantyne, A.P.; Liu, Z.; Anderegg, W.R.; Yu, Z.; Stoy, P.; Poulter, B.; Vanderwall, J.; Watts, J.; Kelsey, K.; Neff, J. Reconciling carbon-cycle processes from ecosystem to global scales. Front. Ecol. Environ. 2021, 19, 57–65. [Google Scholar] [CrossRef]
- Hall, D.O.; Rao, K.K. Photosynthesis, 6th ed.; Cambridge University Press: Cambridge, UK, 1999; 214p. [Google Scholar]
- Davis, S.; Lebauer, D.; Long, S. Light to liquid fuel: Theoretical and realized energy conversion efficiency of plants using Crassulacean Acid Metabolism (CAM) in arid conditions. J. Exp. Bot. 2014, 65, eru163. [Google Scholar] [CrossRef] [PubMed]
- Edwards, E.J. Evolutionary trajectories, accessibility, and other metaphors: The case of C4 and CAM photosynthesis. New Phytol. 2019, 223, 1742–1755. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. What is the maximum efficiency with which photosynthesis can convert solar energy into biomass? Curr. Opin. Biotechnol. 2008, 19, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Vecchi, V.; Barera, S.; Bassi, R.; Dall’Osto, L. Potential and Challenges of Improving Photosynthesis in Algae. Plants 2020, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Treves, H.; Lucius, S.; Feil, R.; Stitt, M.; Hagemann, M.; Arrivault, S. Operation of Carbon-Concentrating Mechanisms in Cyanobacteria and Algae requires altered poising of the Calvin-Benson cycle. bioRxiv 2022. bioRxiv:2022.08.23.504937. [Google Scholar] [CrossRef]
- Singh, U.B.; Ahluwalia, A. Microalgae: A promising tool for carbon sequestration. Mitig. Adapt. Strateg. Glob. Change 2013, 18, 73–95. [Google Scholar] [CrossRef]
- Paul, S.; Bera, S.; Dasgupta, R.; Mondal, S.; Roy, S. Review on the Recent Structural Advances in Open and Closed Systems for Carbon Capture through Algae. Energy Nexus 2021, 4, 100032. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Biology of microalgae. In Microalgae in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2018; pp. 23–72. [Google Scholar] [CrossRef]
- Ciferri, O. Spirulina, the edible microorganism. Microbiol. Rev. 1983, 47, 551–578. [Google Scholar] [CrossRef] [PubMed]
- Dangeard, P. Sur une algue bleue alimentaire pour l’homme: Arthrospira platensis (Nordstedt) Gomont. Actes Soc. Linn. Boreaux Extr. Procés-Verbaux 1940, 91, 39–41. [Google Scholar]
- Geohegan, M. Unicellular Algae as a source of food. Nature 1951, 168, 426–427. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-value products from microalgae—Their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Huang, Q.; Jiang, F.; Wang, L.; Yang, C. Design of Photobioreactors for Mass Cultivation of Photosynthetic Organisms. Engineering 2017, 3, 318–329. [Google Scholar] [CrossRef]
- Wang, B.; Lan, C.Q.; Horsman, M. Closed photobioreactors for production of microalgal biomasses. Biotechnol. Adv. 2012, 30, 904–912. [Google Scholar] [CrossRef]
- Tredici, M.R.; Chini Zittelli, G.; Rodolfi, L. Photobioreactors. In Encyclopedia of Industrial Biotechnology. Bioprocess, Bioseparation, and Cell Technology; Flickinger, M.C., Ed.; Wiley: Hoboken, NJ, USA, 2009; pp. 1–15. [Google Scholar] [CrossRef]
- Oswald, W.J.; Golueke, C.G. Biological transformation of solar energy. Adv. Appl. Microbiol. 1969, 2, 223–262. [Google Scholar] [CrossRef]
- Brenner, A.; Abeliovich, A. Water purification: Algae in wastewater oxidation ponds. In Handbook of Microalgal Culture, 2nd ed.; Richmond, A., Hu, Q., Eds.; Wiley: Hoboken, NJ, USA, 2013; pp. 595–601. [Google Scholar] [CrossRef]
- Slegers, P.M.; Wijffels, R.H.; van Straten, G.; van Boxtel, A.J.B. Design Scenarios for Flat Panel Photobioreactors. Appl. Energy 2011, 88, 3342–3353. [Google Scholar] [CrossRef]
- Fernández, I.; Acién, F.G.; Berenguel, M.; Guzmán, J.L. First principles model of a tubular photobioreactor for microalgal production. Ind. Eng. Chem. Res. 2014, 53, 11121–11136. [Google Scholar] [CrossRef]
- Torzillo, G.; Chini Zittelli, G. Tubular Photobioreactors. In Algal Biorefineries; Prokop, A., Bajpai, R., Zappi, M., Eds.; Springer: Cham, Switzerland, 2015; pp. 105–116. [Google Scholar] [CrossRef]
- Nogueira, N.; Nascimento, F.; Cunha, C.; Cordeiro, N. Nannochloropsis gaditana grown outdoors in annular photobioreactors: Operation strategies. Algal Res. 2020, 48, 101913. [Google Scholar] [CrossRef]
- Ugwu, U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for Mass Cultivation of Algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Masojídek, J.; Ranglová, K.; Lakatos, G.E.; Silva Benavides, A.M.; Torzillo, G. Variables Governing Photosynthesis and Growth in Microalgae Mass Cultures. Processes 2021, 9, 820. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Silva, S.O.; Baptista, J.M.; Malcata, F.X. Light requirements in microalgal photobioreactors: An overview of biophotonic aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef] [PubMed]
- Parlevliet, D.; Moheimani, N. Efficient conversion of solar energy to biomass and electricity. Aquat. Biosyst. 2014, 10, 4. [Google Scholar] [CrossRef]
- Michaelis, L.; Menten, M.L.; Johnson, K.A.; Goody, R.S. The original Michaelis constant: Translation of the 1913 Michaelis-Menten paper. Biochemistry 2011, 50, 8264–8269. [Google Scholar] [CrossRef]
- Straka, L.; Rittmann, B.E. (Growth kinetics and mathematical modeling of Synechocystis sp. PCC 6803 under flashing light. Biotechnol. Bioeng. 2019, 116, 469–474. [Google Scholar] [CrossRef]
- Mairet, F.; Bayen, T. The promise of dawn: Microalgae photoacclimation as an optimal control problem of resource allocation. J. Theor. Biol. 2021, 515, 110597. [Google Scholar] [CrossRef]
- Ananthi, V.; Kathirvel, B.; Pugazhendhi, A.; Arun, A. Impact of abiotic factors on biodiesel production by microalgae. Fuel 2021, 284, 118962. [Google Scholar] [CrossRef]
- Barten, R.; Djohan, Y.; Evers, W.; Wijffels, R.; Barbosa, M. Towards industrial production of microalgae without temperature control: The effect of diel temperature fluctuations on microalgal physiology. J. Biotechnol. 2021, 336, 56–63. [Google Scholar] [CrossRef]
- Uyar, B.; Kapucu, N. Passive temperature control of an outdoor photobioreactor by phase change materials. J. Chem. Technol. Biotechnol. 2015, 90, 915–920. [Google Scholar] [CrossRef]
- Gao, K. Approaches and involved principles to control pH/pCO2 stability in algal cultures. J. Appl. Phycol. 2021, 33, 3497–3505. [Google Scholar] [CrossRef]
- Fox, J.M.; Zimba, P.V. Minerals and trace elements in microalgae. In Microalgae in Health and Disease Prevention; Levine, I.A., Fleurence, J., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 177–193. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Bittencourt Sydney, E.; Assú Tessari, L.F.; Soccol, C.R. Chapter 2—Culture media for mass production of microalgae. In Biofuels from Algae, 2nd ed.; Ashok Pandey, J.-S., Chang, C.R., Lee, D.-J., Chisti, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 33–50. [Google Scholar] [CrossRef]
- Yaakob, M.A.; Mohamed, R.M.S.R.; Al-Gheethi, A.; Aswathnarayana Gokare, R.; Ambati, R.R. Influence of Nitrogen and Phosphorus on Microalgal Growth, Biomass, Lipid, and Fatty Acid Production: An Overview. Cells 2021, 10, 393. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. On the factors, which determine massive beta-carotene accumulation in the halotolerant alga Dunaliella bardawil. Plant Physiol. 1983, 72, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Boussiba, S.; Vonshak, A. Astaxanthin accumulation in the green alga Haematococcus pluvialis. Plant Cell Physiol. 1991, 32, 1077–1082. [Google Scholar] [CrossRef]
- Drira, M.; Elleuch, J.; Ben Hlima, H.; Hentati, F.; Gardarin, C.; Rihouey, C.; Le Cerf, D.; Michaud, P.; Abdelkafi, S.; Fendri, I. Optimization of Exopolysaccharides Production by Porphyridium sordidum and Their Potential to Induce Defense Responses in Arabidopsis thaliana against Fusarium oxysporum. Biomolecules 2021, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Liyanaarachchi, V.; Premaratne, M.; Ariyadasa, T.; Nimarshana, V.; Malik, A. Two-stage cultivation of microalgae for production of high-value compounds and biofuels: A review. Algal Res. 2021, 57, 102353. [Google Scholar] [CrossRef]
- Hegemann, P. Vision in microalgae. Planta 1997, 203, 265–274. [Google Scholar] [CrossRef]
- Walsby, A.E. Gas vesicles. Microbiol. Rev. 1994, 58, 94–144. [Google Scholar] [CrossRef]
- St. Laurent, L.; Garrett, C. The Role of Internal Tides in Mixing the Deep Ocean. J. Phys. Oceanogr. 2022, 32, 2882–2899. [Google Scholar] [CrossRef]
- Chanquia, S.N.; Vernet, G.; Kara, S. Photobioreactors for cultivation and synthesis: Specifications, challenges, and perspectives. Eng. Life Sci. 2022, 22, 712–724. [Google Scholar] [CrossRef]
- Souza Kirnev, P.C.; Vandenberghe, L.P.d.S.; Soccol, C.R.; de Carvalho, J.C. Mixing and Agitation in Photobioreactors. In Innovations in Fermentation and Phytopharmaceutical Technologies; Thatoi, H., Mohapatra, S., Das, S.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 13–35. [Google Scholar] [CrossRef]
- Rogers, J.; Rosenberg, J.; Guzman, B.; Oh, V.; Mimbela, L.-E.; Ghassemi, A.; Betenbaugh, M.; Oyler, G. A critical analysis of paddlewheel-driven raceway ponds for algal biofuel production at commercial scales. Algal Res. 2013, 4, 76–88. [Google Scholar] [CrossRef]
- Hreiz, R.; Sialve, B.; Morchain, J.; Escudié, R.; Steyer, J.-P.; Guiraud, P. Experimental and numerical investigation of hydrodynamics in raceway reactors used for algaculture. Chem. Eng. J. 2014, 250, 230–239. [Google Scholar] [CrossRef]
- Fan, F.; Fei, Z.; Wan, M.; Huang, J.; Wang, W.; Bai, W.; He, M.; Li, Y. The optimization of centrifugal pump driving horizontal tubular photobioreactor for enhancing astaxanthin production using heterotrophic Haematococcus pluvialis. J. Biotechnol. 2021, 341, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Sánchez Mirón, A.; García Camacho, F.; Contreras Gómez, A.; Grima, E.M.; Chisti, Y. Bubble-column and airlift photobioreactors for algal culture. AIChE J. 2000, 46, 1872–1887. [Google Scholar] [CrossRef]
- Wang, C.; Lan, C.Q. Effects of shear stress on microalgae—A review. Biotechnol. Adv. 2018, 36, 986–1002. [Google Scholar] [CrossRef]
- Hoffman, J.; Pate, R.; Drennen, T.; Quinn, J. Techno-economic assessment of open microalgae production systems. Algal Res. 2017, 23, 51–57. [Google Scholar] [CrossRef]
- Schagerl, M.; Siedler, R.; Konopáčová, E.; Ali, S.S. Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements. Cells 2022, 11, 2455. [Google Scholar] [CrossRef]
- Cho, C.; Nam, K.; Seo, Y.H.; Kim, J.Y.; Lee, S.; Lee, Y.G.; Yang, J.W. Study of Optical Configurations for Multiple Enhancement of Microalgal Biomass Production. Sci. Rep. 2019, 9, 1723. [Google Scholar] [CrossRef]
- Illman, A.M.; Scragg, A.H.; Shales, S.W. Increase in Chlorella strains calorific values when grown in low nitrogen medium. Enzym. Microb. Technol. 2000, 27, 631–635. [Google Scholar] [CrossRef]
- Borowitzka, A.M.; Moheimani, R.N. Algae for Biofuels and Energy. In Developments in Applied Phycology; Preface; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; New York, NY, USA; London, UK, 2013; Volume 5. [Google Scholar] [CrossRef]
- De Vree, J.H.; Bosma, R.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H.; Lamers, P.P. Comparison of four outdoor pilot-scale photobioreactors. Biotechnol. Biofuels 2015, 8, 215. [Google Scholar] [CrossRef]
- BP. Statistical Review of World Energy 2021. 70th Ed. Available online: https://www.bp.com/content/dam/bp/business-sites/en/global/corporate/pdfs/energy-economics/statistical-review/bp-stats-review-2021-full-report.pdf (accessed on 13 January 2023).
- Ritchie, H.; Roser, M.; Rosado, P. Energy. Our World in Data 2022. Available online: https://ourworldindata.org/energy (accessed on 13 January 2023).
- Princiotta, F.T. The Climate Mitigation Challenge-Where Do We Stand? J. Air Waste Manag. Assoc. 2021, 71, 1234–1250. [Google Scholar] [CrossRef] [PubMed]
- Clémençon, R. The Two Sides of the Paris Climate Agreement: Dismal Failure or Historic Breakthrough? J. Environ. Dev. 2016, 25, 3–24. [Google Scholar] [CrossRef]
- Sayre, R. Microalgae: The Potential for Carbon Capture. BioScience 2010, 60, 722–727. [Google Scholar] [CrossRef]
- Prasad, R.; Gupta, S.K.; Shabnam, N.; Oliveira, C.Y.B.; Nema, A.K.; Ansari, F.A.; Bux, F. Role of Microalgae in Global CO2 Sequestration: Physiological Mechanism, Recent Development, Challenges, and Future Prospective. Sustainability 2021, 13, 13061. [Google Scholar] [CrossRef]
- Derakhshandeh, M.; Atici, T.; Tezcan Un, U. Evaluation of Wild-Type Microalgae Species Biomass as Carbon Dioxide Sink and Renewable Energy Resource. Waste Biomass Valorization 2021, 12, 105–121. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). Land Use in Agriculture by the Numbers 2020. 2020. Available online: https://www.fao.org/sustainability/news/detail/en/c/1274219/ (accessed on 22 March 2023).
- Ritchie, H.; Roser, M.; Rosado, P. China: CO2 country profile. Our World in Data 2020. Available online: https://ourworldindata.org/co2/country/china?country=~CHN (accessed on 22 March 2023).
- Ebhodaghe, S.O.; Imanah, O.E.; Ndibe, H. Biofuels from microalgae biomass: A review of conversion processes and procedures. Arab. J. Chem. 2022, 15, 103591. [Google Scholar] [CrossRef]
- Singh, G.; Patidar, S.K. Microalgae harvesting techniques: A review. J. Environ. Manag. 2018, 217, 499–508. [Google Scholar] [CrossRef]
- Carmichael, W.; Drapeau, C.; Anderson, D. Harvesting of Aphanizomenon flos-aquae Ralfs ex Born. & Flah. Var. flos-aquae (Cyanobacteria) from Klamath Lake for human dietary use. J. Appl. Phycol. 2000, 12, 585–595. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.; Verspagen, J.; Visser, P. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Olofsson, M.; Suikkanen, S.; Kobos, J.; Wasmund, N.; Karlson, B. Basin-specific changes in filamentous cyanobacteria community composition across four decades in the Baltic Sea. Harmful Algae 2020, 91, 101685. [Google Scholar] [CrossRef]
- Olofsson, M.; Klawonn, I.; Karlson, B. Nitrogen fixation estimates for the Baltic Sea indicate high rates for the previously overlooked Bothnian Sea. Ambio 2021, 50, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Falcón, L.I.; Cipriano, F.; Chistoserdov, A.Y.; Carpenter, E.J. Diversity of diazotrophic unicellular cyanobacteria in the tropical North Atlantic Ocean. Appl. Environ. Microbiol. 2002, 68, 5760–5764. [Google Scholar] [CrossRef] [PubMed]
- Pruvost, J.; Goetz, V.; Artu, A.; Das, P.; Aljabri, H. Thermal modeling and optimization of microalgal biomass production in the harsh desert conditions of State of Qatar. Algal Res. 2019, 38, 101381. [Google Scholar] [CrossRef]
- Schmitt, R. Salinity and the Global Water Cycle. Oceanography 2008, 21, 12–19. [Google Scholar] [CrossRef]
- De Luca, R.; Bezzo, F.; Béchet, Q.; Bernard, O. Meteorological Data-Based Optimal Control Strategy for Microalgae Cultivation in Open Pond Systems. Complexity 2019, 2019, 4363895. [Google Scholar] [CrossRef]
- Tian, H.; Bian, Z.; Shi, H.; Qin, X.; Pan, N.; Lu, C.; Pan, S.; Tubiello, F.N.; Chang, J.; Conchedda, G.; et al. History of anthropogenic Nitrogen inputs (HaNi) to the terrestrial biosphere: A 5 arcmin resolution annual dataset from 1860 to 2019. Earth Syst. Sci. Data 2022, 14, 4551. [Google Scholar] [CrossRef]
- Mahata, C.; Mishra, S.; Dhar, S.; Ray, S.; Mohanty, K.; Das, D. Utilization of dark fermentation effluent for algal cultivation in a modified airlift photobioreactor for biomass and biocrude production. J. Environ. Manag. 2022, 330, 11712. [Google Scholar] [CrossRef]
- Kumar, K.; Mella-Herrera, R.A.; Golden, J.W. Cyanobacterial heterocysts. Cold Spring Harb. Perspect. Biol. 2010, 2, a000315. [Google Scholar] [CrossRef]
- Qadir, M.; Drechsel, P.; Jiménez Cisneros, B.; Kim, Y.; Pramanik, A.; Mehta, P.; Olaniyan, O. Global and regional potential of wastewater as a water, nutrient and energy source. Nat. Resour. Forum 2020, 44, 40–51. [Google Scholar] [CrossRef]
- Wang, L.; Qin, T.; Zhao, J.; Zhang, Y.; Wu, Z.; Cui, X.; Zhou, G.; Li, C.; Guo, L.; Jiang, G. Exploring the nitrogen reservoir of biodegradable household garbage and its potential in replacing synthetic nitrogen fertilizers in China. PeerJ 2022, 10, e12621. [Google Scholar] [CrossRef]
- Udayan, A.; Sirohi, R.; Sreekumar, N.; Sang, B.-I.; Sim, S.J. Mass Cultivation and Harvesting of Microalgal Biomass: Current Trends and Future Perspectives. Bioresour. Technol. 2022, 344, 126406. [Google Scholar] [CrossRef] [PubMed]
- Hachicha, R.; Elleuch, F.; Ben Hlima, H.; Dubessay, P.; de Baynast, H.; Delattre, C.; Pierre, G.; Hachicha, R.; Abdelkafi, S.; Michaud, P.; et al. Biomolecules from Microalgae and Cyanobacteria: Applications and Market Survey. Appl. Sci. 2022, 12, 1924. [Google Scholar] [CrossRef]
- Onyeaka, H.; Miri, T.; Kechrist, O.; Hart, A.; Anumudu, C.; Al-sharify, Z.T. Minimizing carbon footprint via microalgae as a biological capture. Carbon Capture Sci. Technol. 2021, 1, 100007. [Google Scholar] [CrossRef]
- Han, Y.; Hoekman, S.; Cui, Z.; Jena, U.; Das, P. Hydrothermal liquefaction of marine microalgae biomass using co-solvents. Algal Res. 2019, 38, 101421. [Google Scholar] [CrossRef]
- Aliyu, A.; Lee, J.; Harvey, A. Microalgae for biofuels: A review of thermochemical conversion processes and associated opportunities and challenges. Bioresour. Technol. Rep. 2021, 15, 103591. [Google Scholar] [CrossRef]
- Ganesh Saratale, R.; Kumar, G.; Banu, R.; Xia, A.; Periyasamy, S.; Saratale, G.D. A critical review on anaerobic digestion of microalgae and macroalgae and co-digestion of biomass for enhanced methane generation. Bioresour. Technol. 2018, 262, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Romero, A.; Martin, M.; Boyer, A.; Bolzoni, R.; Matricon, L.; Sassi, J.F.; Steyer, J.P.; Delrue, F. Microalgae adaptation as a strategy to recycle the aqueous phase from hydrothermal liquefaction. Bioresour. Technol. 2023, 371, 128631. [Google Scholar] [CrossRef]
- Ritchie, H.; Rosado, P.; Roser, M. Meat and Dairy Production. Our World in Data 2017. Available online: https://ourworldindata.org/meat-production (accessed on 22 March 2023).
- Mottet, A.; De Haan, C.; Falcucci, A.; Tempio, G.; Opio, C.; Gerber, P.J. Livestock: On our plates or eating at our table? A new analysis of the feed/food debate. Glob. Food Secur. 2017, 14, 1–7. [Google Scholar] [CrossRef]
- Kusmayadi, A.; Leong, Y.K.; Yen, H.W.; Huang, C.Y.; Chang, J.S. Microalgae as sustainable food and feed sources for animals and humans—Biotechnological and environmental aspects. Chemosphere 2021, 271, 129800. [Google Scholar] [CrossRef]
- Madeira, M.; Cardoso, C.; Lopes, P.; Coelho, D.; Afonso, C.; Bandarra, N.; Prates, J. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 47–59. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Cabrita, A.R.J.; Maia, M.R.G.; Valente, I.M.; Engrola, S.; Fonseca, A.J.M.; Ribeiro, D.M.; Lordelo, M.; Martins, C.F.; Cunha, L.F.; et al. Microalgae as feed ingredients for livestock production and aquaculture. In Microalgae; Academic Press: Cambridge, MA, USA, 2021; pp. 239–312. [Google Scholar] [CrossRef]
- Saadaoui, I.; Rasheed, R.; Aguilar, A.; Cherif, M.; Al Jabri, H.; Sayadi, S.; Manning, S.R. Microalgal-based feed: Promising alternative feedstocks for livestock and poultry production. J. Anim. Sci. Biotechnol. 2021, 12, 76. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Hassan, S.W.; Banat, F. An overview of microalgae biomass as a sustainable aquaculture feed ingredient: Food security and circular economy. Bioengineered 2022, 13, 9521–9547. [Google Scholar] [CrossRef] [PubMed]
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: Strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef]
- Bele, V.; Rajagopal, R.; Goyette, B. Closed-loop bioeconomy opportunities through the integration of microalgae cultivation with anaerobic digestion: A critical review. Bioresour. Technol. Rep. 2023, 21, 101336. [Google Scholar] [CrossRef]
- Rajagopal, R.; Mousavi, S.E.; Goyette, B.; Adhikary, S. Coupling of Microalgae Cultivation with Anaerobic Digestion of Poultry Wastes: Toward Sustainable Value Added Bioproducts. Bioengineering 2021, 8, 57. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, J.; Chen, P.; Ji, C.; Kang, Q.; Lu, B.; Li, K.; Liu, J.; Ruan, R. Bio-mitigation of carbon dioxide using microalgal systems: Advances and perspectives. Renewable and Sustainable. Energy Rev. 2017, 76, 1163–1175. [Google Scholar] [CrossRef]
- Ighalo, J.O.; Dulta, K.; Kurniawan, S.B.; Omoarukhe, F.O.; Ewuzie, U.; Eshiemogie, S.O.; Ojo, A.U.; Sheikh Abdullah, S.R. Progress in microalgae application for CO2 sequestration. Clean. Chem. Eng. 2022, 3, 100044. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Wang, X.; Jin, G.; Pan, K.; Wang, Y.; Li, Q.; Zhang, H. Effects of fluctuating temperature in open raceway ponds on the biomass accumulation and harvest efficiency of Spirulina in large-scale cultivation. Environ. Sci. Pollut. Res. 2021, 28, 20794–20802. [Google Scholar] [CrossRef]
- Acién Fernández, F.G.; González-López, C.V.; Fernández-Sevilla, J.M.; Molina-Grima, E. Conversion of CO2 into biomass by microalgae: How realistic a contribution may it be to significant CO2 removal? Appl. Microbiol. Biotechnol. 2012, 96, 577–586. [Google Scholar] [CrossRef]
- Leena, M.C.; Hausrath, E.M.; Ming, D.W.; Adcock, C.T.; Raymond, J.; Remias, D.; Ruemmele, W.P. Investigating the Growth of Algae Under Low Atmospheric Pressures for Potential Food and Oxygen Production on Mars. Front. Microbiol. 2021, 12, 733244. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Emissions (Mt CO2/Year) | Total Surface (×106 km2) | Proportional Culture Surface (×103 km2) | % Total Surface |
|---|---|---|---|---|
| China | 10,065 | 9.60 | 494.9 | 5.2 |
| USA | 5416 | 9.83 | 266.3 | 2.7 |
| India | 2654 | 3.29 | 130.5 | 4.0 |
| Russia | 1711 | 17.10 | 84.1 | 0.5 |
| Japan | 1162 | 0.38 | 57.1 | 15.1 |
| Germany | 759 | 0.36 | 37.3 | 10.4 |
| Iran | 720 | 1.65 | 35.4 | 2.1 |
| South Korea | 659 | 0.10 | 32.4 | 32.4 |
| Saudi Arabia | 621 | 2.15 | 30.5 | 1.4 |
| Indonesia | 615 | 1.90 | 30.2 | 1.6 |
| Canada | 568 | 9.98 | 27.9 | 0.3 |
| Mexico | 477 | 1.97 | 23.5 | 1.2 |
| South Africa | 468 | 1.22 | 23.0 | 1.9 |
| Brazil | 457 | 8.52 | 22.5 | 0.3 |
| Turkey | 428 | 0.78 | 21.0 | 2.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafrilla, B.; Matarredona, L.; Bonete, M.-J.; Zafrilla, G.; Esclapez, J. Phytoplankton as CO2 Sinks: Redirecting the Carbon Cycle. Appl. Sci. 2024, 14, 8657. https://doi.org/10.3390/app14198657
Zafrilla B, Matarredona L, Bonete M-J, Zafrilla G, Esclapez J. Phytoplankton as CO2 Sinks: Redirecting the Carbon Cycle. Applied Sciences. 2024; 14(19):8657. https://doi.org/10.3390/app14198657
Chicago/Turabian StyleZafrilla, Basilio, Laura Matarredona, María-José Bonete, Guillermo Zafrilla, and Julia Esclapez. 2024. "Phytoplankton as CO2 Sinks: Redirecting the Carbon Cycle" Applied Sciences 14, no. 19: 8657. https://doi.org/10.3390/app14198657