
Structural Connectivity Alterations in Operculo-Insular Epilepsy

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Standard Protocol Approvals and Patient Consents
2.3. Image Acquisition
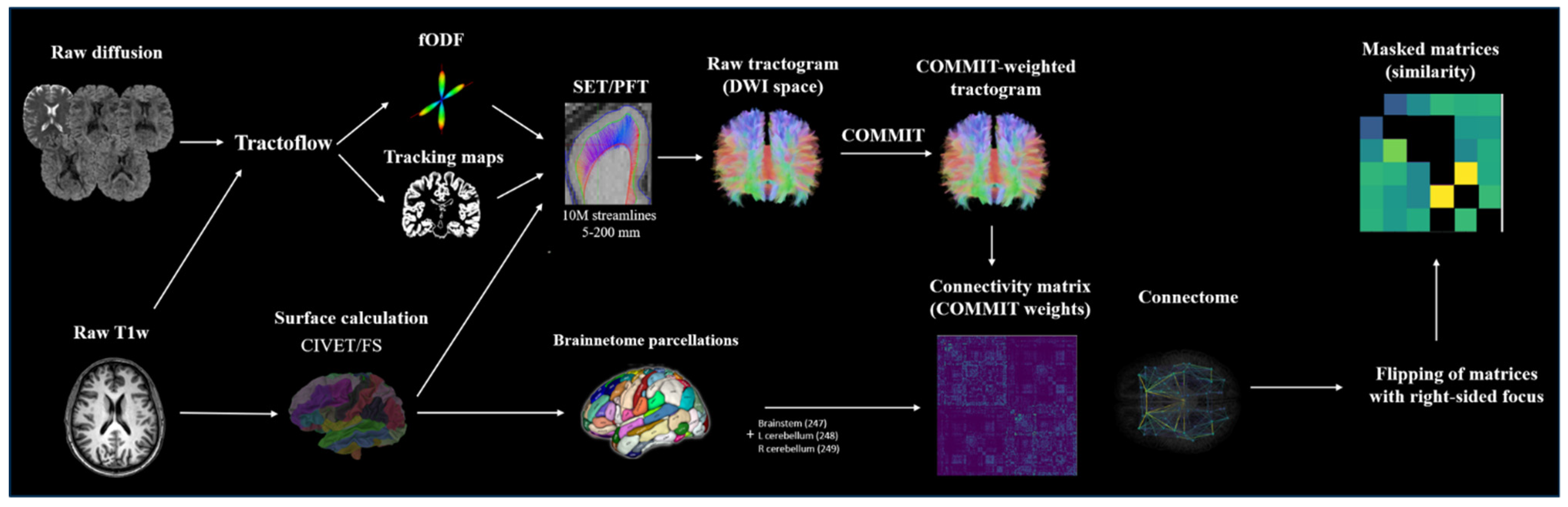
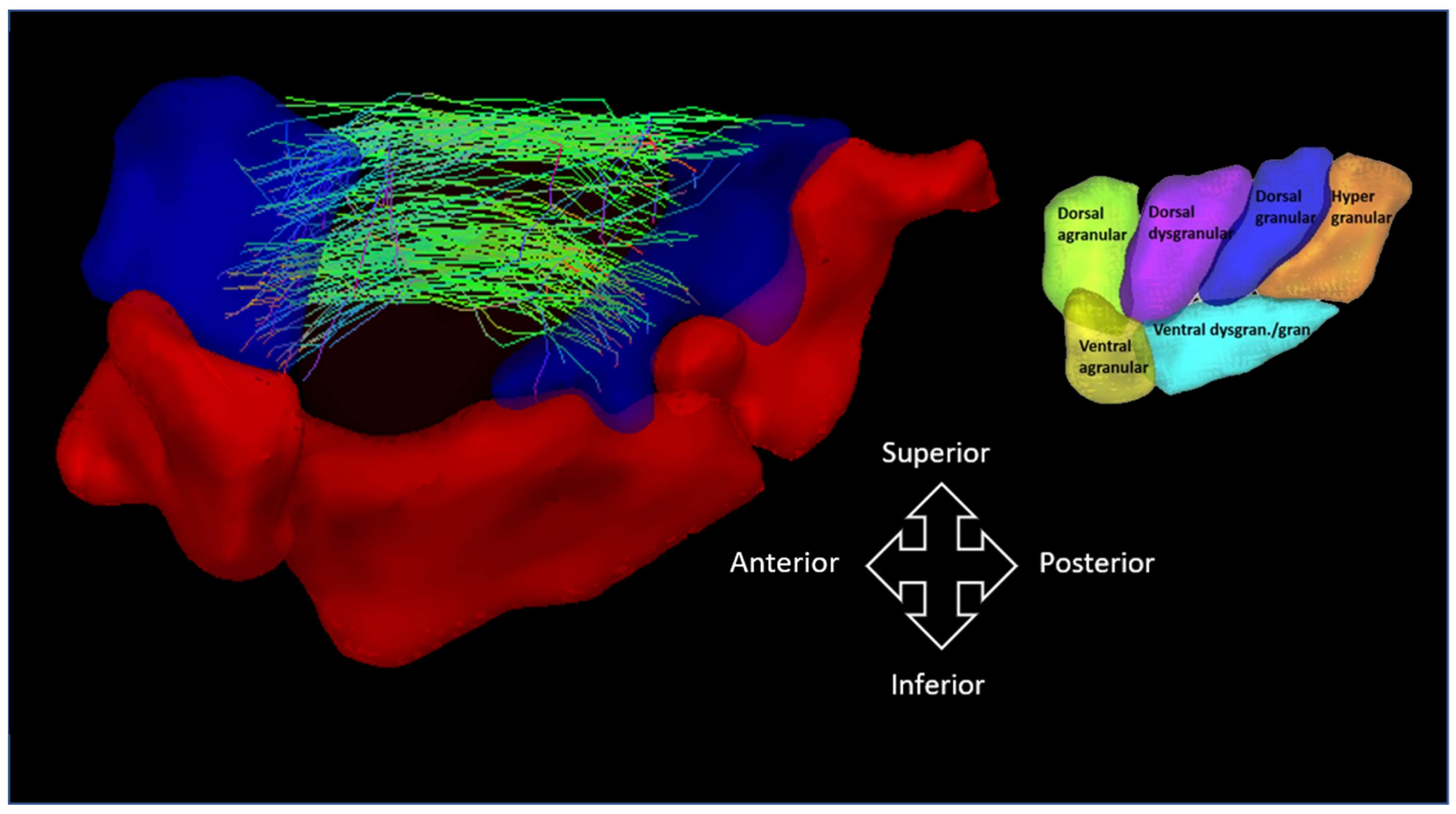
2.4. Image Processing and Connectivity Matrix Construction
2.5. Group Comparisons of COMMIT-Weighted Matrices
2.6. Group Comparisons of Graph Theoretic Measures
2.7. Visualization
3. Results
3.1. Patient Population
3.2. Group Comparisons of COMMIT-Weighted Matrices
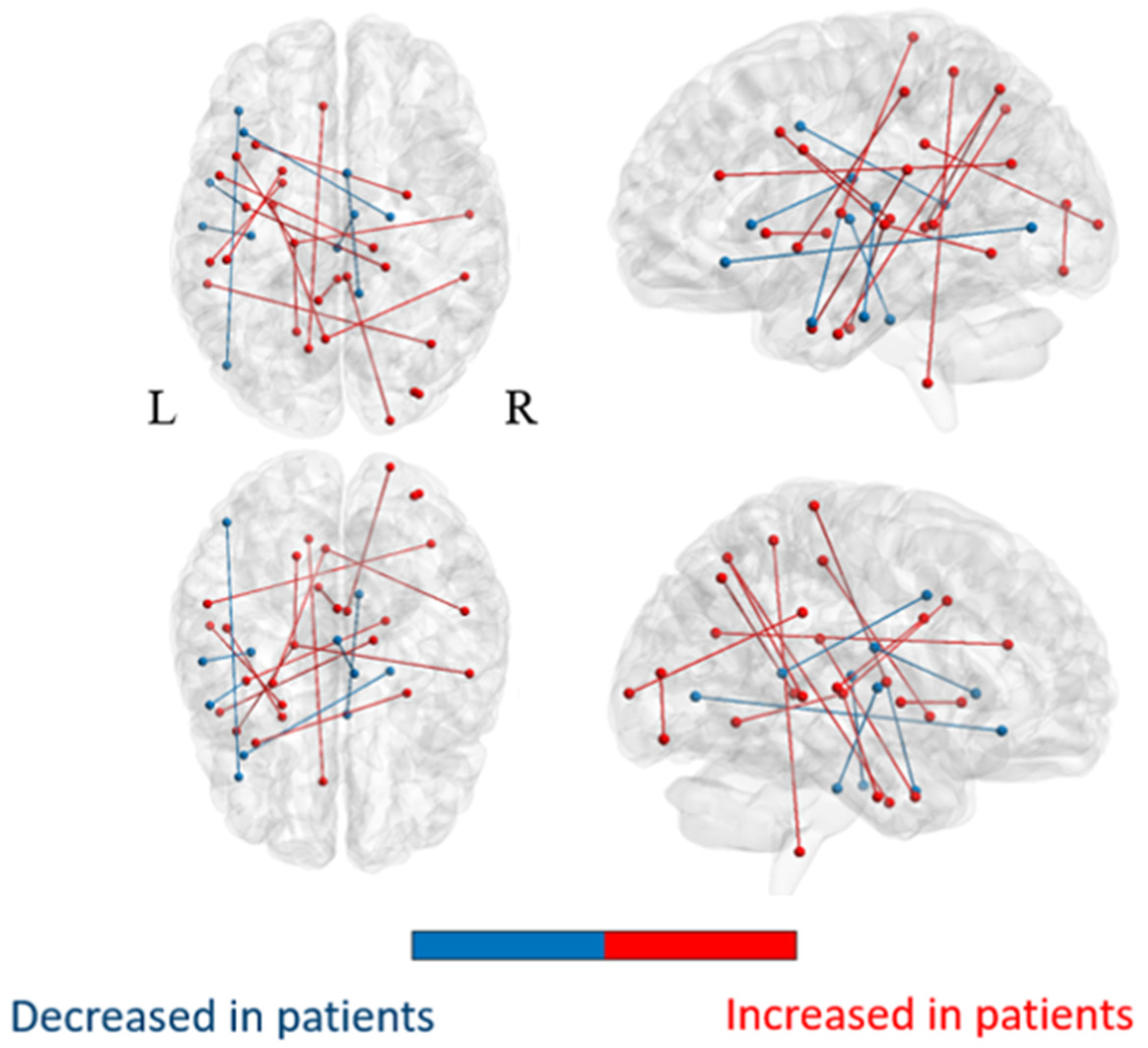
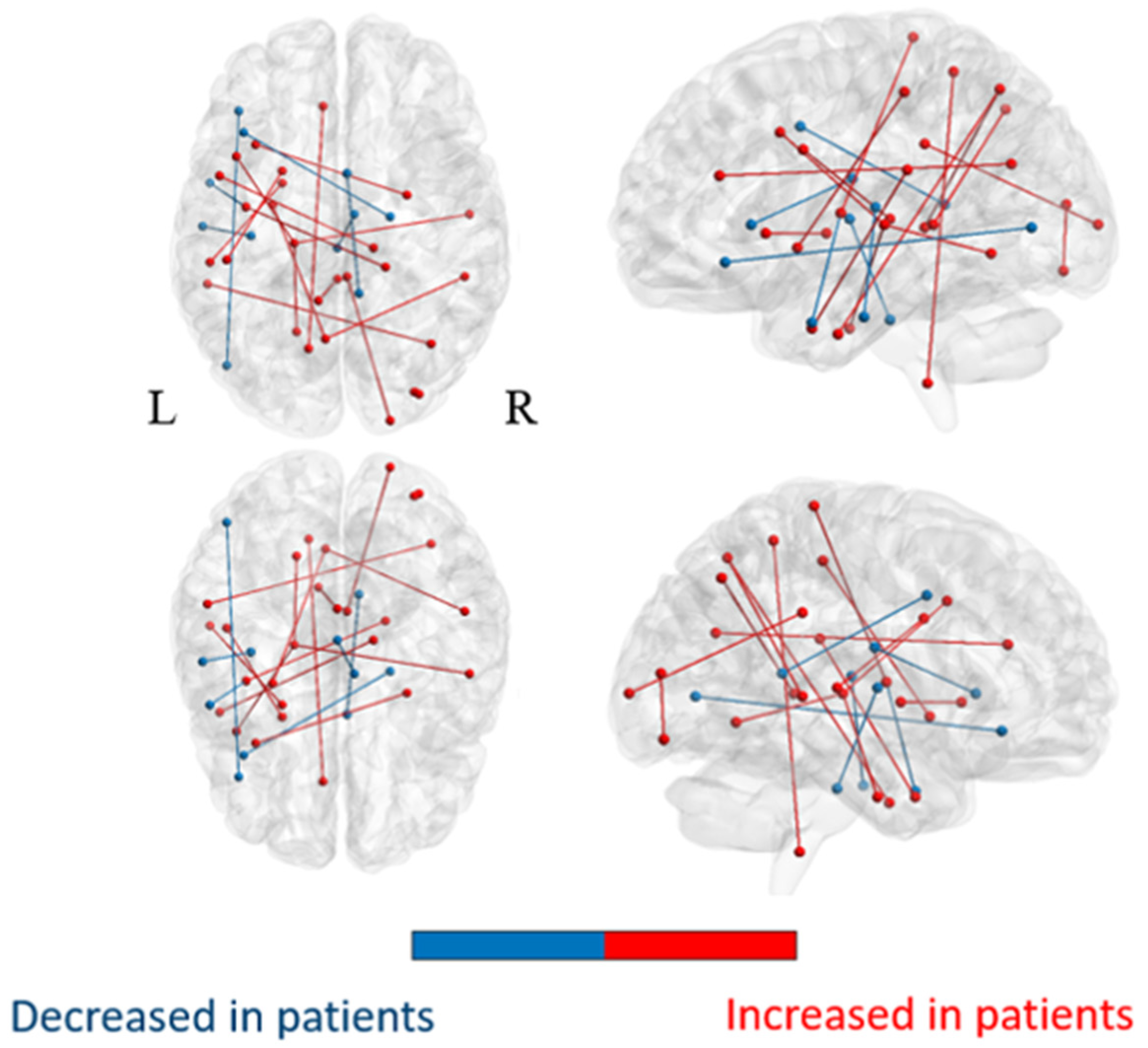
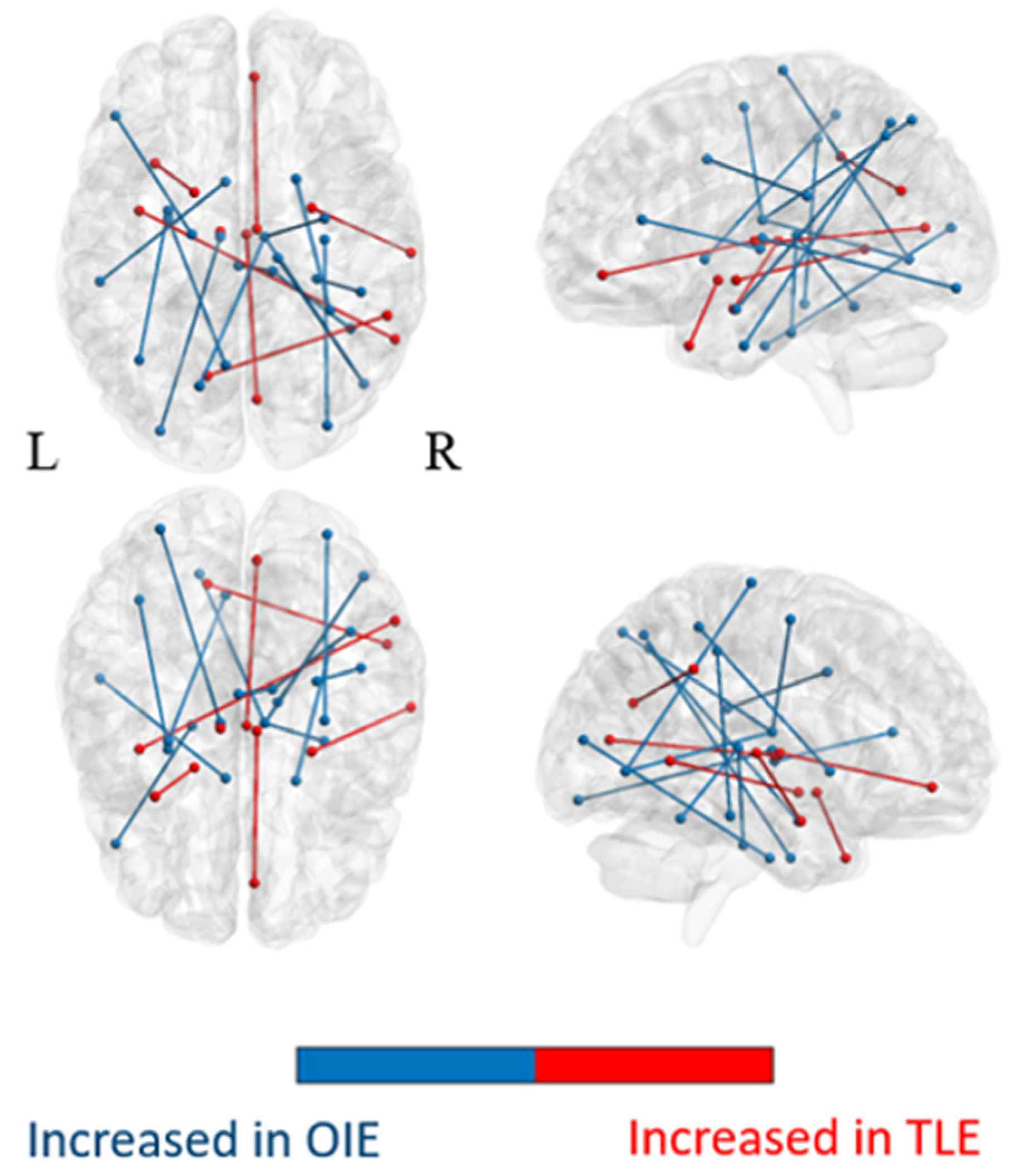
3.2.1. Comparison of the Whole-Brain Network
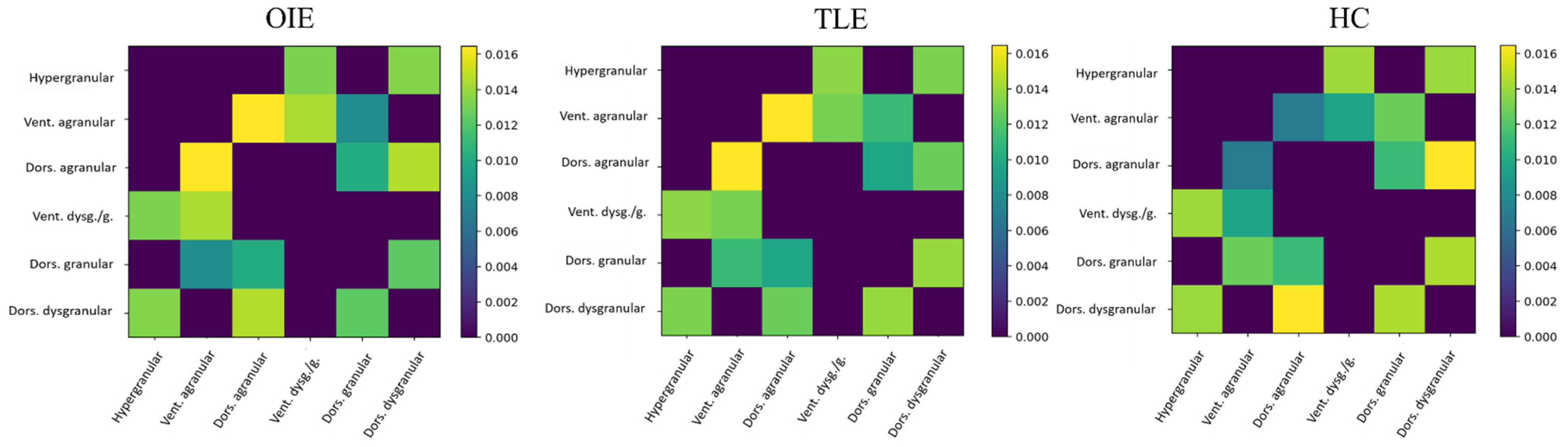
3.2.2. Comparison of Subnetworks
3.3. Group Comparisons of Graph Theoretic Measures
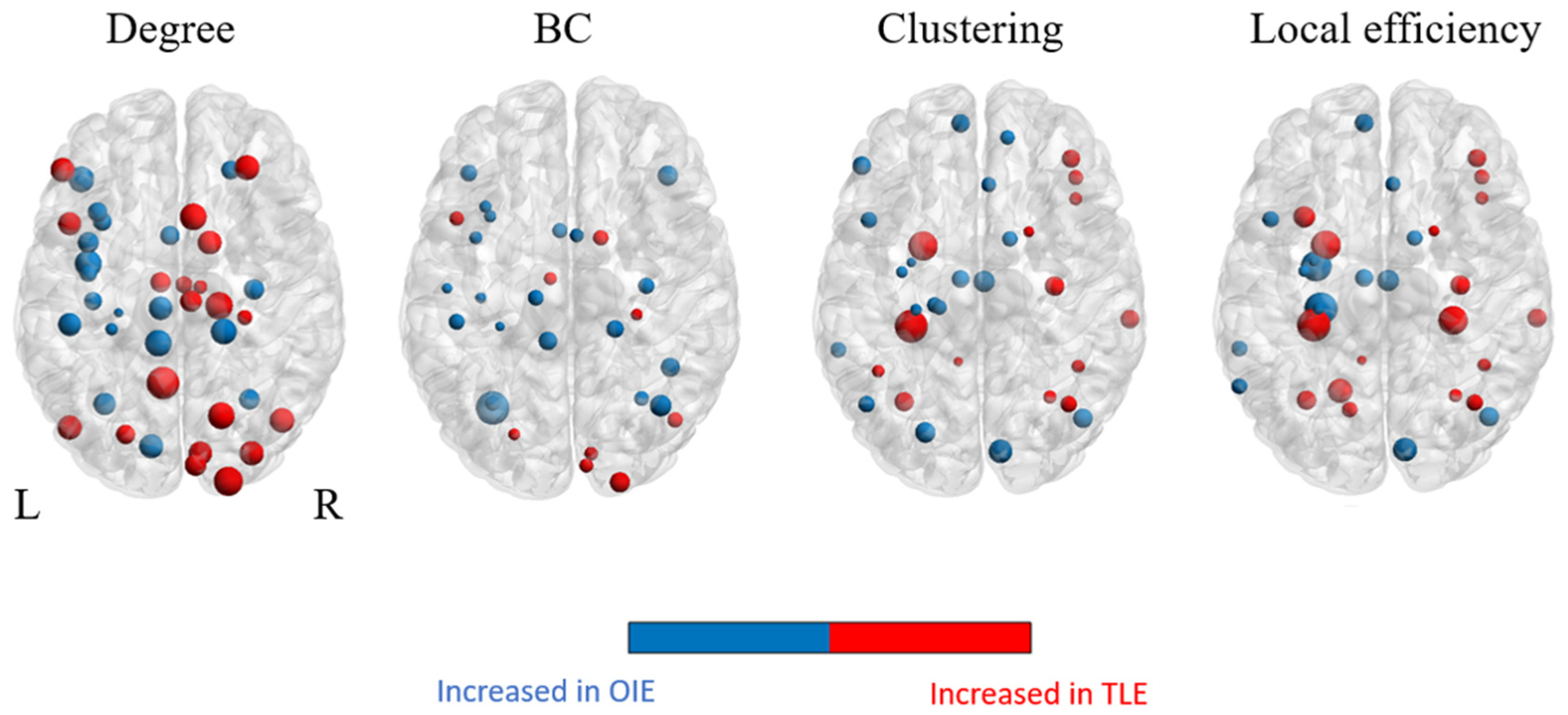
3.3.1. Regional Graphical Properties
3.3.2. Global Graphical Properties
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obaid, S.; Zerouali, Y.; Nguyen, D.K. Insular Epilepsy: Semiology and Noninvasive Investigations. J. Clin. Neurophysiol. 2017, 34, 315–323. [Google Scholar] [CrossRef]
- Cauda, F.; D’Agata, F.; Sacco, K.; Duca, S.; Geminiani, G.; Vercelli, A. Functional connectivity of the insula in the resting brain. Neuroimage 2011, 55, 8–23. [Google Scholar] [CrossRef]
- Deen, B.; Pitskel, N.B.; Pelphrey, K.A. Three systems of insular functional connectivity identified with cluster analysis. Cereb. Cortex 2011, 21, 1498–1506. [Google Scholar] [CrossRef]
- Cerliani, L.; Thomas, R.M.; Jbabdi, S.; Siero, J.C.W.; Nanetti, L.; Crippa, A.; Gazzola, V.; D’Arceuil, H.; Keysers, C. Probabilistic tractography recovers a rostrocaudal trajectory of connectivity variability in the human insular cortex. Hum. Brain Mapp. 2012, 33, 2005–2034. [Google Scholar] [CrossRef] [PubMed]
- Cloutman, L.L.; Binney, R.J.; Drakesmith, M.; Parker, G.J.M.; Lambon Ralph, M.A. The variation of function across the human insula mirrors its patterns of structural connectivity: Evidence from in vivo probabilistic tractography. Neuroimage 2012, 59, 3514–3521. [Google Scholar] [CrossRef] [PubMed]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Boucher, O.; Houde, J.C.; Descoteaux, M.; Obaid, S.; Gilbert, G.; Rouleau, I.; Nguyen, D.K. Subcortical structural connectivity of insular subregions. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ghaziri, J.; Tucholka, A.; Girard, G.; Houde, J.C.; Boucher, O.; Gilbert, G.; Descoteaux, M.; Lippé, S.; Rainville, P.; Nguyen, D.K. The Corticocortical Structural Connectivity of the Human Insula. Cereb. Cortex 2017, 27, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Almashaikhi, T.; Rheims, S.; Ostrowsky-Coste, K.; Montavont, A.; Jung, J.; De Bellescize, J.; Arzimanoglou, A.; Keo Kosal, P.; Guénot, M.; Bertrand, O.; et al. Intrainsular functional connectivity in human. Hum. Brain Mapp. 2014, 35, 2779–2788. [Google Scholar] [CrossRef]
- Nguyen, D.K.; Nguyen, D.B.; Malak, R.; Leroux, J.M.; Carmant, L.; Saint-Hilaire, J.M.; Giard, N.; Cossette, P.; Bouthillier, A. Revisiting the role of the insula in refractory partial epilepsy. Epilepsia 2009, 50, 510–520. [Google Scholar] [CrossRef]
- Gras-Combe, G.; Minotti, L.; Hoffmann, D.; Krainik, A.; Kahane, P.; Chabardes, S. Surgery for Nontumoral Insular Epilepsy Explored by Stereoelectroencephalography. Neurosurgery 2016, 79, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Isnard, J.; Guénot, M.; Sindou, M.; Mauguière, F. Clinical manifestations of insular lobe seizures: A stereo- electroencephalographic study. Epilepsia 2004, 45, 1079–1090. [Google Scholar] [CrossRef]
- Mohamed, I.S.; Gibbs, S.A.; Robert, M.; Bouthillier, A.; Leroux, J.M.; Nguyen, D.K. The utility of magnetoencephalography in the presurgical evaluation of refractory insular epilepsy. Epilepsia 2013, 54, 1950–1959. [Google Scholar] [CrossRef] [Green Version]
- Ryvlin, P.; Minotti, L.; Demarquay, G.; Hirsch, E.; Arzimanoglou, A.; Hoffman, D.; Guénot, M.; Picard, F.; Rheims, S.; Kahane, P. Nocturnal hypermotor seizures, suggesting frontal lobe epilepsy, can originate in the insula. Epilepsia 2006, 47, 755–765. [Google Scholar] [CrossRef]
- Zerouali, Y.; Pouliot, P.; Robert, M.; Mohamed, I.; Bouthillier, A.; Lesage, F.; Nguyen, D.K. Magnetoencephalographic signatures of insular epileptic spikes based on functional connectivity. Hum. Brain Mapp. 2016, 37, 3250–3261. [Google Scholar] [CrossRef]
- Sala-Padro, J.; Fong, M.; Rahman, Z.; Bartley, M.; Gill, D.; Dexter, M.; Bleasel, A.; Wong, C. A study of perfusion changes with Insula Epilepsy using SPECT. Seizure 2019, 69, 44–50. [Google Scholar] [CrossRef]
- Yu, T.; Ni, D.; Zhang, X.; Wang, X.; Qiao, L.; Zhou, X.; Wang, Y.; Li, Y.; Zhang, G. The role of magnetoencephalography in the presurgical evaluation of patients with MRI-negative operculo-insular epilepsy. Seizure 2018, 61, 104–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.; Zhang, X.; Chen, Z.; Li, X.; Wu, S.; Lv, P.; Wang, Y. Detection and localization of interictal ripples with magnetoencephalography in the presurgical evaluation of drug-resistant insular epilepsy. Brain Res. 2019, 1706, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Bou Assi, E.; Rihana, S.; Nguyen, D.K.; Sawan, M. Effective connectivity analysis of iEEG and accurate localization of the epileptogenic focus at the onset of operculo-insular seizures. Epilepsy Res. 2019, 152, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Obaid, S.; Tucholka, A.; Ghaziri, J.; Jodoin, P.M.; Morency, F.; Descoteaux, M.; Bouthillier, A.; Nguyen, D.K. Cortical thickness analysis in operculo-insular epilepsy. NeuroImage Clin. 2018, 19, 727–733. [Google Scholar] [CrossRef] [PubMed]
- James, J.S.; Radhakrishnan, A.; Thomas, B.; Madhusoodanan, M.; Kesavadas, C.; Abraham, M.; Menon, R.; Rathore, C.; Vilanilam, G. Diffusion tensor imaging tractography of Meyer’s loop in planning resective surgery for drug-resistant temporal lobe epilepsy. Epilepsy Res. 2015, 110, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Englot, D.J.; Konrad, P.E.; Morgan, V.L. Regional and global connectivity disturbances in focal epilepsy, related neurocognitive sequelae, and potential mechanistic underpinnings. Epilepsia 2016, 57, 1546–1557. [Google Scholar] [CrossRef] [Green Version]
- Dinkelacker, V.; Valabregue, R.; Thivard, L.; Lehéricy, S.; Baulac, M.; Samson, S.; Dupont, S. Hippocampal-thalamic wiring in medial temporal lobe epilepsy: Enhanced connectivity per hippocampal voxel. Epilepsia 2015, 56, 1217–1226. [Google Scholar] [CrossRef]
- Bonilha, L.; Nesland, T.; Martz, G.U.; Joseph, J.E.; Spampinato, M.V.; Edwards, J.C.; Tabesh, A. Medial temporal lobe epilepsy is associated with neuronal fibre loss and paradoxical increase in structural connectivity of limbic structures. J. Neurol. Neurosurg. Psychiatry 2012, 83, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilha, L.; Helpern, J.A.; Sainju, R.; Nesland, T.; Edwards, J.C.; Glazier, S.S.; Tabesh, A. Presurgical connectome and postsurgical seizure control in temporal lobe epilepsy. Neurology 2013, 81, 1704–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Diessen, E.; Zweiphenning, W.J.E.M.; Jansen, F.E.; Stam, C.J.; Braun, K.P.J.; Otte, W.M. Brain network organization in focal epilepsy: A systematic review and meta-analysis. PLoS ONE 2014, 9, e114606. [Google Scholar] [CrossRef] [Green Version]
- Tavakol, S.; Royer, J.; Lowe, A.J.; Bonilha, L.; Tracy, J.I.; Jackson, G.D.; Duncan, J.S.; Bernasconi, A.; Bernasconi, N.; Bernhardt, B.C. Neuroimaging and connectomics of drug-resistant epilepsy at multiple scales: From focal lesions to macroscale networks. Epilepsia 2019, 60, 593–604. [Google Scholar] [CrossRef]
- Bonilha, L.; Jensen, J.H.; Baker, N.; Breedlove, J.; Nesland, T.; Lin, J.J.; Drane, D.L.; Saindane, A.M.; Binder, J.R.; Kuzniecky, R.I. The brain connectome as a personalized biomarker of seizure outcomes after temporal lobectomy. Neurology 2015, 84, 1846–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, B.C.; Bonilha, L.; Gross, D.W. Network analysis for a network disorder: The emerging role of graph theory in the study of epilepsy. Epilepsy Behav. 2015, 50, 162–170. [Google Scholar] [CrossRef]
- St-Onge, E.; Daducci, A.; Girard, G.; Descoteaux, M. Surface-enhanced tractography (SET). Neuroimage 2018, 169, 524–539. [Google Scholar] [CrossRef]
- Daducci, A.; Dal Palù, A.; Lemkaddem, A.; Thiran, J.P. COMMIT: Convex optimization modeling for microstructure informed tractography. IEEE Trans. Med. Imaging 2015, 34, 246–257. [Google Scholar] [CrossRef] [Green Version]
- Daducci, A.; Dal Palu, A.; Lemkaddem, A.; Thiran, J.P. A Convex Optimization Framework for Global Tractography. In Proceedings of the Proceedings-International Symposium on Biomedical Imaging, San Francisco, CA, USA, 7–11 April 2013. [Google Scholar]
- Descoteaux, M.; Deriche, R.; Knösche, T.R.; Anwander, A. Deterministic and probabilistic tractography based on complex fibre orientation distributions. IEEE Trans. Med. Imaging 2009, 28, 269–286. [Google Scholar] [CrossRef]
- Tournier, J.D.; Calamante, F.; Connelly, A. Robust determination of the fibre orientation distribution in diffusion MRI: Non-negativity constrained super-resolved spherical deconvolution. Neuroimage 2007, 35, 1459–1472. [Google Scholar] [CrossRef]
- Levy, A.; Yen Tran, T.P.; Boucher, O.; Bouthillier, A.; Nguyen, D.K. Operculo-Insular Epilepsy: Scalp and Intracranial Electroencephalographic Findings. J. Clin. Neurophysiol. 2017, 34, 438–447. [Google Scholar] [CrossRef]
- Bouthillier, A.; Weil, A.G.; Martineau, L.; Létourneau-Guillon, L.; Nguyen, D.K. Operculoinsular cortectomy for refractory epilepsy. Part 1: Is it effective? J. Neurosurg. 2020, 133, 950–959. [Google Scholar] [CrossRef]
- Li, X.; Morgan, P.S.; Ashburner, J.; Smith, J.; Rorden, C. The first step for neuroimaging data analysis: DICOM to NIfTI conversion. J. Neurosci. Methods 2016, 264, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Theaud, G.; Houde, J.C.; Boré, A.; Rheault, F.; Morency, F.; Descoteaux, M. TractoFlow: A robust, efficient and reproducible diffusion MRI pipeline leveraging Nextflow & Singularity. Neuroimage 2020, 218. [Google Scholar] [CrossRef]
- Girard, G.; Whittingstall, K.; Deriche, R.; Descoteaux, M. Towards quantitative connectivity analysis: Reducing tractography biases. Neuroimage 2014, 98, 266–278. [Google Scholar] [CrossRef] [PubMed]
- June, S.K.; Singh, V.; Jun, K.L.; Lerch, J.; Ad-Dab’bagh, Y.; MacDonald, D.; Jong, M.L.; Kim, S.I.; Evans, A.C. Automated 3-D extraction and evaluation of the inner and outer cortical surfaces using a Laplacian map and partial volume effect classification. Neuroimage 2005, 27, 210–221. [Google Scholar] [CrossRef]
- Fan, L.; Li, H.; Zhuo, J.; Zhang, Y.; Wang, J.; Chen, L.; Yang, Z.; Chu, C.; Xie, S.; Laird, A.R.; et al. The Human Brainnetome Atlas: A New Brain Atlas Based on Connectional Architecture. Cereb. Cortex 2016, 26, 3508–3526. [Google Scholar] [CrossRef]
- Jones, D.K.; Knösche, T.R.; Turner, R. White matter integrity, fiber count, and other fallacies: The do’s and don’ts of diffusion MRI. Neuroimage 2013, 73, 239–254. [Google Scholar] [CrossRef]
- Maier-Hein, K.H.; Neher, P.F.; Houde, J.C.; Côté, M.A.; Garyfallidis, E.; Zhong, J.; Chamberland, M.; Yeh, F.C.; Lin, Y.C.; Ji, Q.; et al. The challenge of mapping the human connectome based on diffusion tractography. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Rheault, F.; De Benedictis, A.; Daducci, A.; Maffei, C.; Tax, C.M.W.; Romascano, D.; Caverzasi, E.; Morency, F.C.; Corrivetti, F.; Pestilli, F.; et al. Tractostorm: The what, why, and how of tractography dissection reproducibility. Hum. Brain Mapp. 2020, 41, 1859–1874. [Google Scholar] [CrossRef] [Green Version]
- Winkler, A.M.; Ridgway, G.R.; Webster, M.A.; Smith, S.M.; Nichols, T.E. Permutation inference for the general linear model. Neuroimage 2014, 92, 381–397. [Google Scholar] [CrossRef] [Green Version]
- Bassett, D.S.; Bullmore, E. Small-world brain networks. Neuroscientist 2006, 12, 512–523. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. Complex brain networks: Graph theoretical analysis of structural and functional systems. Nat. Rev. Neurosci. 2009, 10, 186–198. [Google Scholar] [CrossRef]
- Sporns, O. Graph theory methods: Applications in brain networks. Dialogues Clin. Neurosci. 2018, 20, 111–120. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef]
- Gleichgerrcht, E.; Kocher, M.; Bonilha, L. Connectomics and graph theory analyses: Novel insights into network abnormalities in epilepsy. Epilepsia 2015, 56, 1660–1668. [Google Scholar] [CrossRef]
- Hosseini, S.M.H.; Hoeft, F.; Kesler, S.R. Gat: A graph-theoretical analysis toolbox for analyzing between-group differences in large-scale structural and functional brain networks. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A Network Visualization Tool for Human Brain Connectomics. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besson, P.; Bandt, S.K.; Proix, T.; Lagarde, S.; Jirsa, V.K.; Ranjeva, J.P.; Bartolomei, F.; Guye, M. Anatomic consistencies across epilepsies: A stereotactic-EEG informed high-resolution structural connectivity study. Brain 2017, 140, 2639–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, S.S. Neural networks in human epilepsy: Evidence of and implications for treatment. Epilepsia 2002, 43, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.N.; Han, C.E.; Schoene-Bake, J.C.; Weber, B.; Kaiser, M. Structural connectivity changes in temporal lobe epilepsy: Spatial features contribute more than topological measures. NeuroImage Clin. 2015, 8, 322–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhardt, B.C.; Fadaie, F.; Liu, M.; Caldairou, B.; Gu, S.; Jefferies, E.; Smallwood, J.; Bassett, D.S.; Bernasconi, A.; Bernasconi, N. Temporal lobe epilepsy: Hippocampal pathology modulates connectome topology and controllability. Neurology 2019, 92, e2209–e2220. [Google Scholar] [CrossRef]
- Bernhardt, B.C.; Worsley, K.J.; Besson, P.; Concha, L.; Lerch, J.P.; Evans, A.C.; Bernasconi, N. Mapping limbic network organization in temporal lobe epilepsy using morphometric correlations: Insights on the relation between mesiotemporal connectivity and cortical atrophy. Neuroimage 2008, 42, 515–524. [Google Scholar] [CrossRef]
- Lin, J.J.; Salamon, N.; Lee, A.D.; Dutton, R.A.; Geaga, J.A.; Hayashi, K.M.; Luders, E.; Toga, A.W.; Engel, J.; Thompson, P.M. Reduced neocortical thickness and complexity mapped in mesial temporal lobe epilepsy with hippocampal sclerosis. Cereb. Cortex 2007, 17, 2007–2018. [Google Scholar] [CrossRef] [Green Version]
- Zerouali, Y.; Ghaziri, J.; Nguyen, D.K. Multimodal Investigation of Epileptic Networks: The Case of Insular Cortex Epilepsy, 1st ed.; Elsevier, B.V.: Amsterdam, The Netherlands, 2016; Volume 226. [Google Scholar]
- Wang, X.; Hu, W.; McGonigal, A.; Zhang, C.; Sang, L.; Zhao, B.; Sun, T.; Wang, F.; Zhang, J.G.; Shao, X.; et al. Electroclinical features of insulo-opercular epilepsy: An SEEG and PET study. Ann. Clin. Transl. Neurol. 2019, 6, 1165–1177. [Google Scholar] [CrossRef]
- DeSalvo, M.N.; Douw, L.; Tanaka, N.; Reinsberger, C.; Stufflebeam, S.M. Altered structural connectome in temporal lobe epilepsy. Radiology 2014, 270, 842–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.N.; Sinha, N.; Wang, Y.; Vos, S.B.; de Tisi, J.; Miserocchi, A.; McEvoy, A.W.; Winston, G.P.; Duncan, J.S. The impact of epilepsy surgery on the structural connectome and its relation to outcome. NeuroImage Clin. 2018, 18, 202–214. [Google Scholar] [CrossRef]
- Ponten, S.C.; Bartolomei, F.; Stam, C.J. Small-world networks and epilepsy: Graph theoretical analysis of intracerebrally recorded mesial temporal lobe seizures. Clin. Neurophysiol. 2007, 118, 918–927. [Google Scholar] [CrossRef]
- Schindler, K.A.; Bialonski, S.; Horstmann, M.T.; Elger, C.E.; Lehnertz, K. Evolving functional network properties and synchronizability during human epileptic seizures. Chaos 2008, 18. [Google Scholar] [CrossRef] [Green Version]
- Concha, L.; Beaulieu, C.; Gross, D.W. Bilateral limbic diffusion abnormalities in unilateral temporal lobe epilepsy. Ann. Neurol. 2005, 57, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Otte, W.M.; Dijkhuizen, R.M.; van Meer, M.P.A.; van der Hel, W.S.; Verlinde, S.A.M.W.; van Nieuwenhuizen, O.; Viergever, M.A.; Stam, C.J.; Braun, K.P.J. Characterization of functional and structural integrity in experimental focal epilepsy: Reduced network efficiency coincides with white matter changes. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Dyhrfjeld-Johnsen, J.; Santhakumar, V.; Morgan, R.J.; Huerta, R.; Tsimring, L.; Soltesz, I. Topological determinants of epileptogenesis in large-scale structural and functional models of the dentate gyrus derived from experimental data. J. Neurophysiol. 2007, 97, 1566–1587. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.S.; Lowenstein, D.H. Mechanisms of disease: Epilepsy. N. Engl. J. Med. 2003, 349. [Google Scholar] [CrossRef]
- Bernhardt, B.C.; Bernasconi, N.; Hong, S.J.; Dery, S.; Bernasconi, A. Subregional mesiotemporal network topology is altered in temporal lobe epilepsy. Cereb. Cortex 2016, 26, 3237–3248. [Google Scholar] [CrossRef] [Green Version]
- Ji, G.J.; Zhang, Z.; Xu, Q.; Wei, W.; Wang, J.; Wang, Z.; Yang, F.; Sun, K.; Jiao, Q.; Liao, W.; et al. Connectome reorganization associated with surgical outcome in temporal lobe epilepsy. Medicine 2015, 94, e1737. [Google Scholar] [CrossRef]
- Webster, J.G.; Descoteaux, M. High Angular Resolution Diffusion Imaging (HARDI). In Wiley Encyclopedia of Electrical and Electronics Engineering; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 1–25. [Google Scholar] [CrossRef]
- Zalesky, A.; Fornito, A.; Cocchi, L.; Gollo, L.L.; van den Heuvel, M.P.; Breakspear, M. Connectome sensitivity or specificity: Which is more important? Neuroimage 2016, 142, 407–420. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age at MRI | Women | Age of Onset | Left-Sided Epilepsy | Duration | |
|---|---|---|---|---|---|
| OIE (n = 9) | 30 ± 8 (18–44) | 7 | 16 ± 10 | 4 | 16 ± 12 |
| TLE (n = 8) | 27 ± 5 (20–34) | 4 | 16.5 ± 10 | 5 | 11 ± 10 |
| Healthy controls (n = 22) | 29 ± 5 (24–40) | 10 | NA | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Obaid, S.; Rheault, F.; Edde, M.; Guberman, G.I.; St-Onge, E.; Sidhu, J.; Bouthillier, A.; Daducci, A.; Ghaziri, J.; Bojanowski, M.W.; et al. Structural Connectivity Alterations in Operculo-Insular Epilepsy. Brain Sci. 2021, 11, 1041. https://doi.org/10.3390/brainsci11081041
Obaid S, Rheault F, Edde M, Guberman GI, St-Onge E, Sidhu J, Bouthillier A, Daducci A, Ghaziri J, Bojanowski MW, et al. Structural Connectivity Alterations in Operculo-Insular Epilepsy. Brain Sciences. 2021; 11(8):1041. https://doi.org/10.3390/brainsci11081041
Chicago/Turabian StyleObaid, Sami, François Rheault, Manon Edde, Guido I. Guberman, Etienne St-Onge, Jasmeen Sidhu, Alain Bouthillier, Alessandro Daducci, Jimmy Ghaziri, Michel W. Bojanowski, and et al. 2021. "Structural Connectivity Alterations in Operculo-Insular Epilepsy" Brain Sciences 11, no. 8: 1041. https://doi.org/10.3390/brainsci11081041
APA StyleObaid, S., Rheault, F., Edde, M., Guberman, G. I., St-Onge, E., Sidhu, J., Bouthillier, A., Daducci, A., Ghaziri, J., Bojanowski, M. W., Nguyen, D. K., & Descoteaux, M. (2021). Structural Connectivity Alterations in Operculo-Insular Epilepsy. Brain Sciences, 11(8), 1041. https://doi.org/10.3390/brainsci11081041