Abstract
Six wild-type non-Saccharomyces strains, belonging to the species Zygosaccharomyces bailii, Priceomyces carsonii, Trigonopsis californica, and Pichia manshurica, were evaluated for white wine production using Assyrtiko grapes from Santorini in Greece. Fermentation kinetics, in terms of glucose and fructose consumption and sensory analysis, was first employed to test the enological potential of the yeast strains. Based on their performance, two strains of Z. bailii (Zb-A19Y5 and Zb-K29Y2) and one strain of T. californica (Tc-A9Y1) selected for further analysis. The selected strains were tested in larger fermentation volumes for sugar consumption, while the produced wines were assessed for classical enological parameters, volatile compounds (GC/MS), and sensory analysis. Tc-A9Y1 could lead to dry wine (1 g/L of residual sugars) with 1.6 vol (12%) less alcohol than the other experimental wines. The wines that were fermented with the strain Zb-K29Y2 exhibited very high concentrations of isoamyl alcohol (604.33 ± 76.8 mg/L), but at the same time, they were characterized by low fruity notes. None of the produced wines presented any off-flavor aromas. Exploiting non-Saccharomyces strains with great fermentation capacity, which are able to produce high-quality wines and adapted to global warming conditions, is a new challenge for the wine industry.
1. Introduction
Assyrtiko is a Greek grape variety with unique sensorial attributes. This dynamic variety is famous worldwide due to the production of high-acidity wines, with typical varietal aromas, even more so when grown in the volcanic terroir of Santorini Island [1]. Assyrtiko is considered a variety that is well adapted to climate change, characterized by resistance to heat and drought. Recently, a great collection of wine yeast has been collected from Greek wines at the end of the fermentation process, including Assyrtiko wines [2]. The wine market is becoming extremely competitive, so an effort to differentiate and create a signature wine style is vital [3]. The role of microorganisms constitutes an integral modulator of wine quality [4,5]. Modern winemaking seeks and invests in the selection of so-called indigenous, autochthonous, or native local strains. Indigenous strains are believed to be able to maintain the typical sensory properties and enhance the unique properties of a wine, demonstrating improved adaptation to their native environment [6]. Complexity can be added through the inoculation of various species either under co-inoculation or sequential inoculation schemes [7,8,9].
Non-Saccharomyces yeasts are usually found in the early stages of alcoholic fermentation, and the most dominant ones belong to the genera of Hanseniaspora, Candida, Torulaspora, Pichia, Rhodotorula, and Zygosaccharomyces [4,10]. These non-Saccharomyces yeasts may play a positive role in wine quality through the production of secondary metabolites with enhanced sensorial contribution [11,12,13]. A new application of non-Saccharomyces has emerged in terms of bioprotection, aiming for a reduction in sulfite in wines [11]. Furthermore, their ability to increase the release of volatile thiols, particularly during the pre-fermentation stage, when this microbiological sub-population is dominant, was previously noted [12]. In any case, their low fermentation capacity and high sensitivity to alcohol make them inappropriate starters for wine fermentation. Thus, mixed cultures with S. cerevisiae strains designed, leading to more or less complex wines based on the yeast strains and winemaking conditions [14,15,16]. Non-Saccharomyces yeast can also be classified as spoilage wine agents. The species that are most at risk of provoking organoleptic deviation is Brettanomyces bruxellensis, through the production of active odorous compounds that are described as “horse sweat”, “animal”, “plastic”, etc. Volatile phenols are the main compounds implicated in the spoilage effect, which can also be produced by genera such as Candida, Pichia, and Trigonopsis [17,18,19]. In more detail, Trigonopsis species were shown to be able to grow in the presence of ethanol by expressing heat shock proteins and a DNA damage-related protein and to be able to confer spoilage characteristics on wine [17]. Additionally, Pichia species that are recovered from grapes, grape juice, and winery equipment that is in contact with grape juice, but not from wines, produced 4-ethylphenol, an unpleasant taint described as a ‘‘phenolic odor” [19].
The present study aims to evaluate the performance of indigenous and rarely isolated non-Saccharomyces strains in monoculture fermentation schemes for their enological potential. Namely, the previously isolated species Zygosaccharomyces bailii, Priceomyces carsonii, Trigonopsis californica, and Pichia manshurica [2] were tested for their fermentation capacity and their sensorial contribution to Assyrtiko must. All the aforementioned species, based on the bibliography, are often correlated with microbial spoilage and have never before been evaluated for any beneficial potential.
2. Materials and Methods
2.1. Yeast Isolates
Seventeen microbial isolates belonging to 5 different species were previously isolated and identified in wines at the end of spontaneous alcoholic fermentation (AF) [2]. More specifically, Trigonopsis californica (1 isolate), Zygosaccharomyces bailii (8 isolates), Priceomyces carsonii (1 isolate), and Pichia manshurica (7 isolates) survived and co-existed with S. cerevisiae strains during AF. All the isolates were preserved at −20 °C in YPD broth [(g/L): yeast extract 10, Bacteriological peptone 20, Dextrose 20] with 30% of glycerol added.
2.2. Genotyping by Rep-PCR
Total genomic DNA from the isolates was extracted in previous work and stored at −20 °C [2]. Quantification and quality control of DNA extract were performed by spectrophotometer (Epoch, Biotek, Winooski, VT, USA) at wavelengths of 260, 280, and 230 nm. Prior to determination of the isolated yeast strains, rep-PCR (Repetitive Polymerase Chain Reaction) with GTG5 (5′-GTG GTG GTG GTG GTG-3′) primer was performed.
PCR amplification was conducted in 20 μL final reaction volumes, containing 10 µL of OneTaq-quick load 2× Mastermix with standard buffer (New England Biolands, Hitchin, UK), 10 mM GTG5, and 50 ng of template DNA. The amplification program consisted of 2 min of initial denaturation at 95 °C, 35 cycles of 30 s at 94 °C, 1 min at 45 °C, 4 min at 72 °C, concluding with 5 min at 65 °C. All PCR products were run on a 1.5% (w/v) agarose gel in 1 × TAE buffer, stained with ethidium bromide (20 min), at 110 V for 140 min and scanned under ultraviolet light (MiniBIS, DNR, Jerusalem, Israel). A 1Kb DNA ladder (Nippon Genetics, Düren, Germany) served as size standard. The resulting fingerprints were digitally captured, converted, normalized, and analyzed using the Dice coefficient with Bionumerics software version 6.1 (Applied Maths, Sint-Martens-Latem, Belgium). Means of the Unweighted Pair Group Method using the Arithmetic Average (UPGMA) clustering algorithm led to the formation of the strain-specific dendrogram.
2.3. Experimental Design and Winemaking Conditions
2.3.1. Fermentation Kinetics of Six Strains
Micro-fermentation trials evaluated the fermentation potential of the yeast strains. Fermentations were carried out in 50 mL of pasteurized (72 °C, 10 min) Assyrtiko must, which was provided by Gaia (Santorini, Greece) winery (vintage 2021), under static conditions. The initial pasteurized grape must (pH = 3.20 ± 0.03, total acidity = 5.77 ± 0.06 g tartaric acid/L, YAN = 609.00 ± 4.76 mgN2/L, 16.60 ± 0.01 mgSO2, 5.10 ± 0.10 free SO2) contained 119.5 ± 2.5 g/L of glucose and 120.1 ± 3.1 g/L of fructose. Precultures were grown in YPD broth at 28 °C for 48 h, and then used to inoculate each fermentation (106 cell/mL). The viability of the inoculum was examined by classical microbiological techniques at 0 and 48 h. The pasteurized must was also microbiologically analyzed, and no viable, culturable cells were noticed, ensuring the success of pasteurization. Briefly, 1 mL of the wine or must sample was serially diluted in 9 mL Ringer’s solution following decimal dilutions. Then, 0.1 mL of the appropriate dilutions spread in duplicate on YPD agar [(g/L): yeast extract 10, Bacteriological peptone 20, Dextrose 20, Agar 20] substrate to quantify the microbial population. All assessed fermentations duplicated, and each analysis conducted twice.
Fermentation kinetics were monitored by residual sugar (glucose and fructose) determination on a daily basis using Enzytec kit-liquid Glucose-Fructose (R-biopharm, Darmstadt, Germany). Fermentations were carried out at 18 °C in 100 mL Duran under static conditions. The temperature condition was chosen so as to be close to the real winemaking conditions. As previously described, cv. Assyrtiko fermentations are usually performed at 18 °C [20]. The sample was homogenized before every sampling point. Double trials of pure cultures were carried out for each different strain.
2.3.2. Fermentation Kinetics of Three Selected Strains
The strains with higher enological potential were further subjected to bigger laboratory fermentations (1.2 L total volume). The fermentation media and inoculum preparation were the same as described before Section 2.1. Fermentations were also carried out at 18 °C in 1.5 L glass bottles under static conditions. The sample was homogenized before every sampling point. Fermentation kinetics were monitored by residual sugar (glucose and fructose) determination a daily using an automatic enzymatic Analyzer (Hyperlab, Steroglass, San Martino in Campo, Italy). Two trials of pure cultures were carried out for each different strain.
2.4. Chemical Analysis
2.4.1. Classical Enological Parameters
Alcoholic fermentation was monitored daily through the determination of glucose and fructose. AF was considered completed when the total sugar concentration was below 4 g/L. Analysis of glucose/fructose, acetic acid, and malic acid was determined using an automatic enzymatic Analyzer (Hyperlab, Steroglass, San Martino in Campo, Italy). pH was measured using a pHmeter (HI2210, Hanna Instruments, Smithfield, VA, USA). Yeast assimilable nitrogen (YAN) was measured following the formol method [21]. Determination of free and total SO2 was performed by the iodometric method, and ethanol content and total acidity were determined according to OIV methods [22]. All duplicated fermentations were analyzed twice.
2.4.2. Identification and Quantification of Volatile Compounds
The methodology selected for optimizing the extraction of aromatic compounds from the wine samples, preceding their introduction into the gas chromatography–mass spectrometry (GC-MS) apparatus, involved the implementation of a liquid–liquid extraction technique. This extraction approach was adapted from the protocol established previously [23]. All duplicated fermentations were analyzed twice.
The analytical instruments employed in this study included a gas chromatograph (Shimadzu (Kyoto, Japan), Nexis GC-2030) equipped with an autosampler/injector (Shimadzu (Kyoto, Japan), AOC 20i Plus). In the case of liquid–liquid extraction (LLE), the injector was maintained at a temperature of 250 °C, operating in split/splitless mode with a 1/100 split ratio. THE MEGA-WAX MS capillary column was utilized, measuring 30 m in length, with an i.d. of 0.25 mm and a film thickness of 0.25 μm. The stationary phase consisted of polyethylene glycol (PEG) (Agilent Technologies, Santa Clara, CA, USA). The injector temperature was maintained at 250 °C. The temperature program for the gas chromatograph commenced at 50 °C for 2.5 min, followed by a ramping rate of 2.5 °C/min to 100 °C, 4 °C/min to 165 °C, and 7 °C/min to 250 °C, with a 2 min hold at the final temperature. Helium served as the carrier gas, with a constant flow rate of 1.5 mL/min.
The mass spectrometer was operated in electron impact mode at 70 eV within the mass range of m/z 50–550 amu, while maintaining the ion source and transfer line at 230 °C. All analyses were conducted in duplicate. In accordance with established protocols, concentrations of volatile compounds were determined utilizing an internal standard (n-undecane). Compound identification was validated when a minimum match factor of 80% was attained by comparing the mass spectra of the compounds to those available in the NIST library.
2.5. Sensory Analysis
The six produced wines from the fermentation of six strains of five species were categorized based on their aromatic profile (odor) using a free sorting task test, as described previously [2]. Free sorting task is an efficient technique for assessing the perception of a set of products by a panel of subjects and is also widespread in wine science [24,25,26]. Samples were evaluated in individual booths, using covered, ISO glasses (NF V09-110, 1971) containing about 25 mL of liquid, coded with three-digit random numbers. A total of 10 experienced panelists participated in the pilot study. Panelists were not informed about the nature of the samples. All duplicated fermentations were evaluated twice by each panelist (n = 4).
Sensory analysis of the selected strains was performed as described by Dimopoulou et al. [15]. Samples were evaluated by descriptive analysis in individual booths, using covered, ISO glasses (NF V09-110, 1971) containing about 25 mL of liquid, coded with three-digit random numbers [15]. The panel was composed of 10 trained tasters, selected for their experience in assessing aromas in Greek white wines. Panelists were not informed about the nature of the samples, and all duplicated fermentations were evaluated twice by each panelist (n = 4). The intensity of the examined sensory attributes was evaluated using a 10-point scale (1: null; 10: very strong) and scored manually.
2.6. Statistical Analysis
Significant differences were evaluated by One-way Analysis of Variance (ANOVA), which was followed by Tukey’s post hoc test (p < 0.05). All statistical analyses were performed using Statgraphics Centurion 18 software (Statgraphics Technologies, Inc., The Plains, VA, USA). Encoding free sorting data was the key to categorizing wine samples based on the results of the sensory assessment. For each group, results were encoded in an individual similarity matrix (wines × wines), in which 1 stands for two wines set in the same group and 0 for two wines placed in different groups. These individual matrices were summed across subjects; the resulting co-occurrence matrix represents the global similarity matrix, where larger numbers indicate higher similarity between samples. The assumption underlying this method is that grouped samples are more similar than samples that are sorted into different groups. The resulting co-occurrence matrix was submitted to HCA (ward coefficient) to derive a spatial arrangement of wines with R (3.6.2) software analysis.
3. Results and Discussion
During the spontaneous fermentation process, non-Saccharomyces yeasts usually dominate and start the AF, while the conversion of sugars into ethanol is often completed by S. cerevisiae strains [27,28]. However, all the 18 non-Saccharomyces yeasts that were investigated in this survey, presented in Table 1, are isolated from spontaneous fermented wines which exhibited no off-odor or off-flavor characteristics, as previously described [2]. The aforementioned species, which have rarely been isolated from the wine environment, are often characterized as spoilage yeasts. More precisely, T. californica is characterized by the literature as a contaminant of Californian wines and genetically closely related to Trigonopsis cantarellii, Trigonopsis variabilis, and Trigonopsis vinaria [29,30]. Recent research suggests that T. californica is considered a high acetic acid producer and has been isolated from wines at the end of AF and during aging, conferring a “Brett” character to wine [31,32]. Additionally, P. manshurica is found in several fermented foods and alcoholic drinks [33,34,35,36]. The spoilage capacity of P. manshurica regional isolates was evaluated in red wine, and the production of volatile phenols and cadaverine was evidenced [34,35,37].

Table 1.
Sample coding, species, and strain differentiation of the isolates. The selected strains are marked in bold.
However, only a few researchers have investigated their contribution to a wine’s microbial environment. For instance, Z. bailii stands out as a spoilage yeast, mainly in acidified food products, various non-carbonated fruit drinks, and in wine with fermentative metabolism [38,39,40]. Despite the higher levels of produced sorbic, benzoic, and especially acetic acid that are produced by Z. bailii compared to those produced by S. cerevisiae, no other off-flavor observations have been reported to indicate spoilage behavior in wines [40,41]. Finally, according to Cordero-Bueso et al. [42], P. carsonii was reported to influence the volatile composition of musts from grape berries and was not able to catabolize all residual sugars on must. P. carsonii has also been isolated from oils, fermented sausages, and lambic beer [43,44,45]. The main objective of the present survey was to examine the performance of these indigenous and rarely isolated non-Saccharomyces strains in monocultures’ fermentation schemes as potential starter cultures.
Strain differentiation was achieved using repetitive extragenic palindromic PCR (rep-PCR) using the primer GTG5 (Table 1). For isolates from the same species, which were also identified within the same wine samples, a lack of strain diversity was predominantly observed. Consequently, only one strain per species of T. californica, P. carsonii, and P. manshurica was identified. On the contrary, when investigating Z. bailii, nine isolates originating from various samples were analyzed, and molecular fingerprinting revealed the presence of three distinct strains. It is noteworthy that Zb-Z3 is detected in both red and white wines from two different wineries. A single representative strain for each species was subsequently selected for further investigation of its enological potential.
3.1. Yeast Growth and Enological Parameters during Fermentation
Fermentation kinetics were monitored for each single monoculture by sugar consumption. Figure 1 shows the glucose and fructose consumption during the AF of pasteurized Assyrtiko must under the different inoculation schemes. The microbiological analysis affirmed the initial inoculum concentration (0 h) and the survival of the inoculated strains at 48 h. The population of the inoculum in all modalities at 48 h ranged from 7.8 ± 0.02 to 8.2 ± 0.05 log CFU/mL. After 20 days of fermentation at 18 °C, two strains (Pc-K21Y9, Pm-K34Y10) could not lead to dry wines. Pc-K21Y9 demonstrated fructophilic behavior, since fructose was catabolized quickly, but it could not catabolize glucose, while the residual sugars were estimated at 24.7 ± 1.4 g/L. The term “fructophilic” refers to the preferential capacity of metabolizing fructose when both fructose and glucose are present in high concentrations in the surrounding environment [46]. Additionally, the strain Pm-K34Y10, after 20 days of AF, could not catabolize a concentration of 20.8 ± 2.2 g of glucose/L and 51.78 ± 3.0 g fructose/L. All fermentations that were performed with Z. bailii and T. californica strains led to dry wines. More specifically, Zb-A19Y5, Zb-K29Y2, Tc-A9Y1, and Zb-K16Y1 completed alcoholic fermentation at 12, 16, 20, and 16 days, respectively. A recent study investigated 20 indigenous S. cerevisiae’s fermentative capacity using the same pasteurized Assyrtiko must, under the same laboratory conditions [2]. The majority of these strains had completed alcoholic fermentation after 9 days. Consequently, the investigated non-Saccharomyces are characterized by a lower fermentation rate compared to S. cerevisiae starters. It is widely recognized that some non-Saccharomyces species are capable of surviving and persisting until the completion of fermentation, as they exhibit a heightened tolerance to ethanol [38]. Our results are in line with this observation, since the increment of alcohol did not significantly impact most of the species in this survey. Numerous non-Saccharomyces yeasts have been explored for their fermentative capabilities, primarily in co-inoculation with S. cerevisiae, given their inability to complete alcoholic fermentation when used as monocultures [37,40,47,48]. The fermentation rate of monocultures of non-Saccharomyces species has been proven to be lower than those with monocultures of S. cerevisiae, sequential inoculations, or co-cultures of non-Saccharomyces species with S. cerevisiae strains. Nevertheless, it is worth noting that the successful completion of alcoholic fermentation by monocultures of non-Saccharomyces species has already been reported [49,50]. Notably, certain strains of the Z. bailii species have been previously identified for their high fermentative efficiency, reaching up to 89% [49].
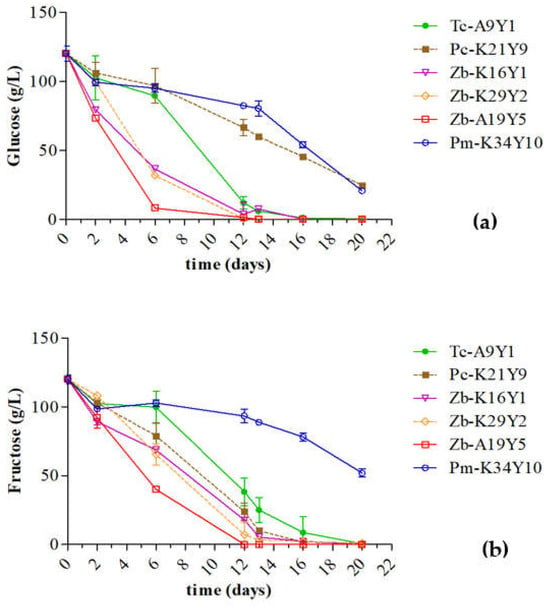
Figure 1.
Changes in sugar consumption [(a) D-glucose and (b) D-fructose] during alcoholic fermentations for the 6 strains. Sample coding: Tc = Trigonopsis californica, Zb = Zygosaccharomyces bailii, Pc = Priceomyces carsonii, Pm = Pichia manshurica.
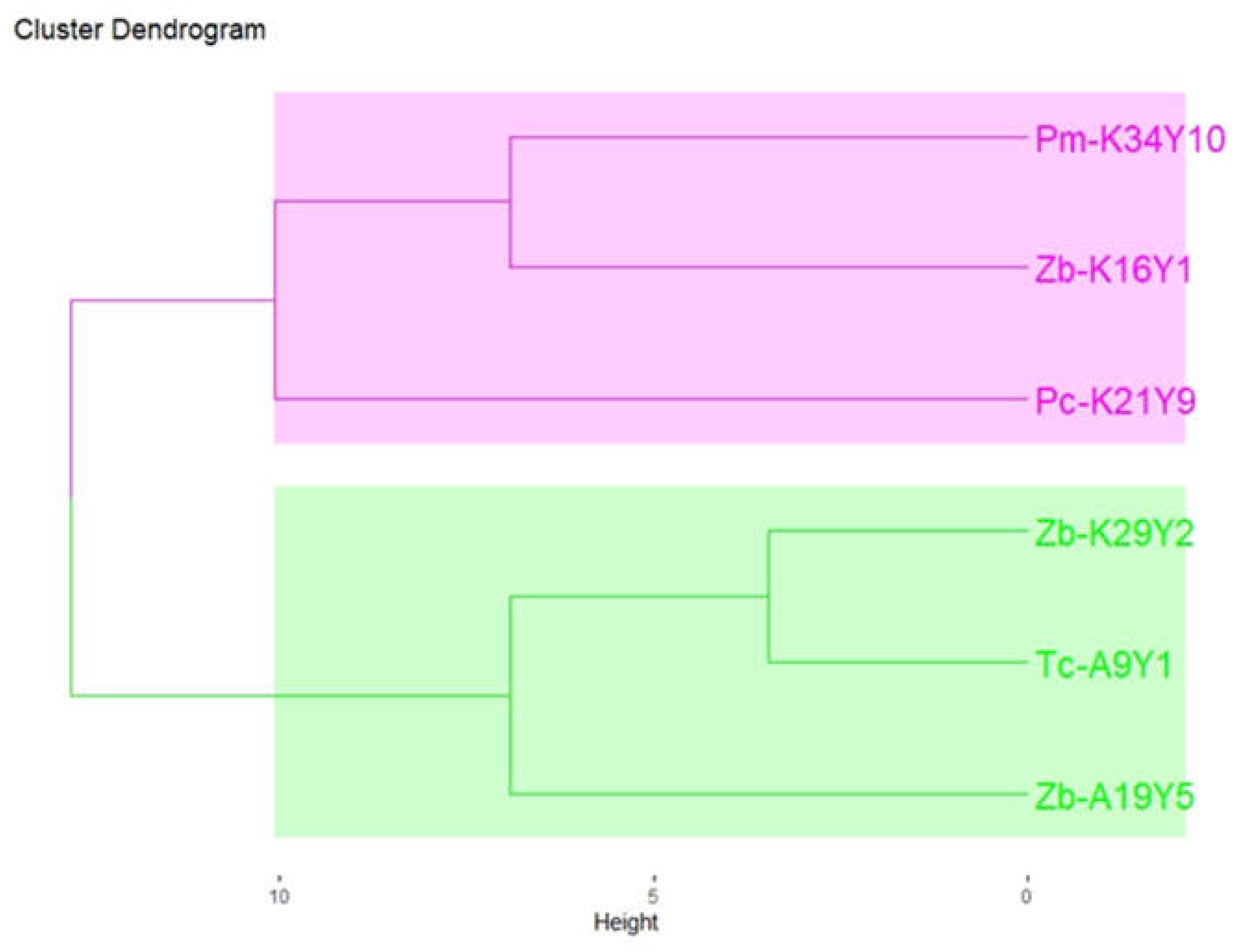
To provide a holistic and non-verbal evaluation of the resulting wines, a sensory analysis was performed. Based on the aroma bouquet of Assyrtiko wines, the determination of the differences among non-Saccharomyces fermentation schemes was achieved. According to the free sorting assessment, wines were grouped according to their perceived similarity by panelists, and sensory descriptors were provided for each group (Figure 2). Two distinct groups are noticed. Firstly, the pink group (Pm-K34Y10, Zb-K16Y1, and Pc-K21Y9) was described by mainly negative attributes (oxidation, H2S, and other off-odor aromas). Additionally, the green group (Zb-K29Y2, Tc-A9Y1, and Zb-A19Y5) was associated with positive attributes such as “fruitiness”, “floral”, “tropical fruits”, and “apple”. The results were expected due to the abnormal fermentation of Pm-K34Y10 and Pc-K21Y9 strains. However, Zb-K29Y2 was the only Z. bailii strain that exhibited off-flavor aromas. Our findings align with the literature that supports strains, belonging to the genera Hanseniaspora and Zygosaccharomyces, revealing differences at both the interspecies and intraspecies levels [37].
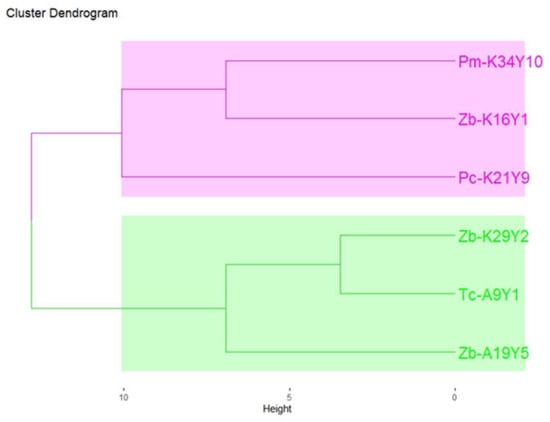
Figure 2.
Hierarchical cluster analysis (HCA) of the different aromatic profiles of the 6 produced wines from different non-Saccharomyces strains, based on the results of the free sorting task with Ward coefficient. Sample coding: Tc = Trigonopsis californica, Zb = Zygosaccharomyces bailii, Pc = Priceomyces carsonii, Pm = Pichia manshurica. The two groups are distinct by colors: Group A = pink, Group B = green.
To further explore the enological potential of the highest performing strains by performing more biochemical analyses, the strains that constituted the green group were subjected to a second laboratory fermentation in a bigger volume (1.2 L) so that a further analysis could be performed. The microbiological analysis affirmed the initial inoculum concentration (0 h) and the survival of the inoculated strains after 48 h of inoculation. The population of the inoculum in all modalities at 48 h varied between 7.9 ± 0.03 and 8.3 ± 0.04 log CFU/mL. As Figure 3 illustrates, the monocultures of the Z. bailii strains (Zb-A19Y5 and Zb-K29Y2) completed alcoholic fermentation within 504 h, while T. californica Tc-A9Y1 required 648 h for fermentation completion. Notably, in both fermentations, it is evident that the T. californica strain exhibited a lower fermentation rate compared to the Z. bailii strains. According to the literature, Z. bailii isolates have been noticed to exhibit fructophilic behavior, but it was difficult to conclude whether the observed phenotype could be linked to high sugar adaptation [38,46,51,52].
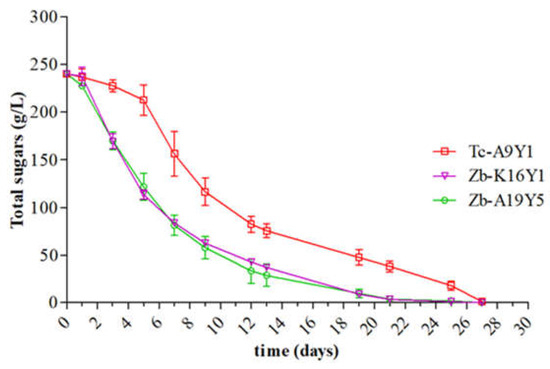
Figure 3.
Changes in total sugar consumption [D- glucose and D- fructose] during alcoholic fermentations for the two Z. bailii (Zb-K16Y1 and Zb-A19Y5) and one T. californica (Tc-A9Y1) strain. Sample coding: Tc = Trigonopsis californica, Zb = Zygosaccharomyces bailii.
3.2. Chemical Analyses
In Table 2, the standard enological parameters of the must before and after the completion of alcoholic fermentation are presented. The final concentration of total sugars was <4 g/L for each wine sample, 3.85 ± 1.16 g/L for Zb-A19Y5, and 3.65 ± 1.32 g/L for Zb-K29Y2 after 21 days, whereas for Tc-A9Y1, it was 1.00 ± 0.00 g/L after 27 days. According to the chemical analysis results, Z. bailii strains, namely, Zb-A19Y5 and Zb-K29Y2, led to the production of wines with a high alcohol content (13.41 ± 0.07%vol–13.11 ± 0.05%vol). In contrast, the T. californica (Tc-A9Y1) strain led to a reduced ethanol wine content (11.79 ± 0.04%vol). Moreover, the yield of ethanol that was produced per unit of total sugar consumed (YEtOH/TS, in g/g) was calculated. The ethanol yield of the two Z. bailii strains (0.45 and 0.44 g/g) was higher than those of T. californica (0.39 g/g). Using the Zb-A19Y5 strain, the ethanol yield was 88% of the maximum theoretical yield (0.51 g/g). The use of strains that lead to low final sugar values and a high alcohol yield is mostly wanted in winemaking. In Assyrtiko must, S. cerevisiae strains have also been tested with a higher alcohol yield (0.49–0.50 g/g) [53,54]. An innovative strategy to produce reduced-alcohol wines takes advantage of wine yeasts that do not belong to the S. cerevisiae species and that metabolize sugar without generating ethanol or do so with less efficiency [55].
Table 2.
Mean concentration of enological parameters of Assyrtiko must and wines produced using three different non-Saccharomyces strains (Zb-A19Y5, Tc-A9Y1, and Zb-K29Y2). Different letters in each row indicate the significance of One-way Analysis of Variance (ANOVA), which was followed by Tukey’s post hoc test (p < 0.05).
Furthermore, the pH value ranged between 3.12 ± 0.01 and 3.19 ± 0.01, with the highest value observed in the wine made with Tc-A9Y1. The total acidity of the wines ranged between 3.92 ± 0.02 and 4.35 ± 0.08 g tartaric acid/L, and yeast-assimilable nitrogen ranged between 99.40 ± 0.74 and 105.70 ± 3.01 mg N2/L, with the highest values observed in the wine made with Zb-A19Y5. The total SO2 in wines made with Z. bailii strains showed the same values of 14.10 ± 1.80 mg/L and 16.00 ± 4.51 mg/L as those where T. californica was inoculated. Low values of SO2 indicate the need for further protection of the produced wines from oxidation. Additionally, even if the values of free and total SO2 are not statistically significant, the higher value of combined SO2 of Tc-A9Y1 compared to Z. bailii strains could be correlated to a lower fermentation rate and possible commitment with the produced acetaldehyde. No statistically significant differences were found between the values of yeast-assimilable nitrogen and sulfur dioxide, whereas L-malic acid was not detected in any of the samples. Z. bailii is conventionally acknowledged as a malic acid-consuming yeast and has been suggested for adjusting pH in fermenting musts with an excess of malic acid [46,56]. Regarding the volatile acidity, the values of acetic acid ranged between 0.41 ± 0.02 and 0.47 ± 0.01, with the highest value observed in the wine made with Tc-A9Y1. The levels in wines made with Z. bailii strains were significantly different from those made with T. californica, but comparable to those obtained from wines inoculated with S. cerevisiae strains in Assyrtiko must [57].
3.3. Volatile Compound Analyses
More than a thousand aromatic compounds, encompassing higher alcohols, esters, organic acids, and other constituents, have been recognized in wine. While it is likely that not all of these compounds play a role in shaping the wine’s aroma, the diverse array of chemical families contributes to the remarkable aromatic complexity of wine. The wine samples were subjected to further analysis of some volatile compounds. These volatiles are potential contributors to the global aroma of wine. It has to be mentioned that Assyrtiko wines are mostly characterized by typical varietal aromas such as earthy, mushroom, and nutty odors, as well as lemon and honey [1]. Therefore, the impact of the inoculated strains on boosting the aromatic profile and typicity of the final product will be very important. Retention time and kovats index of the identified volatile aromatic compounds is reported in Table S1 (Supplementary Materials).
3.3.1. Higher Alcohols
The analysis of volatile alcohols in samples that were fermented with the strains Zb-A19Y5, Tc-A9Y1, and Zb-K29Y2 reveals intriguing insights into the distinct chemical signatures of each sample (Table 3). In general, interspecies differences among Z. bailii strains are noticed regarding the production of higher alcohols. In more detail, isoamyl alcohol demonstrates a significant difference, with sample Zb-K29Y2 exhibiting the highest concentration at 604.33 ± 76.80 mg/L. This discrepancy suggests that variations in isoamyl alcohol levels may contribute to the distinction in the flavor profiles of these wines. Isoamyl alcohol is generally described by banana- and fusel-type odors. A previous study demonstrates an amplification of butyric notes with the addition of isoamyl alcohol (3-methylbutan-1-ol) in a fruity model solution [58]. The literature provides evidence that higher alcohols are found in total concentrations ranging from 0.2 to 1.2 g/L in white wines [59]. While 2-phenylethan-1-ol imparts a rose-like aroma, other higher alcohols are often associated with fusel oil, solvent, or malt scents [60]. Research indicates that higher alcohols may enhance the aromatic complexity of wine or, in some instances, mask certain flavors, contingent upon their concentrations [61]. When present below 300 mg/L, higher alcohols are generally considered contributors to a desirable complexity of wine; however, concentrations exceeding 400 mg/L are deemed to have a negative impact on wine’s quality [61]. Phenylethyl alcohol, linked with floral notes, exhibits a pronounced concentration in the samples, ranging from 246.63 ± 15.09 mg/L to 319.92 ± 36.05 mg/L.
Table 3.
Higher alcohols (mg/L) of wines produced in pilot scale fermentations (±SD) with different non-Saccharomyces strains (Zb-A19Y5, Tc-A9Y1, and Zb-K29Y2). Different letters in each row indicate the significance of One-way Analysis of Variance (ANOVA), which was followed by Tukey’s post hoc test (p < 0.05).
Intriguingly, tryptophol, a secondary metabolite, also reveals noteworthy differences among the samples. Wines that were fermented with Zb-A19Y5 and Tc-A9Y1 strains were registered as having a concentration of 21.21 ± 0.81 mg/L and 20.62 ± 0.80 mg/L, respectively, whereas wines that were inoculated with Zb-K29Y1 differed significantly, with 8.76 ± 0.74 mg/L. This divergence underscores the role of tryptophol, which is a compound that is strongly impacted by yeast metabolism. According to previous research, aromatic alcohols such as phenylethanol contribute positively to the organoleptic characteristics of wines [62]. According to previous research, Z. bailii was found in Maotai-flavor liquor fermentation and enhanced the production of higher alcohols, which greatly impact the unique aroma [63].
3.3.2. Acids
Concerning the analysis of volatile acids in wine samples, several compounds reveal substantial variations in concentration among the three samples (Table 4). Wines that were inoculated with the Zb-A19Y5 strain showed the highest concentration of propanoic acid and propanoic acid, 2-methyl, which imparts an acidic, cheesy aroma, of octanoic acid, with its characteristic waxy notes, and finally oleic acid, which offers a slightly fatty aroma [64]. Meanwhile, the wine that was fermented with the Tc-A9Y2 yeast strain shows its significant differences in its hexanoic acid concentration, known for its pungent, sweaty odor, and octanoic acid, with its characteristic sweaty notes. Finally, the wine that was inoculated with the Zb-K29Y2 strain has the largest concentration of hexadecanoic acid, a fatty acid with the potential for a subtle wax or candle-like aroma, while pentadecanoic acid can contribute to a creamy and waxy aroma. These variations underscore the intricate and diverse nature of wine’s flavor and aroma, reflecting the complex interplay of these compounds in defining the sensory experience. It has been noted that fatty acids can influence the complexity of wine aromas, and this influence is contingent on the concentration of acids in the wine [65], although it is not linked to the overall quality of the wine [66]. For instance, at concentrations ranging from 4 to 10 mg/L, fatty acids may contribute to enhancing the complexity of wine, but the elegance of the wine aroma could be negatively impacted when their concentration exceeds 20 mg/L [67].
Table 4.
Volatile acids (mg/L) of wines produced in pilot scale fermentations (±SD) with different non-Saccharomyces strains (Zb-A19Y5, Tc-A9Y1, and Zb-K29Y2). Different letters in each row indicate the significance of One-way Analysis of Variance (ANOVA), which was followed by Tukey’s post hoc test (p < 0.05).
3.3.3. Esters
All the produced wines were exposed to volatile ester analysis. The wine samples presented the most evident differences in the concentration of octanoic acid, ethyl ester, and decanoic acid, ethyl ester (Table 5). It is known that esters not only contribute to the fruity aroma of young wines but also serve as important markers for fruity notes in wines of different ages [68,69]. Esters have been observed to influence the fruity aroma of wine, even at subthreshold levels, due to their intricate synergistic effects [68,70,71], highlighting their significant contribution to the overall aroma of wine. The big difference between the number of esters in each sample and the concertation highlights the crucial role of the yeasts [72]. For instance, octanoic acid, ethyl ester, was not detected in sample Zb-A19Y5, and Ethyl 9-decenoate was not detected in Tc-A9Y1, while the other samples exhibited a high concentration. Additionally, the fact that wines from the two Z. bailii strains showed statistically significant differences can imply intraspecies variability.
Table 5.
Esters (mg/L) of wines produced in pilot scale fermentations (±SD) with different non-Saccharomyces strains (Zb-A19Y5, Tc-A9Y1, and Zb-K29Y2). Different letters in each row indicate the significance of One-way Analysis of Variance (ANOVA), which was followed by Tukey’s post hoc test (p < 0.05).
3.4. Sensory Impact
The Assyrtiko wines that were produced using the different fermentation schemes described above were all evaluated by a sensory panel of experts. Figure 4 depicts the mean scores of ten sensory characteristics, with a maximum score of 10, of the wines that were inoculated with the different non-Saccharomyces pure cultures. The statistical analysis (ANOVA) indicated a significant difference (p < 0.05) for the fruitiness, citrus, and aromatic aftertaste of the produced wines.
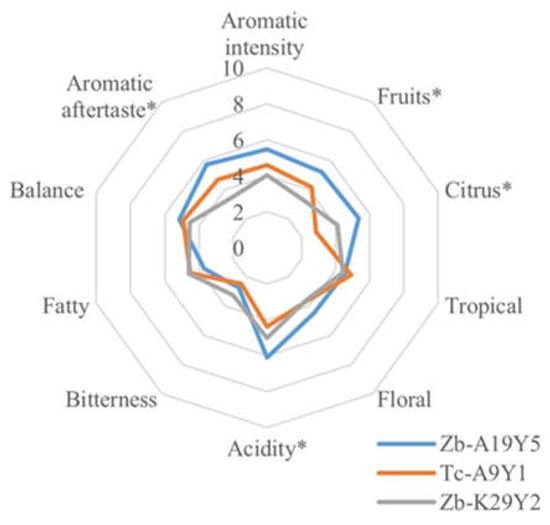
Figure 4.
Spider plot of means of the sensory profile of Assyrtiko wines fermented with different non-Saccharomyces strains at the end of fermentation process. Sample coding: Tc = Trigonopsis californica, Zb = Zygosaccharomyces bailii. (*) indicates the existence of statistically significant differences based on one-way ANOVA, p < 0.05.
In more detail, the statistical analysis (Tuckey’s test) revealed that wines that were inoculated with Zb-A19Y5 and Tc-A9Y1 strains differed the most in terms of fruitiness, citrus, and acidity descriptors. These results are also confirmed by the results of volatile compound analysis. Reversely, wines that were inoculated with Zb-A19Y5 and Zb-K29Y2 strains differed the most in terms of aromatic aftertaste, where the first strain was ranked significantly higher. It is noteworthy that no off-flavor or unpleasant characteristics were noticed by the panelists. Zb-A19Y5 exhibited the highest scores for several attributes, namely, aromatic intensity, fruitiness, citrus, acidity, and aromatic aftertaste. Although the Zb-K29Y2 wines exhibited very high concentrations of isoamyl alcohol (604.33 ± 76.8 mg/L) and differed statistically from the other strains, the fruitiness of these samples was remarkably low. One hypothesis that could explain this phenomenon is the very high concentrations of isoamyl alcohol. It has been underlined, previously, that the presence of isoamyl alcohol alone led to a significant decrease in the “olfactory threshold” of the fruity reconstitution [58]. Motivated by climate change, as well as health issues, significant research efforts have been directed toward strains of yeasts that are capable of generating wines with lower ethanol levels while maintaining a harmonious sensorial profile [72,73,74,75]. According to our results, the T. californica strain Tc-A9Y1 seems to have promising enological potential, well adapted to the wine’s needs. Moreover, the beneficial attributes of the examined yeasts may be remarkably increased or decreased if they are used as co-cultures with S. cerevisiae strains. Additional complementary experiments, designed to investigate the behavior of these yeasts in a must, while considering the presence of other microorganisms, and explore their interactions with S. cerevisiae, could be a very interesting endeavor. For instance, Z. bailii was successfully proposed as a co-culture with S. cerevisiae to improve the production of ethyl esters, which remarkably contributed to the fruitiness and floral wine aroma [76]. Furthermore, mixed starters of Z. bailii and S. cerevisiae also enhanced the wine taste and body by increasing the production of polysaccharides [37].
4. Conclusions
The current study unravels the enological potential of some rarely isolated non-Saccharomyces species that were previously isolated at the end of alcoholic fermentation of spontaneously fermented wines. Strains of Zygosaccharomyces bailii, Priceomyces carsonii, Trigonopsis californica, and Pichia manshurica were investigated for their fermentation capabilities and their impact on the sensory attributes of Assyrtiko must from Santorini. P. carsonii and P. manshurica revealed abnormal fermentations and exhibited an off-odor character. However, T. californica and the two Z. bailii strains led to dry wines which exhibited an interesting organoleptic profile. The ethanol yields of the two Z. bailii strains (0.45 and 0.44 g/g) were higher than those of T. californica (0.39 g/g). Our results confirm that some non-Saccharomyces yeasts are capable of making a positive contribution to volatile compounds in wine. The wines that were inoculated with Zb-A19Y5 were noted as the most promising regarding the overall enological and organoleptic evaluation. Additionally, the ability of a yeast strain to complete fermentation while the final wines show no off-flavor characteristics and reduce the ethanol concentration is also remarkable. Further investigation of the effect of the proposed strains in different must matrixes and with different inoculation combinations is highly recommended.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/app14041522/s1, Table S1: Retention time and kovats index of the identified volatile aromatic compounds.
Author Contributions
Conceptualization, A.T., A.E. and M.D.; methodology, A.T., A.E., G.N., I.P. and M.D.; software, A.T.; validation, A.T., A.E., G.N., I.P. and M.D.; formal analysis, A.T., A.E. and G.N.; writing—original draft preparation, A.T., A.E., G.N., I.P. and M.D.; writing—review and editing, A.T., A.E., G.L. and M.D.; visualization, A.T., A.E. and M.D.; supervision, A.T., A.E. and M.D.; project administration, M.D.; funding acquisition, I.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been co-financed by the European Union and Greek national funds through the Operational Program Competitiveness, Entrepreneurship, and Innovation, under the call “Greece—Israel Call for Proposals for Joint R&D Projects 2019” (project code: T10ΔΙΣ-00060).
Data Availability Statement
The original contributions presented in the study are included in the article and Supplementary Materials, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nanou, E.; Mavridou, E.; Milienos, F.S.; Papadopoulos, G.; Tempère, S.; Kotseridis, Y. Odor Characterization of White Wines Produced from Indigenous Greek Grape Varieties Using the Frequency of Attribute Citation Method with Trained Assessors. Foods 2020, 9, 1396. [Google Scholar] [CrossRef] [PubMed]
- Tzamourani, A.P.; Taliadouros, V.; Paraskevopoulos, I.; Dimopoulou, M. Developing a novel selection method for alcoholic fermentation starters by exploring wine yeast microbiota from Greece. Front. Microbiol. 2023, 14, 1301325. [Google Scholar] [CrossRef] [PubMed]
- Barber, M.; Sung, B.; Lee, S.; Cheah, I. Exploring the influence of novelty and authenticity in wine consumption: The moderating effect of regionality and price. Int. J. Wine Bus. Res. 2021, 33, 288–311. [Google Scholar] [CrossRef]
- Fleet, G.H. Wine Yeasts for the Future. FEMS Yeast Res. 2008, 7, 979–995. [Google Scholar] [CrossRef] [PubMed]
- Fazio, N.A.; Russo, N.; Foti, P.; Pino, A.; Caggia, C.; Randazzo, C.L. Inside Current Winemaking Challenges: Exploiting the Potential of Conventional and Unconventional Yeasts. Microorganisms 2023, 11, 1338. [Google Scholar] [CrossRef] [PubMed]
- Tempère, S.; Marchal, A.; Barbe, J.C.; Bely, M.; Masneuf-Pomarede, I.; Marullo, P.; Albertin, W. The complexity of wine: Clarifying the role of microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 3995–4007. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef]
- Maicas, S.; Mateo, J.J. The Life of Saccharomyces and Non-Saccharomyces Yeasts in Drinking Wine. Microorganisms 2023, 11, 1178. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef]
- Windholtz, S.; Redon, P.; Lacampagne, S.; Farris, L.; Lytra, G.; Cameleyre, M.; Barbe, J.C.; Coulon, J.; Thibon, J.; Masneuf-Pomarède, I. Non-Saccharomyces yeasts as bioprotection in the composition of red wine and in the reduction of sulfur dioxide. LWT 2021, 149, 111781. [Google Scholar] [CrossRef]
- Zott, K.; Thibon, C.; Bely, M.; Lonvaud-Funel, A.; Dubourdieu, D.; Masneuf-Pomarede, I. The grape must non-Saccharomyces microbial community: Impact on volatile thiol release. Int. J. Food Microbiol. 2011, 151, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Guamis, B. Use of UHPH to Obtain Juices With Better Nutritional Quality and Healthier Wines with Low Levels of SO2. Front. Nutr. 2020, 7, 598286. [Google Scholar] [CrossRef]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulou, M.; Goulioti, E.; Troianou, V.; Toumpeki, C.; Paramithiotis, S.; Gosselin, Y.; Dorignac, E.; Papadopoulos, G.; Kotseridis, Y. Effect of Saccharomyces cerevisiae and Saccharomyces pastorianus Co-Inoculation on Alcoholic Fermentation Behavior and Aromatic Profile of Sauvignon Blanc Wine. Fermentation 2022, 8, 539. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Portugal, C.; Pinto, L.; Ribeiro, M.; Tenorio, C.; Igrejas, G.; Fernanda Ruiz-Larrea, F. Potential spoilage yeasts in winery environments: Characterization and proteomic analysis of Trigonopsis cantarellii. Int. J. Food Microbiol. 2015, 210, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. Changes in sour rotten grape berry microbiota during ripening and wine fermentation. Int. J. Food Microbiol. 2012, 154, 152–161. [Google Scholar] [CrossRef]
- Dias, L.; Dias, S.; Sacho, T.; Steinder, H.; Querol, A.; Malfeito-Ferreira, M.; Loureiro, V. Identification of yeasts isolated from wine-related environments and capable of producing 4-ethylphenol. Food Microbiol. 2003, 20, 567–574. [Google Scholar] [CrossRef]
- Kechagia, D.; Paraskevopoulos, Y.; Symeou, E.; Galiotou-Panayotou, M.; Kotseridis, Y. Influence of prefermentative treatments to the major volatile compounds of Assyrtiko wines. J. Agric. Food Chem. 2008, 56, 4555–4563. [Google Scholar] [CrossRef]
- Gump, B.H.; Zoecklein, B.W.; Fugelsang, K.H. Prediction of prefermentation nutritional status of grape juice: The formol method. In Food Microbiology Protocols. Methods in Biotechnology; Humana Press: Totowa, NJ, USA, 2001; pp. 283–296. [Google Scholar] [CrossRef]
- International Organisation of Vine and Wine. Compendium of International Methods of Wine and Must Analysis; OIV: Paris, France, 2021. [Google Scholar]
- Tsapou, E.A.; Dourtoglou, V.; Dourtoglou, T.; Sinanoglou, V.; Koussissi, E. Volatile Profile in Greek Grape Marc Spirits with HS-SPME-GC-MS and Chemometrics: Evaluation of Terroir Impact. ACS Omega 2023, 8, 42803–42814. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Bauer, F.F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- Courcoux, P.; Qannari, E.M.; Faye, P. Free sorting as a sensory profiling technique for product development. In Rapid Sensory Profiling Techniques and Related Methods: Applications in New Product Development and Consumer Research; Delarue, J., Lawlor, J.B., Rogeaux, M., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2015. [Google Scholar] [CrossRef]
- Garofalo, C.; Tristezza, M.; Grieco, F.; Spano, G.; Capozzi, V. From grape berries to wine: Population dynamics of cultivable yeasts associated to “Nero di Troia” autochthonous grape cultivar. World J. Microbiol. Biotechnol. 2016, 32, 59. [Google Scholar] [CrossRef] [PubMed]
- Di Maro, E.; Ercolini, D.; Coppola, S. Yeast dynamics during spontaneous wine fermentation of the Catalanesca grape. Int. J. Food Microbiol. 2007, 117, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. New anamorphic yeast species: Candida infanticola sp. nov., Candida polysorbophila sp. nov., Candida transvaalensis sp. nov. and Trigonopsis californica sp. nov. Antonie Van Leeuwenhoek 2007, 92, 221–231. [Google Scholar] [CrossRef]
- Daniel, H.M.; Lachance, M.A.; Kurtzman, C.P. On the reclassification of species assigned to Candida and other anamorphic ascomycetous yeast genera based on phylogenetic circumscription. Antonie van Leeuwenhoek 2014, 106, 67–84. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Boekhout, T.; Gojkovic, Z.; Katz, M. Evaluation of non-Saccharomyces yeasts in the fermentation of wine, beer and cider for the development of new beverages. J. Inst. Brew. 2018, 124, 389–402. [Google Scholar] [CrossRef]
- Picariello, L.; Rinaldi, A.; Blaiotta, G.; Moio, L.; Pirozzi, P.; Gambuti, A. Effectiveness of chitosan as an alternative to sulfites in red wine production. Eur. Food Res. Technol. 2020, 246, 1795–1804. [Google Scholar] [CrossRef]
- Visintin, S.; Alessandria, V.; Valente, A.; Dolci, P.; Cocolin, L. Molecular identification and physiological characterization of yeasts, lactic acid bacteria and acetic acid bacteria isolated from heap and box cocoa bean fermentations in West Africa. Int. J. Food Microbiol. 2016, 216, 69–78. [Google Scholar] [CrossRef]
- Saez, J.S.; Lopes, C.A.; Kirs, V.E.; Sangorrín, M. Production of volatile phenols by Pichia manshurica and Pichia membranifaciens isolated from spoiled wines and cellar environment in Patagonia. Food Microbiol. 2011, 28, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Tittarelli, F.; Battistelli, N.; Suzzi, G.; Tofalo, R. Contribution of Pichia manshurica strains to aroma profile of organic wines. Eur. Food Res. Technol. 2020, 246, 1405–1417. [Google Scholar] [CrossRef]
- Tzamourani, A.P.; Kasimati, A.; Karagianni, E.; Manthou, E.; Panagou, E.Z. Exploring microbial communities of Spanish-style green table olives of Conservolea and Halkidiki cultivars during modified atmosphere packaging in multi-layered pouches through culture-dependent techniques and metataxonomic analysis. Food Microbiol. 2022, 107, 104063. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.; Augustyn, O.P.H.; Pretorius, I.S. The role and use of non-saccharomyces yeasts in wine production. South Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Wrent, P.; Rivas, E.M.; Peinado, J.M.; de Silóniz, M.I. Strain typing of Zygosaccharomyces yeast species using a single molecular method based on polymorphism of the intergenic spacer region (IGS). Int. J. Food Microbiol. 2010, 142, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Innocente, G.; Gatto, V.; Celebrin, A.; Polo, M.; Felis, G.E.; Torriani, S. Exploring the diversity of a collection of native non-Saccharomyces yeasts to develop co-starter cultures for winemaking. Food Res. Int. 2019, 122, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New insights on the use of wine yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Díaz, M.; Arroyo, T. Biotechnological potential of non-Saccharomyces yeasts isolated during spontaneous fermentations of Malvar (Vitis vinifera cv. L.). Eur. Food Res. Technol. 2013, 236, 193–207. [Google Scholar] [CrossRef]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef]
- Sidira, M.; Galanis, A.; Nikolaou, A.; Kanellaki, M.; Kourkoutas, Y. Evaluation of Lactobacillus casei ATCC 393 protective effect against spoilage of probiotic dry-fermented sausages. Food Control. 2014, 42, 315–320. [Google Scholar] [CrossRef]
- Gamero, A.; Quintilla, R.; Groenewald, M.; Alkema, W.; Boekhout, T.; Hazelwood, L. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiol. 2016, 60, 147–159. [Google Scholar] [CrossRef]
- Solieri, L. The revenge of Zygosaccharomyces yeasts in food biotechnology and applied microbiology. World J. Microbiol. Biotechnol. 2021, 37, 96. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces Yeasts nitrogen source preferences: Impact on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [PubMed]
- Albertin, W.; Zimmer, A.; Miot-Sertier, C.; Bernard, M.; Coulon, J.; Moine, V.; Colonna-Ceccaldi, B.; Bely, M.; Marullo, P.; Masneuf-Pomarede, I. Combined effect of the Saccharomyces cerevisiae lag phase and the non-Saccharomyces consortium to enhance wine fruitiness and complexity. Appl. Microbiol. Biotechnol. 2017, 101, 7603–7620. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, M.; De Vero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative aptitude of non-saccharomyces wine yeast for reduction in the ethanol content in wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Pina, C.; Gonçalves, P.; Prista, C.; Loureiro-Dias, M.C. Ffz1, a new transporter specific for fructose from Zygosaccharomyces bailii. Microbiology 2004, 150, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, J.M.; Childs, B.C.; Edwards, C.G. Evaluation of Zygosaccharomyces bailii to metabolize residual sugar present in partially-fermented red wines. Fermentation 2015, 1, 3–12. [Google Scholar] [CrossRef]
- Basa, K.; Papanikolaou, S.; Dimopoulou, M.; Terpou, A.; Kallithraka, S.; Nychas, G.J. Trials of Commercial-and Wild-Type Saccharomyces Cerevisiae Strains under Aerobic and Microaerophilic/Anaerobic Conditions: Ethanol Production and Must Fermentation from Grapes of Santorini (Greece) Native Varieties. Fermentation 2022, 8, 249. [Google Scholar] [CrossRef]
- Christofi, S.; Papanikolaou, S.; Dimopoulou, M.; Terpou, A.; Cioroiu, I.B.; Cotea, V.; Kallithraka, S. Effect of Yeast Assimilable Nitrogen Content on Fermentation Kinetics, Wine Chemical Composition and Sensory Character in the Production of Assyrtiko Wines. Appl. Sci. 2022, 12, 1405. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Vilela, A. Biological Demalication and deacetification of musts and wines: Can wine yeasts make the wine taste better? Fermentation 2017, 3, 51. [Google Scholar] [CrossRef]
- Sgouros, G.; Mallouchos, A.; Dourou, D.; Banilas, G.; Chalvantzi, I.; Kourkoutas, Y.; Nisiotou, A. Torulaspora delbrueckii May Help Manage Total and Volatile Acidity of Santorini-Assyrtiko Wine in View of Global Warming. Foods 2023, 12, 191. [Google Scholar] [CrossRef]
- Cameleyre, M.; Lytra, G.; Tempere, S.; Barbe, J.C. Olfactory Impact of Higher Alcohols on Red Wine Fruity Ester Aroma Expression in Model Solution. J. Agric. Food Chem. 2015, 63, 9777–9788. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B.; Lonvaud-Funel, A. Handbook of Enology—The Chemistry of Wine: Stabilisationand Treatments, 6th ed.; Dunod: Paris, France, 2012; Volume 1. [Google Scholar]
- Czerny, M.; Christlbauer, M.; Christlbauer, M.; Fischer, A.; Granvogl, M.; Hammer, M.; Hartl, C.; Hernandez, N.M.; Schieberle, P. Re-investigation on odour thresholds of key food aroma compounds and development of an aroma language based on odour qualities of defined aqueous odorant solutions. Eur. Food Res. Technol. 2008, 228, 265–273. [Google Scholar] [CrossRef]
- Etievant, P.X. Wine. In Volatile Compounds in Foods and Beverages; Maarse, H., Ed.; CRC Press: New York, NY, USA, 1991; pp. 483–546. [Google Scholar]
- González, B.; Vázquez, J.; Morcillo-Parra, M.Á.; Mas, A.; Torija, M.J.; Beltran, G. The production of aromatic alcohols in non-Saccharomyces wine yeast is modulated by nutrient availability. Food Microbiol. 2018, 74, 64–74. [Google Scholar] [CrossRef]
- Wu, Q.; Chen, L.; Xu, Y. Yeast community associated with the solid state fermentation of traditional Chinese Maotai-flavor liquor. Int. J. Food Microbiol. 2013, 166, 323–330. [Google Scholar] [CrossRef]
- Vilanova, M.; Escudero, A.; Graña, M.; Cacho, J. Volatile composition and sensory properties of North West Spain white wines. Food Res. Int. 2013, 54, 562–568. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57 (Suppl. A), 131–175. [Google Scholar] [CrossRef]
- Varela, C.; Torrea, D.; Schmidt, S.A.; Ancin-Azpilicueta, C.; Henschke, P.A. Effect of oxygen and lipid supplementation on the volatile composition of chemically defined medium and Chardonnay wine fermented with Saccharomyces cerevisiae. Food Chem. 2012, 135, 2863–2871. [Google Scholar] [CrossRef]
- Duan, W.P.; Zhu, B.Q.; Song, R.R.; Zhang, B.; Lan, Y.-B.; Zhu, X.; Duan, C.Q.; Han, S.Y. Volatile composition and aromatic attributes of wine made with Vitis vinifera cv. Cabernet Sauvignon grapes in the Xinjiang region of China: Effect of different commercial yeasts. Int. J. Food Prop. 2018, 21, 1423–1441. [Google Scholar] [CrossRef]
- Lytra, G.; Tempere, S.; Le Floch, A.; De Revel, G.; Barbe, J. C Study of sensory interactions among red wine fruity esters in a model solution. J. Agric. Food Chem. 2013, 61, 8504–8513. [Google Scholar] [CrossRef] [PubMed]
- Antalick, G.; Perello, M.C.; de Revel, G. Esters in wines: New insight through the establishment of a database of French wines. Am. J. Enol. Vitic. 2014, 65, 293–304. [Google Scholar] [CrossRef]
- Escudero, A.; Campo, E.; Fariña, L.; Cacho, J.; Ferreira, V. Analytical characterization of the aroma of five premium red wines. Insights into the role of odor families and the concept of fruitiness of wines. J. Agric. Food Chem. 2007, 55, 4501–4510. [Google Scholar] [CrossRef] [PubMed]
- Pineau, B.; Barbe, J.C.; Van Leeuwen, C.; Dubourdieu, D. Examples of perceptive interactions involved in specific “Red-” and “Black-berry” aromas in red wines. J. Agric. Food Chem. 2009, 57, 3702–3708. [Google Scholar] [CrossRef] [PubMed]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.C.; Bely, M. Increase of fruity aroma during mixed wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef]
- Kutyna, D.R.; Varela, C.; Henschke, P.A.; Chambers, P.J.; Stanley, G.A. Microbiological approaches to lowering ethanol concentration in wine. Trends Food Sci. Technol. 2010, 21, 293–302. [Google Scholar] [CrossRef]
- Varela, C.; Dry, P.R.; Kutyna, D.R.; Francis, I.L.; Henschke, P.A.; Curtin, C.D.; Chambers, P.J. Strategies for reducing alcohol concentration in wine. Aust. J. Grape Wine Res. 2015, 21, 670–679. [Google Scholar] [CrossRef]
- Varela, C.; Kutyna, D.R.; Solomon, M.R.; Black, C.A.; Borneman, A.; Henschke, P.A.; Pretorius, I.S.; Chambers, P.J. Evaluation of gene modification strategies for the development of low-alcohol wine yeasts. Appl. Environ. Microbiol. 2012, 78, 6068–6077. [Google Scholar] [CrossRef]
- Garavaglia, J.; Schneider, R.d.C.d.S.; Camargo Mendes, S.D.; Welke, J.E.; Zini, C.A.; Caramão, E.B.; Valente, P. Evaluation of Zygosaccharomyces bailii BCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbiol. Res. 2015, 173, 59–65. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).