
The Regional EEG Pattern of the Sleep Onset Process in Older Adults

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Polysomnographic Recordings
2.4. Data Analyses
2.4.1. PSG Measures
2.4.2. Quantitative Analyses of the EEG Signals
2.4.3. Single Hz EEG Topography
2.4.4. Time Course of the EEG Frequency Bands
2.4.5. Comparison between Young Adults and Older Subjects
3. Results
3.1. PSG Measures
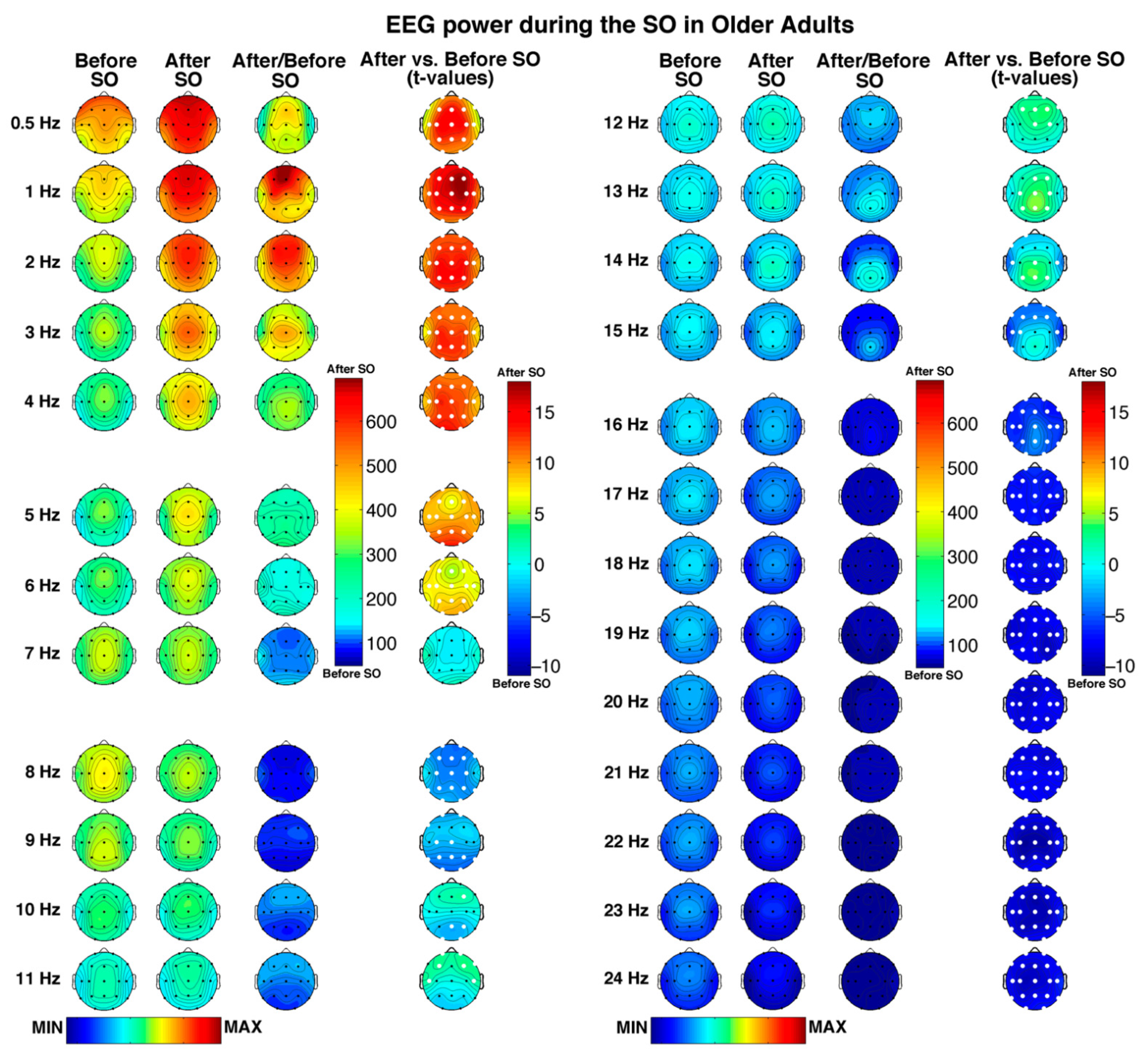
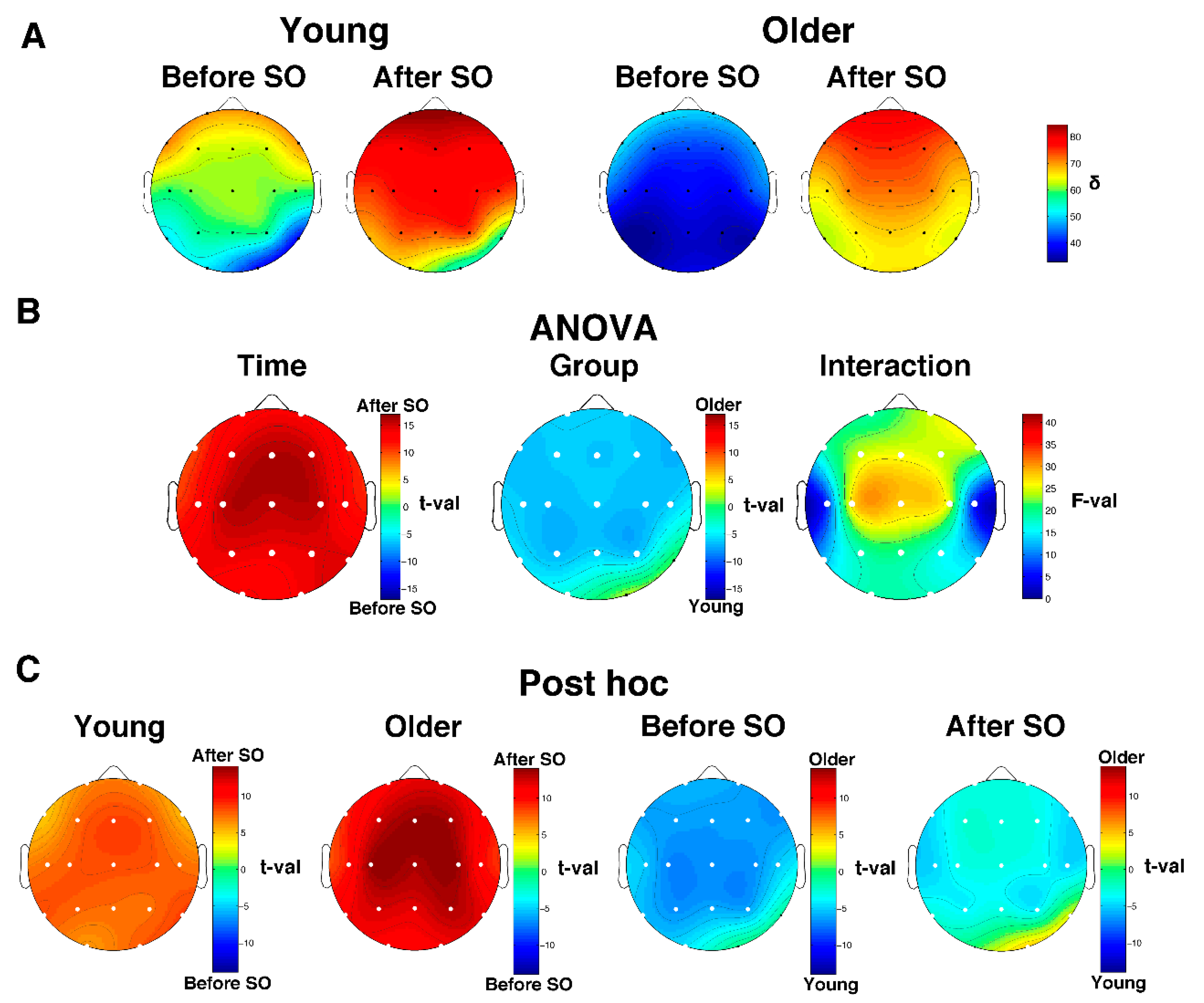
3.2. Scalp Topography of the Single Hz EEG
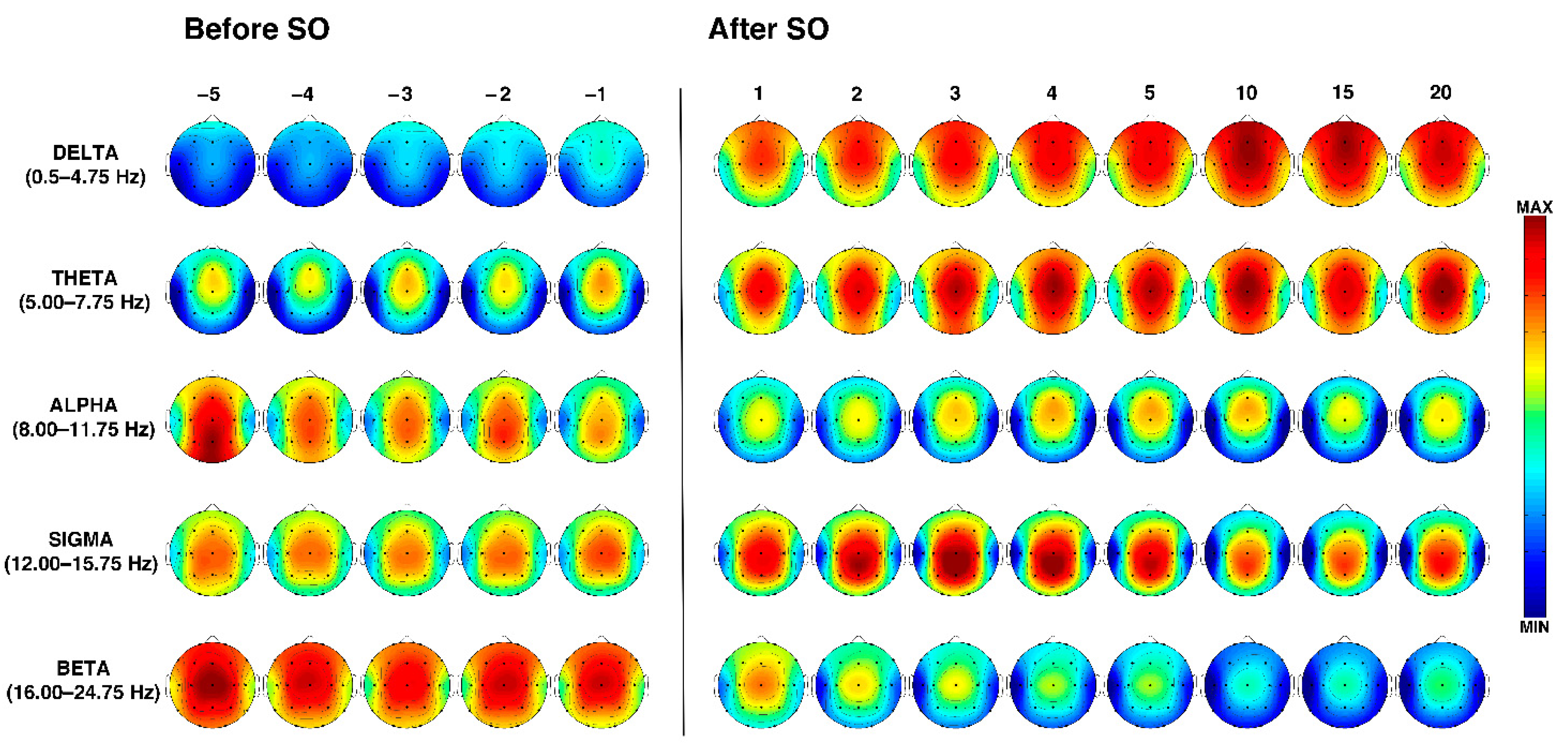
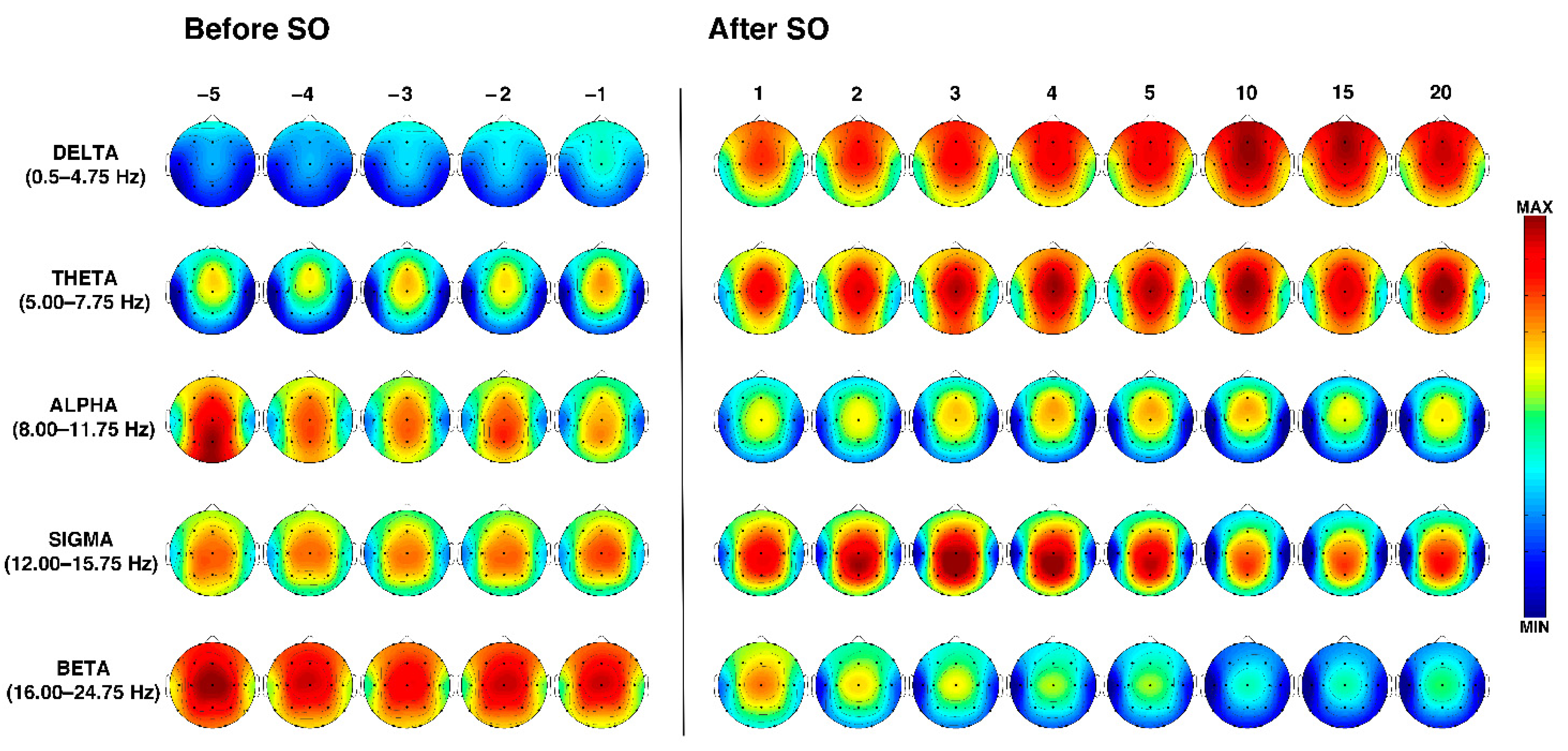
3.3. Time Course of the EEG Frequency Bands
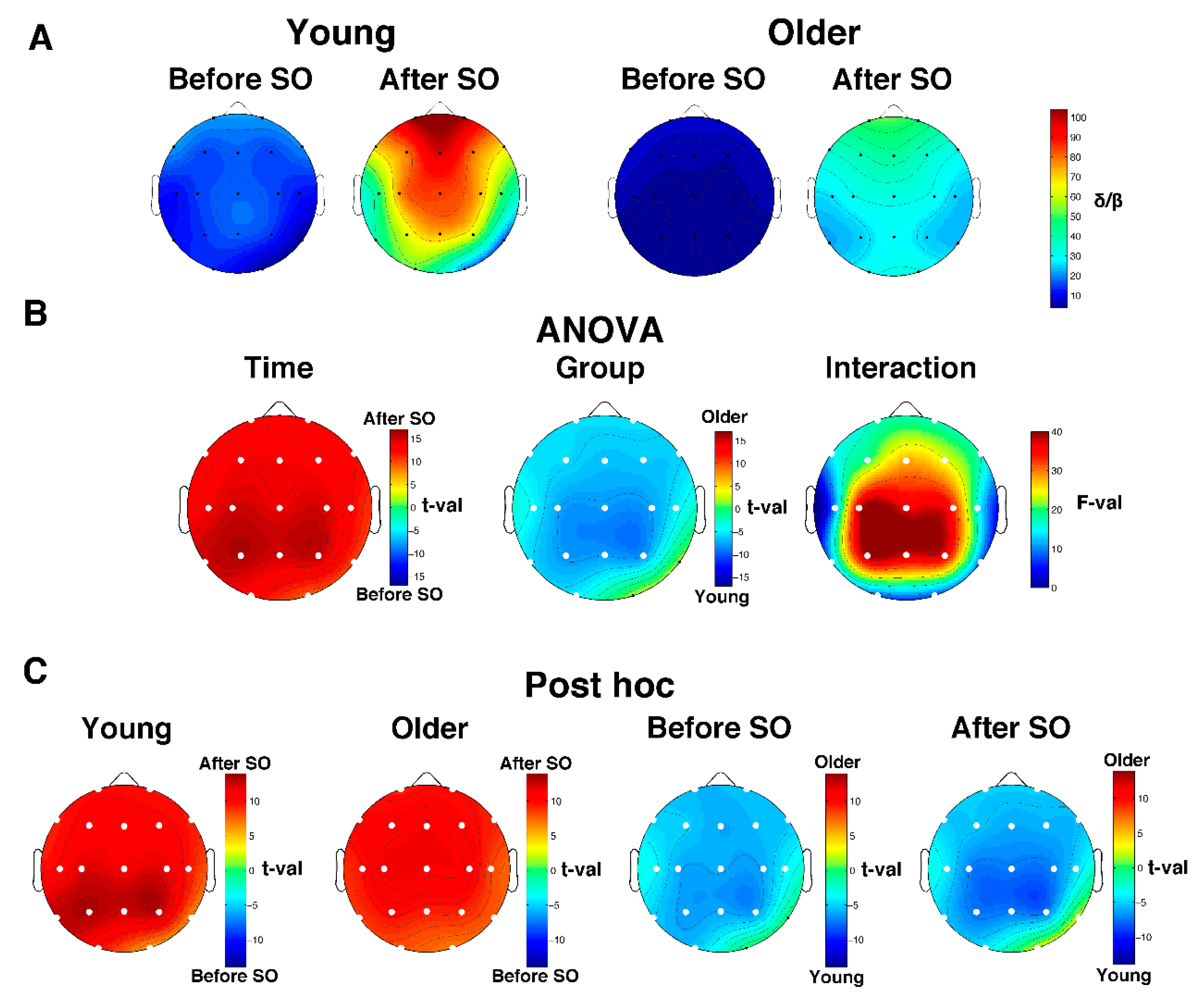
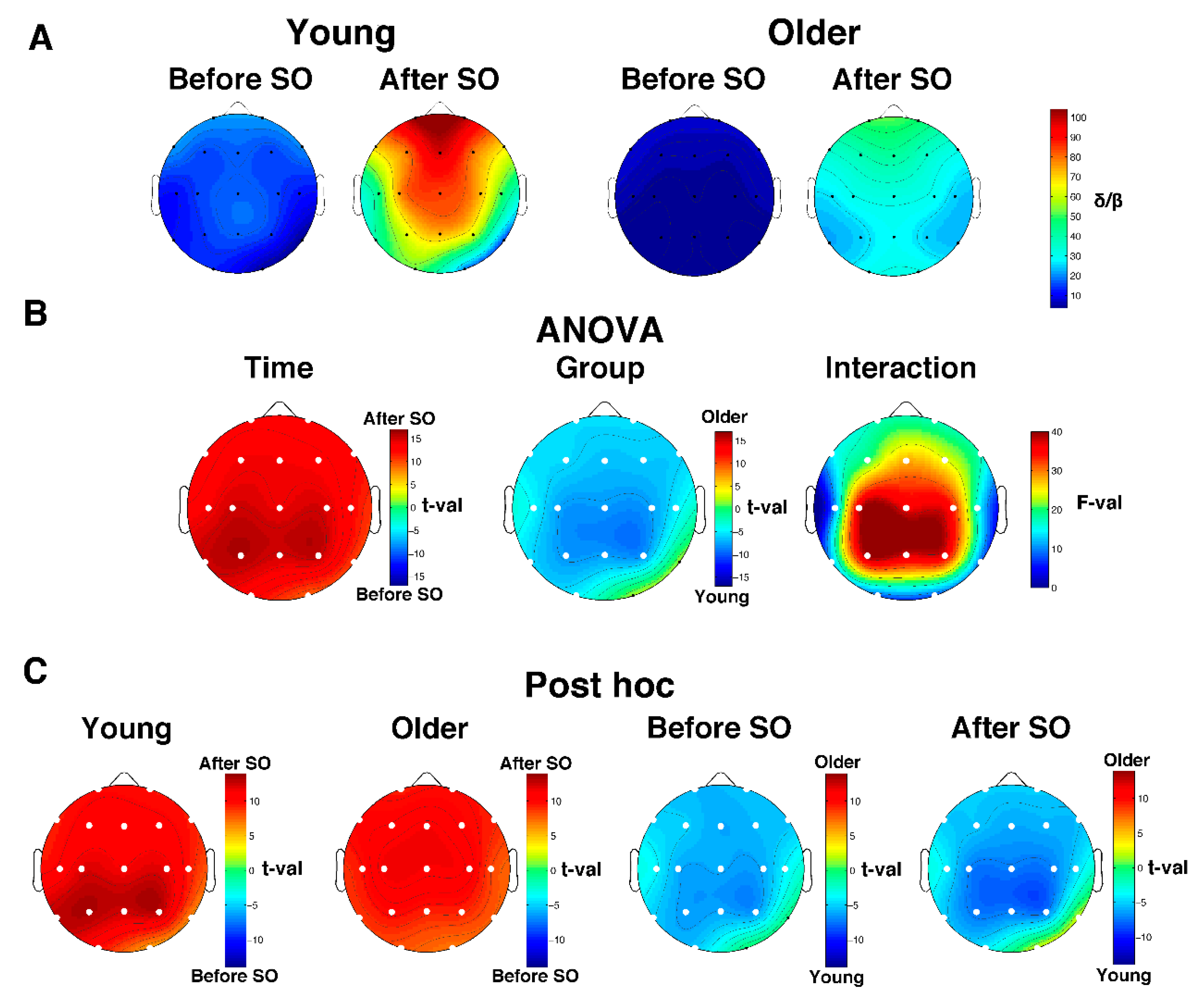
3.4. Age-Dependent Modifications in SWA and Delta/Beta Ratio
3.4.1. Relative Delta Power
3.4.2. Delta/Beta Ratio
4. Discussion
4.1. SWA and Delta/Beta Ratio
4.2. Theta Activity
4.3. Alpha Activity
4.4. Sigma Activity
4.5. Beta Activity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferrara, M.; De Gennaro, L. Going local: Insights from EEG and stereo-EEG studies of the human sleep-wake cycle. Curr. Top. Med. Chem. 2011, 11, 2423–2437. [Google Scholar] [CrossRef]
- Siclari, F.; Tononi, G. Local aspects of sleep and wakefulness. Curr. Opin. Neurobiol. 2017, 44, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.M.; Rector, D.M.; Roy, S.; Van Dongen, H.P.A.; Belenky, G.; Panksepp, J. Sleep as a fundamental property of neuronal assemblies. Nat. Rev. Neurosci. 2008, 9, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.M.; Nguyen, J.T.; Dykstra-Aiello, C.J.; Taishi, P. Local sleep. Sleep Med. Rev. 2019, 43, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.; Winer, J.R.; Walker, M.P. Sleep and Human Aging. Neuron 2017, 94, 19–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprecher, K.E.; Riedner, B.A.; Smith, R.F.; Tononi, G.; Davidson, R.; Benca, R.M. High Resolution Topography of Age-Related Changes in Non-Rapid Eye Movement Sleep Electroencephalography. PLoS ONE 2016, 11, e0149770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dijk, D.J.; Beersma, D.G.; van den Hoofdakker, R.H. All night spectral analysis of EEG sleep in young adult and middle-aged male subjects. Neurobiol. Aging 1989, 10, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Landolt, H.-P.; Dijk, D.-J.; Achermann, P.; Borbély, A.A. Effect of age on the sleep EEG: Slow-wave activity and spindle frequency activity in young and middle-aged men. Brain Res. 1996, 738, 205–212. [Google Scholar] [CrossRef]
- Carrier, J.; Land, S.; Buysse, D.J.; Kupfer, D.J.; Monk, T.H. The effects of age and gender on sleep EEG power spectral density in the middle years of life (ages 20–60 years old). Psychophysiology 2001, 38, 232–242. [Google Scholar] [CrossRef]
- Mander, B.A.; Rao, V.; Lu, B.; Saletin, J.; Lindquist, J.R.; Ancoli-Israel, S.; Jagust, W.; Walker, M.P. Prefrontal atrophy, disrupted NREM slow waves and impaired hippocampal-dependent memory in aging. Nat. Neurosci. 2013, 16, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Mander, B.A.; Rao, V.; Lu, B.; Saletin, J.M.; Ancoli-Israel, S.; Jagust, W.J.; Walker, M.P. Impaired Prefrontal Sleep Spindle Regulation of Hippocampal-Dependent Learning in Older Adults. Cereb. Cortex 2013, 24, 3301–3309. [Google Scholar] [CrossRef] [Green Version]
- Mander, B.A.; Zhu, A.H.; Lindquist, J.R.; Villeneuve, S.; Rao, V.; Lu, B.; Saletin, J.M.; Ancoli-Israel, S.; Jagust, W.J.; Walker, M.P. White Matter Structure in Older Adults Moderates the Benefit of Sleep Spindles on Motor Memory Consolidation. J. Neurosci. 2017, 37, 11675–11687. [Google Scholar] [CrossRef] [Green Version]
- Martin, N.; Lafortune, M.; Godbout, J.; Barakat, M.; Robillard, R.; Poirier, G.; Bastien, C.; Carrier, J. Topography of age-related changes in sleep spindles. Neurobiol. Aging 2012, 34, 468–476. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.-P.; Borbély, A.A. Age-dependent changes in sleep EEG topography. Clin. Neurophysiol. 2001, 112, 369–377. [Google Scholar] [CrossRef]
- Münch, M.; Knoblauch, V.; Blatter, K.; Schröder, C.; Schnitzler, C.; Kräuchi, K.; Wirz-Justice, A.; Cajochen, C. The frontal predominance in human EEG delta activity after sleep loss decreases with age. Eur. J. Neurosci. 2004, 20, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.; Duffy, J.F.; Riel, E.; Shanahan, T.L.; Czeisler, C.A. Ageing and the circadian and homeostatic regulation of human sleep during forced desynchrony of rest, melatonin and temperature rhythms. J. Physiol. 1999, 516, 611–627. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, R.F.; Mander, B.A.; Jagust, W.J.; Knight, R.T.; Walker, M.P. Old Brains Come Uncoupled in Sleep: Slow Wave-Spindle Synchrony, Brain Atrophy, and Forgetting. Neuron 2017, 97, 221–230.e4. [Google Scholar] [CrossRef] [Green Version]
- Muehlroth, B.E.; Sander, M.C.; Fandakova, Y.; Grandy, T.H.; Rasch, B.; Shing, Y.L.; Werkle-Bergner, M. Precise Slow Os-cillation–Spindle Coupling Promotes Memory Consolidation in Younger and Older Adults. Sci. Rep. 2019, 9, 1940. [Google Scholar] [CrossRef] [Green Version]
- Scarpelli, S.; D’Atri, A.; Bartolacci, C.; Mangiaruga, A.; Gorgoni, M.; De Gennaro, L. Oscillatory EEG Activity During REM Sleep in Elderly People Predicts Subsequent Dream Recall After Awakenings. Front. Neurol. 2019, 10, 985. [Google Scholar] [CrossRef]
- Ogilvie, R.D. The process of falling asleep. Sleep Med. Rev. 2001, 5, 247–270. [Google Scholar] [CrossRef]
- Merica, H.; Fortune, R.D. State transitions between wake and sleep, and within the ultradian cycle, with focus on the link to neuronal activity. Sleep Med. Rev. 2004, 8, 473–485. [Google Scholar] [CrossRef]
- Gorgoni, M.; D’Atri, A.; Scarpelli, S.; Ferrara, M.; De Gennaro, L. Timing and Topography of Sleep Onset: Asynchronies and Regional Changes of Brain Activity. In Handbook of Behavioral Neuroscience; Elsevier: Amsterdam, The Netherlands, 2019; Volume 30, pp. 19–31. [Google Scholar]
- Gorgoni, M.; D’Atri, A.; Scarpelli, S.; Ferrara, M.; De Gennaro, L. The electroencephalographic features of the sleep onset process and their experimental manipulation with sleep deprivation and transcranial electrical stimulation protocols. Neurosci. Biobehav. Rev. 2020, 114, 25–37. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Ferrara, M.; Bertini, M. The boundary between wakefulness and sleep: Quantitative electroencephalo-graphic changes during the sleep onset period. Neuroscience 2001, 107, 1–11. [Google Scholar] [CrossRef]
- De Gennaro, L.; Ferrara, M.; Curcio, G.; Cristiani, R. Antero-posterior EEG changes during the wakefulness-sleep transition. Clin. Neurophysiol. 2001, 112, 1901–1911. [Google Scholar] [CrossRef]
- Marzano, C.; Moroni, F.; Gorgoni, M.; Nobili, L.; Ferrara, M.; De Gennaro, L. How we fall asleep: Regional and temporal differences in electroencephalographic synchronization at sleep onset. Sleep Med. 2013, 14, 1112–1122. [Google Scholar] [CrossRef]
- Siclari, F.; Bernardi, G.; Riedner, B.A.; Larocque, J.J.; Benca, R.M.; Tononi, G. Two Distinct Synchronization Processes in the Transition to Sleep: A High-Density Electroencephalographic Study. Sleep 2014, 37, 1621–1637. [Google Scholar] [CrossRef]
- Magnin, M.; Rey, M.; Bastuji, H.; Guillemant, P.; Mauguière, F.; Garcia-Larrea, L. Thalamic deactivation at sleep onset precedes that of the cerebral cortex in humans. Proc. Natl. Acad. Sci. USA 2010, 107, 3829–3833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarasso, S.; Proserpio, P.; Pigorini, A.; Moroni, F.; Ferrara, M.; De Gennaro, L.; De Carli, F.; Lo Russo, G.; Massimini, M.; Nobili, L. Hippocampal sleep spindles preceding neocortical sleep onset in humans. NeuroImage 2014, 86, 425–432. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Vecchio, F.; Ferrara, M.; Curcio, G.; Rossini, P.M.; Babiloni, C. Changes in fronto-posterior functional cou-pling at sleep onset in humans. J. Sleep Res. 2004, 13, 209–217. [Google Scholar] [CrossRef]
- Larson-Prior, L.J.; Zempel, J.M.; Nolan, T.S.; Prior, F.W.; Snyder, A.Z.; Raichle, M.E. Cortical network functional connectivity in the descent to sleep. Proc. Natl. Acad. Sci. USA 2009, 106, 4489–4494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchio, F.; Miraglia, F.; Gorgoni, M.; Ferrara, M.; Iberite, F.; Bramanti, P.; De Gennaro, L.; Rossini, P.M. Cortical connectivity modulation during sleep onset: A study via graph theory on EEG data. Hum. Brain Mapp. 2017, 38, 5456–5464. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, A.F.; Achermann, P. Intracortical Causal Information Flow of Oscillatory Activity (Effective Connectivity) at the Sleep Onset Transition. Front. Neurosci. 2018, 12, 912. [Google Scholar] [CrossRef]
- De Gennaro, L.; Vecchio, F.; Ferrara, M.; Curcio, G.; Rossini, P.M.; Babiloni, C. Antero-posterior functional coupling at sleep onset: Changes as a function of increased sleep pressure. Brain Res. Bull. 2005, 65, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, A.F.; Achermann, P. Brain dynamics during the sleep onset transition: An EEG source localization study. Neurobiol. Sleep Circadian Rhythm. 2018, 6, 24–34. [Google Scholar] [CrossRef]
- Gorgoni, M.; Bartolacci, C.; D’Atri, A.; Scarpelli, S.; Marzano, C.; Moroni, F.; Ferrara, M.; De Gennaro, L. The Spatiotemporal Pattern of the Human Electroencephalogram at Sleep Onset after a Period of Prolonged Wakefulness. Front. Neurosci. 2019, 13, 312. [Google Scholar] [CrossRef] [Green Version]
- Miraglia, F.; Tomino, C.; Vecchio, F.; Gorgoni, M.; De Gennaro, L.; Rossini, P.M. The brain network organization during sleep onset after deprivation. Clin. Neurophysiol. 2021, 132, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Spiess, M.; Bernardi, G.; Kurth, S.; Ringli, M.; Wehrle, F.M.; Jenni, O.G.; Huber, R.; Siclari, F. How do children fall asleep? A high-density EEG study of slow waves in the transition from wake to sleep. NeuroImage 2018, 178, 23–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Atri, A.; Scarpelli, S.; Schiappa, C.; Pizza, F.; Vandi, S.; Ferrara, M.; Cipolli, C.; Plazzi, G.; De Gennaro, L. Cortical activation during sleep predicts dream experience in narcolepsy. Ann. Clin. Transl. Neurol. 2019, 6, 445–455. [Google Scholar] [CrossRef]
- Scarpelli, S.; D’Atri, A.; Bartolacci, C.; Gorgoni, M.; Mangiaruga, A.; Ferrara, M.; De Gennaro, L. Dream Recall upon Awakening from Non-Rapid Eye Movement Sleep in Older Adults: Electrophysiological Pattern and Qualitative Features. Brain Sci. 2020, 10, 343. [Google Scholar] [CrossRef]
- Krystal, A.D. Non-REM sleep EEG spectral analysis in insomnia. Psychiatr. Ann. 2008, 38, 615–620. [Google Scholar]
- Maes, J.; Verbraecken, J.; Willemen, M.; De Volder, I.; van Gastel, A.; Michiels, N.; Verbeek, I.; Vandekerckhove, M.; Wuyts, J.; Haex, B.; et al. Sleep misperception, EEG characteristics and Autonomic Nervous System activity in primary insomnia: A retrospective study on polysomnographic data. Int. J. Psychophysiol. 2014, 91, 163–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechtschaffen, A.; Kales, A. A Manual of Standardized Terminology, Techniques and Scoring System for Sleep Stages of Human Subjects; UCLA Brain Information Service: Los Angeles, CA, USA, 1968. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Gorgoni, M.; Ferrara, M.; D’Atri, A.; Lauri, G.; Scarpelli, S.; Truglia, I.; De Gennaro, L. EEG topography during sleep inertia upon awakening after a period of increased homeostatic sleep pressure. Sleep Med. 2015, 16, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Gorgoni, M.; Lauri, G.; Truglia, I.; Cordone, S.; Sarasso, S.; Scarpelli, S.; Mangiaruga, A.; D’Atri, A.; Tempesta, D.; Ferrara, M.; et al. Parietal Fast Sleep Spindle Density Decrease in Alzheimer’s Disease and Amnesic Mild Cognitive Impairment. Neural Plast. 2016, 2016, 8376108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gennaro, L.; Gorgoni, M.; Reda, F.; Lauri, G.; Truglia, I.; Cordone, S.; Scarpelli, S.; Mangiaruga, A.; D’Atri, A.; Lacidogna, G.; et al. The Fall of Sleep K-Complex in Alzheimer Disease. Sci. Rep. 2017, 7, 39688. [Google Scholar] [CrossRef] [Green Version]
- Reda, F.; Gorgoni, M.; Lauri, G.; Truglia, I.; Cordone, S.; Scarpelli, S.; Mangiaruga, A.; D’Atri, A.; Ferrara, M.; Lacidogna, G.; et al. In Search of Sleep Biomarkers of Alzheimer’s Disease: K-Complexes Do Not Discriminate between Patients with Mild Cognitive Impairment and Healthy Controls. Brain Sci. 2017, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- D’Atri, A.; Scarpelli, S.; Gorgoni, M.; Truglia, I.; Lauri, G.; Cordone, S.; Ferrara, M.; Marra, C.; Rossini, P.M.; De Gennaro, L. EEG alterations during wake and sleep in mild cognitive impairment and Alzheimer’s disease. iScience 2021, 24, 102386. [Google Scholar] [CrossRef]
- Tanaka, H.; Hayashi, M.; Hori, T. Topographical characteristics of slow wave activities during the transition from wake-fulness to sleep. Clin. Neurophysiol. 2000, 111, 417–427. [Google Scholar] [CrossRef]
- Werth, E.; Achermann, P.; Borbély, A. Fronto-occipital EEG power gradients in human sleep. J. Sleep Res. 1997, 6, 102–112. [Google Scholar] [CrossRef]
- Carrier, J.; Viens, I.; Poirier, G.; Robillard, R.; Lafortune, M.; Vandewalle, G.; Martin, N.; Barakat, M.; Paquet, J.; Filipini, D. Sleep slow wave changes during the middle years of life. Eur. J. Neurosci. 2011, 33, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Dubé, J.; LaFortune, M.; Bedetti, C.; Bouchard, M.; Gagnon, J.F.; Doyon, J.; Evans, A.C.; Lina, J.-M.; Carrier, J. Cortical Thinning Explains Changes in Sleep Slow Waves during Adulthood. J. Neurosci. 2015, 35, 7795–7807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, J.; Broughton, R. Quantitative topographic EEG mapping during drowsiness and sleep onset. In Sleep Onset: Normal and Abnormal Processes; Ogilvie, R.D., Harsh, J.R., Eds.; American Psychological Association: Washington, DC, USA, 1994; pp. 219–235. [Google Scholar]
- Tanaka, H.; Hayashi, M.; Hori, T. Topographical Characteristics and Principal Component Structure of the Hypnagogic EEG. Sleep 1997, 20, 523–534. [Google Scholar] [CrossRef] [Green Version]
- Pivik, R.T.; Harman, K. A reconceptualization of EEG alpha activity as an index of arousal during sleep: All alpha activity is not equal. J. Sleep Res. 1995, 4, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Gaal, Z.A.; Boha, R.; Stam, C.J.; Molnar, M. Age-dependent features of EEG-reactivity—Spectral, complexity, and network characteristics. Neurosci. Lett. 2010, 479, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Klass, D.W.; Brenner, R.P. Electroencephalography of the Elderly. J. Clin. Neurophysiol. 1995, 12, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Klimesch, W. EEG alpha and theta oscillations reflect cognitive and memory performance: A review and analysis. Brain Res. Rev. 1999, 29, 169–195. [Google Scholar] [CrossRef]
- Babiloni, C.; Binetti, G.; Cassarino, A.; Dal Forno, G.; Del Percio, C.; Ferreri, F.; Ferri, R.; Frisoni, G.B.; Galderisi, S.; Hirata, K.; et al. Sources of cortical rhythms in adults during physiological aging: A multicentric EEG study. Hum. Brain Mapp. 2005, 27, 162–172. [Google Scholar] [CrossRef] [Green Version]
- Uhlhaas, P.J.; Singer, W. Neural Synchrony in Brain Disorders: Relevance for Cognitive Dysfunctions and Pathophysiology. Neuron 2006, 52, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Finnigan, S.; Robertson, I.H. Resting EEG theta power correlates with cognitive performance in healthy older adults: Resting theta EEG correlates with cognitive aging. Psychophysiology 2011, 48, 10831087. [Google Scholar] [CrossRef]
- ElShafei, H.A.; Fornoni, L.; Masson, R.; Bertrand, O.; Bidet-Caulet, A. Age-related modulations of alpha and gamma brain activities underlying anticipation and distraction. PLoS ONE 2020, 15, e0229334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niedermeyer, E. Alpha rhythms as physiological and abnormal phenomena. Int. J. Psychophysiol. 1997, 26, 31–49. [Google Scholar] [CrossRef]
- Başar, E. A review of alpha activity in integrative brain function: Fundamental physiology, sensory coding, cognition and pathology. Int. J. Psychophysiol. 2012, 86, 1–24. [Google Scholar] [CrossRef] [PubMed]
- De Gennaro, L.; Ferrara, M. Sleep spindles: An overview. Sleep Med. Rev. 2003, 7, 423–440. [Google Scholar] [CrossRef]
- Celesia, G.G. EEG and Event-Related Potentials in Aging and Dementia. J. Clin. Neurophysiol. 1986, 3, 99–111. [Google Scholar] [CrossRef]
- Vlahou, E.L.; Thurm, F.; Kolassa, I.-T.; Schlee, W. Resting-state slow wave power, healthy aging and cognitive performance. Sci. Rep. 2015, 4, 5101. [Google Scholar] [CrossRef] [PubMed]
- Merica, H.; Fortune, R.D.; Gaillard, J.M. Hemispheric temporal organization during the onset of sleep in normal subjects. In Phasic Events and Dynamic Organization of Sleep; Terzano, M.G., Halász, P.L., Declerck, A.C., Eds.; Raven Press: New York, NY, USA, 1991; pp. 73–83. [Google Scholar]
- Merica, H.; Gaillard, J.-M. The EEG of the sleep onset period in insomnia: A discriminant analysis. Physiol. Behav. 1992, 52, 199–204. [Google Scholar] [CrossRef]
- Başar-Eroglu, C.; Strüber, D.; Schürmann, M.; Stadler, M.; Başar, E. Gamma-band responses in the brain: A short review of psychophysiological correlates and functional significance. Int. J. Psychophysiol. 1996, 24, 101–112. [Google Scholar] [CrossRef]
- Kilavik, B.E.; Zaepffel, M.; Brovelli, A.; MacKay, W.A.; Riehle, A. The ups and downs of β oscillations in sensorimotor cortex. Exp. Neurol. 2013, 245, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Merker, B. Cortical gamma oscillations: The functional key is activation, not cognition. Neurosci. Biobehav. Rev. 2013, 37, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Neuper, C.; Pfurtscheller, G. Event-related dynamics of cortical rhythms: Frequency specific features and functional correlates. Int. J. Psychophysiol. 2001, 43, 41–58. [Google Scholar] [CrossRef]
- Ishii, R.; Canuet, L.; Aoki, Y.; Hata, M.; Iwase, M.; Ikeda, S.; Nishida, K.; Ikeda, M. Healthy and Pathological Brain Aging: From the Perspective of Oscillations, Functional Connectivity, and Signal Complexity. Neuropsychobiology 2017, 75, 151–161. [Google Scholar] [CrossRef] [PubMed]
- D’Atri, A.; De Simoni, E.; Gorgoni, M.; Ferrara, M.; Ferlazzo, F.; Rossini, P.; De Gennaro, L. Electrical stimulation of the frontal cortex enhances slow-frequency EEG activity and sleepiness. Neuroscience 2016, 324, 119–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Atri, A.; Romano, C.; Gorgoni, M.; Scarpelli, S.; Alfonsi, V.; Ferrara, M.; Ferlazzo, F.; Rossini, P.M.; De Gennaro, L. Bilateral 5 Hz transcranial alternating current stimulation on fronto-temporal areas modulates resting-state EEG. Sci. Rep. 2017, 7, 15672. [Google Scholar] [CrossRef] [PubMed]
- D’Atri, A.; Scarpelli, S.; Gorgoni, M.; Alfonsi, V.; Annarumma, L.; Giannini, A.M.; Ferrara, M.; Ferlazzo, F.; Rossini, P.M.; De Gennaro, L. Bilateral Theta Transcranial Alternating Current Stimulation (tACS) Modulates EEG Activity: When tACS Works Awake It Also Works Asleep. Nat. Sci. Sleep 2019, 11, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Cellini, N.; Mednick, S.C. Stimulating the sleeping brain: Current approaches to modulating memory-related sleep physiology. J. Neurosci. Methods 2018, 316, 125–136. [Google Scholar] [CrossRef]
- Fehér, K.D.; Wunderlin, M.; Maier, J.G.; Hertenstein, E.; Schneider, C.L.; Mikutta, C.; Züst, M.A.; Klöppel, S.; Nissen, C. Shaping the slow waves of sleep: A systematic and integrative review of sleep slow wave modulation in humans using non-invasive brain stimulation. Sleep Med. Rev. 2021, 58, 101438. [Google Scholar] [CrossRef]
- Salfi, F.; D’Atri, A.; Tempesta, D.; De Gennaro, L.; Ferrara, M. Boosting Slow Oscillations during Sleep to Improve Memory Function in Elderly People: A Review of the Literature. Brain Sci. 2020, 10, 300. [Google Scholar] [CrossRef]
- Cordone, S.; Scarpelli, S.; Alfonsi, V.; De Gennaro, L.; Gorgoni, M. Sleep-Based Interventions in Alzheimer’s Disease: Promising Approaches from Prevention to Treatment along the Disease Trajectory. Pharmaceuticals 2021, 14, 383. [Google Scholar] [CrossRef]
- Curcio, G.; Ferrara, M.; Piergianni, A.; Fratello, F.; De Gennaro, L. Paradoxes of the first-night effect: A quantitative analysis of antero-posterior EEG topography. Clin. Neurophysiol. 2004, 115, 1178–1188. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Older Adults Mean ± SE | Young Adults Mean ± SE | T | p | |
|---|---|---|---|---|
| Stage 1 latency (min) | 14.9 ± 2.7 | 6.6 ± 0.9 | 3.9 | * 0.003 |
| Stage 2 latency (min) | 15.7 ± 2.6 | 11.1 ± 1.8 | 1.4 | 0.20 |
| Stage 1 (%) | 5.0 ± 0.5 | 6.3 ± 0.5 | −1.9 | 0.06 |
| Stage 2 (%) | 77.1 ± 0.9 | 59.3 ± 1.1 | 12.6 | * <0.00001 |
| SWS (%) | 0.8 ± 0.2 | 10.3 ± 1.0 | −8.8 | * <0.00001 |
| REM (%) | 17.1 ± 0.8 | 24.1 ± 0.8 | −6.2 | * <0.00001 |
| WASO (min) | 72.2 ± 7.2 | 26.1 ± 3.0 | 6.1 | * <0.00001 |
| Awakenings (#) | 18.9 ± 1.4 | 28.6 ± 1.7 | −4.3 | * 0.00005 |
| Arousals (#) | 19.9 ± 1.8 | 35.4 ± 2.8 | −4.5 | * 0.00002 |
| TST (min) | 298.2 ± 9.1 | 441.4 ± 6.1 | −13.3 | * <0.00001 |
| TBT (min) | 385.7 ± 7.4 | 484.8 ± 10.1 | −7.8 | * <0.00001 |
| SEI % (TST/TBT) | 77.5 ± 2.0 | 91.7 ± 1.1 | −6.4 | * <0.00001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorgoni, M.; Scarpelli, S.; Annarumma, L.; D’Atri, A.; Alfonsi, V.; Ferrara, M.; De Gennaro, L. The Regional EEG Pattern of the Sleep Onset Process in Older Adults. Brain Sci. 2021, 11, 1261. https://doi.org/10.3390/brainsci11101261
Gorgoni M, Scarpelli S, Annarumma L, D’Atri A, Alfonsi V, Ferrara M, De Gennaro L. The Regional EEG Pattern of the Sleep Onset Process in Older Adults. Brain Sciences. 2021; 11(10):1261. https://doi.org/10.3390/brainsci11101261
Chicago/Turabian StyleGorgoni, Maurizio, Serena Scarpelli, Ludovica Annarumma, Aurora D’Atri, Valentina Alfonsi, Michele Ferrara, and Luigi De Gennaro. 2021. "The Regional EEG Pattern of the Sleep Onset Process in Older Adults" Brain Sciences 11, no. 10: 1261. https://doi.org/10.3390/brainsci11101261
APA StyleGorgoni, M., Scarpelli, S., Annarumma, L., D’Atri, A., Alfonsi, V., Ferrara, M., & De Gennaro, L. (2021). The Regional EEG Pattern of the Sleep Onset Process in Older Adults. Brain Sciences, 11(10), 1261. https://doi.org/10.3390/brainsci11101261