
The Relationship between Short- and Long-Term Memory Is Preserved across the Age Range
Abstract
:1. Introduction
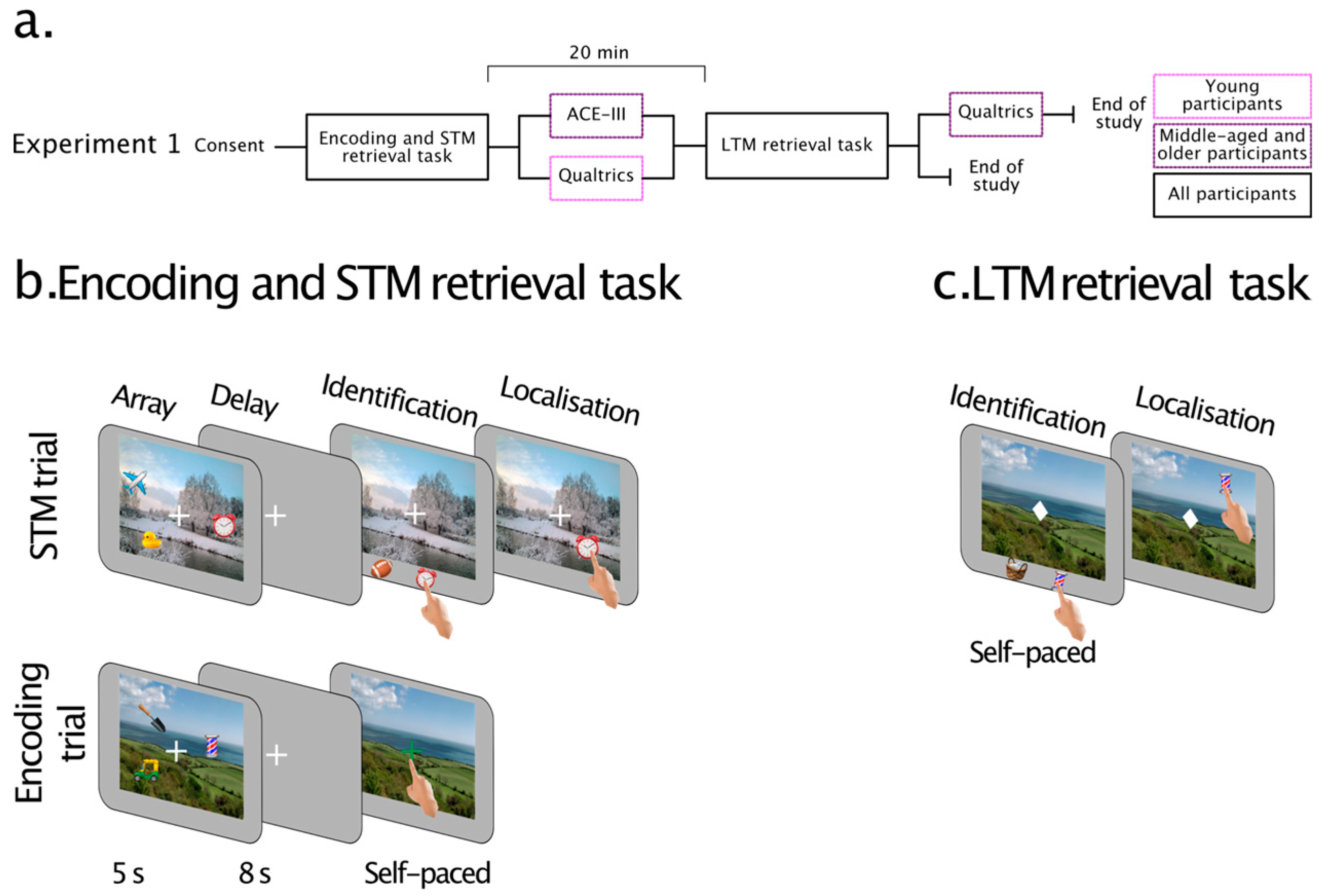
2. Experiment 1
2.1. Methods
2.1.1. Participants
2.1.2. Stimuli
2.1.3. Tasks and Procedures
Encoding and Short-Term Memory Task
Explicit Long-Term Memory Retrieval
2.1.4. Data Analysis
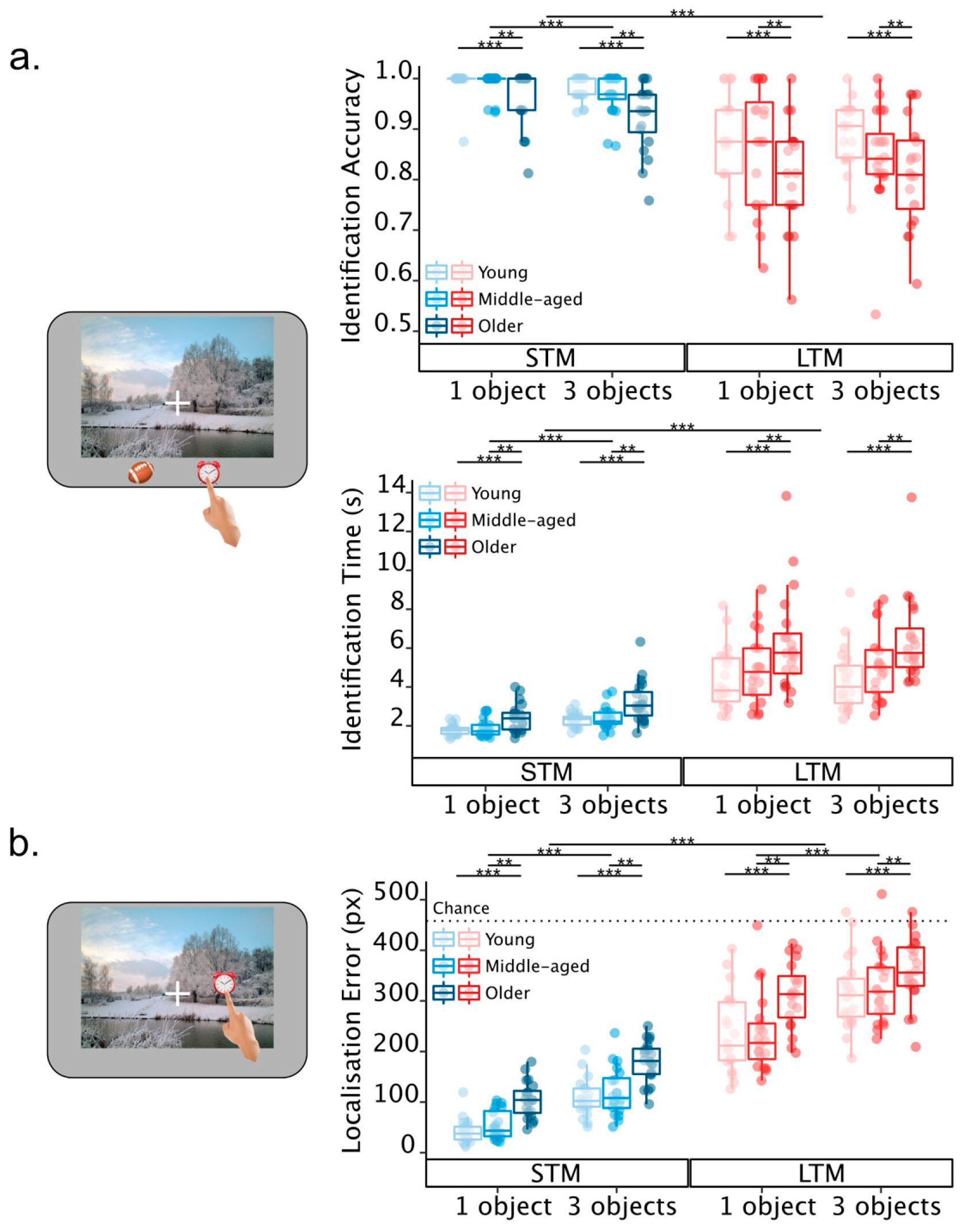
2.2. Results
2.2.1. Participants
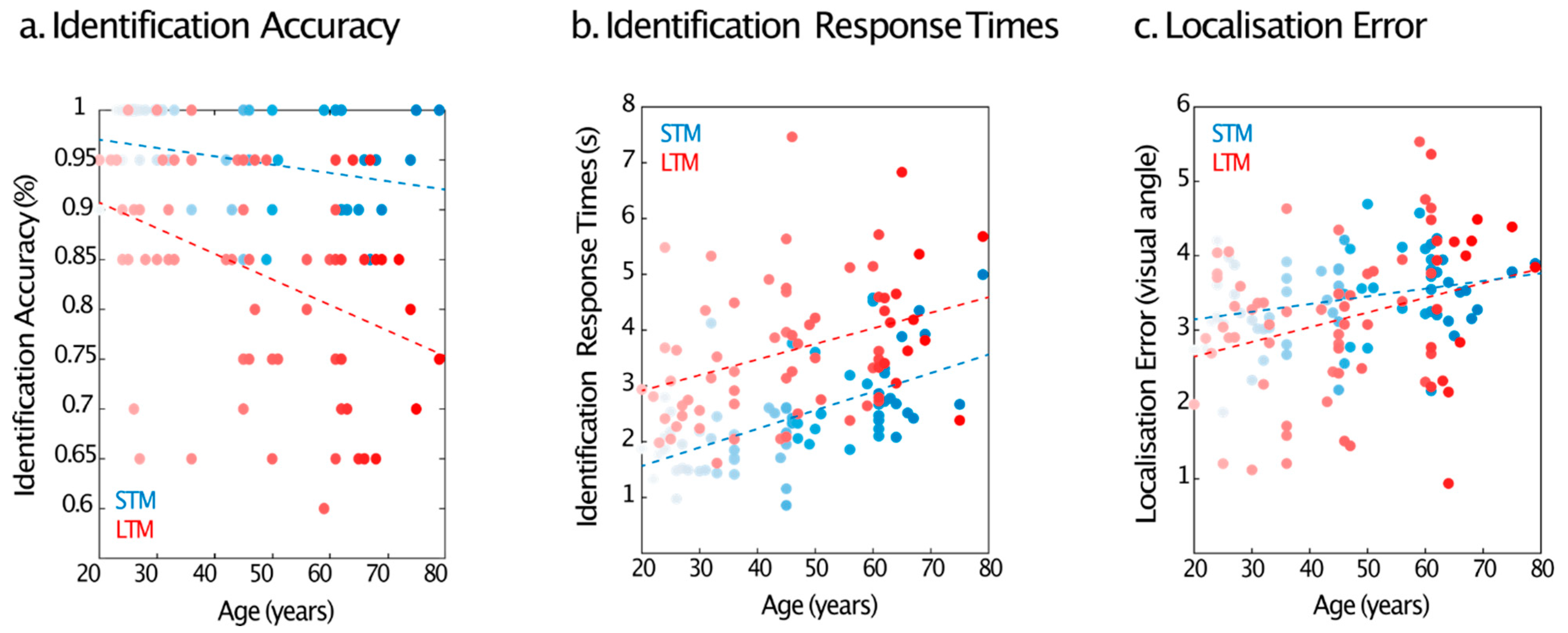
2.2.2. Identification Performance
2.2.3. Localisation Error
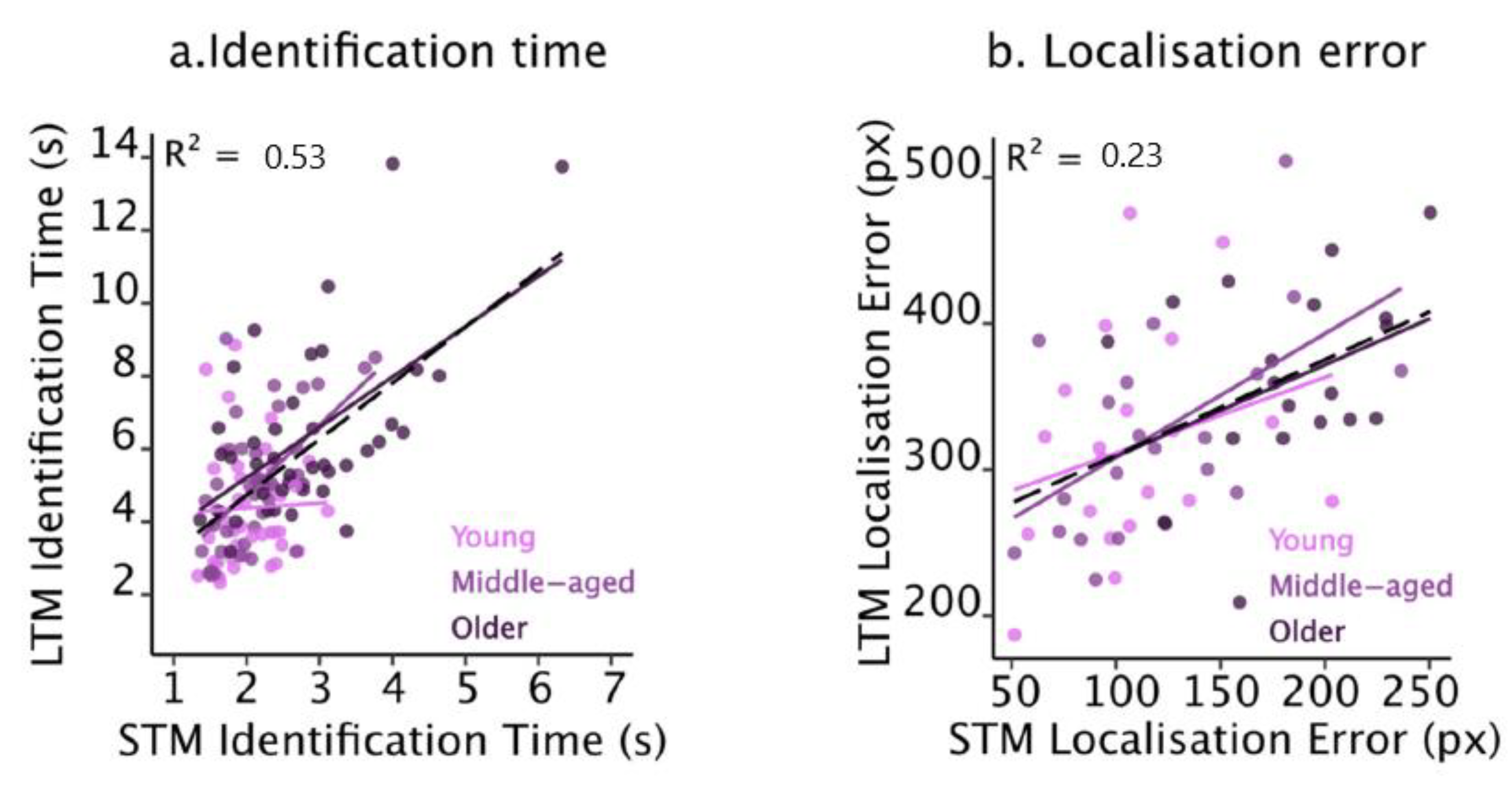
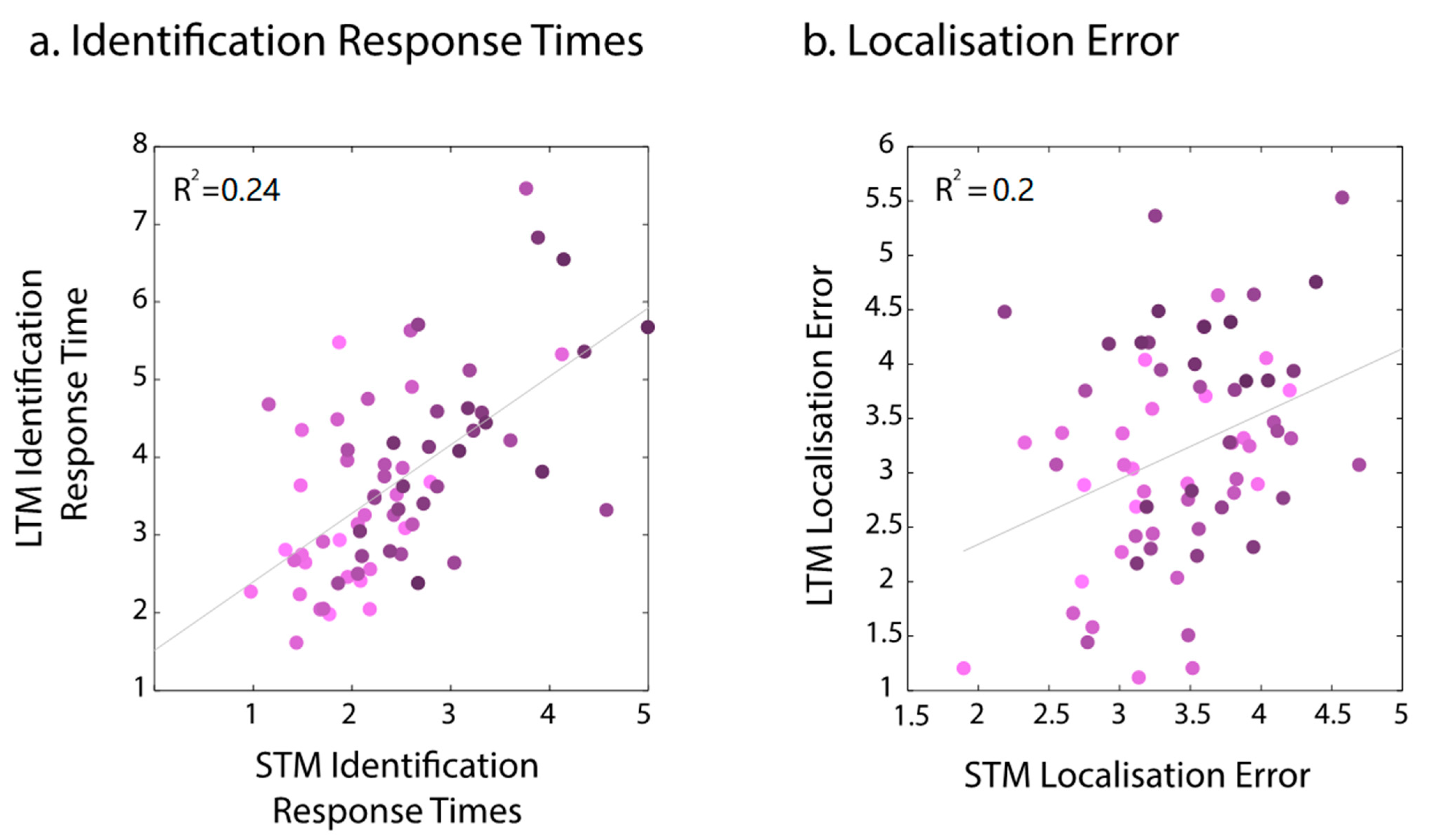
2.2.4. Relationship between Short- and Long-Term Memories
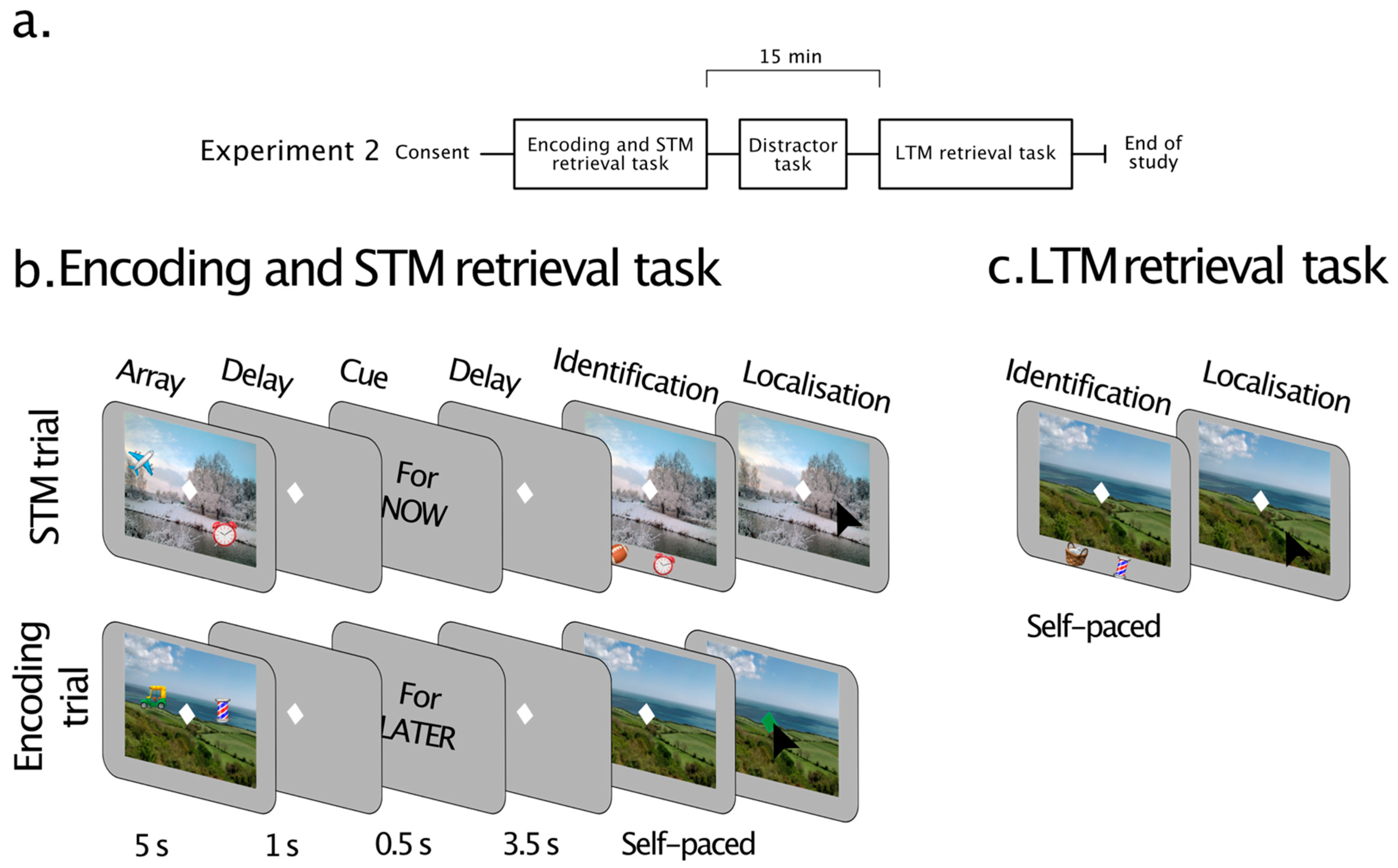
3. Experiment 2
3.1. Methods
3.1.1. Participants
3.1.2. Stimuli
3.1.3. Tasks and Procedures
Short-Term Memory/Encoding Task
Explicit Long-Term Memory Retrieval
3.1.4. Data Analysis
3.2. Results
3.2.1. Identification Performance
3.2.2. Localisation Error
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atkinson, R.C.; Shiffrin, R.M. Human Memory: A Proposed System and Its Control Processes. In Psychology of Learning and Motivation; Academic Press: New York, NY, US, 1968; Volume 2, pp. 89–195. ISBN 978-0-12-543302-0. [Google Scholar]
- Shiffrin, R.M.; Atkinson, R.C. Storage and Retrieval Processes in Long-Term Memory. Psychol. Rev. 1969, 76, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Baddeley, A. The Episodic Buffer: A New Component of Working Memory? Trends Cogn. Sci. 2000, 4, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Hebb, D.O. Distinctive Features of Learning in the Higher Animal; Blackwell: Oxford, UK, 1961. [Google Scholar]
- Oberauer, K.; Lewandowsky, S.; Awh, E.; Brown, G.D.A.; Conway, A.; Cowan, N.; Donkin, C.; Farrell, S.; Hitch, G.J.; Hurlstone, M.J.; et al. Benchmarks for Models of Short-Term and Working Memory. Psychol. Bull. 2018, 144, 885–958. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, A. Working Memory: Theories, Models, and Controversies. Annu. Rev. Psychol. 2012, 63, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Brady, T.F.; Konkle, T.; Gill, J.; Oliva, A.; Alvarez, G.A. Visual Long-Term Memory Has the Same Limit on Fidelity as Visual Working Memory. Psychol. Sci. 2013, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D.A.; Neath, I.; Chater, N. A Temporal Ratio Model of Memory. Psychol. Rev. 2007, 114, 539–576. [Google Scholar] [CrossRef] [Green Version]
- Cowan, N. Memory Theories From A to Z. Contemp. Psychol. A J. Rev. 1995, 40, 552–555. [Google Scholar] [CrossRef]
- Forsberg, A.; Guitard, D.; Cowan, N. Working Memory Limits Severely Constrain Long-Term Retention. Psychon. Bull. Rev. 2021, 28, 537–547. [Google Scholar] [CrossRef]
- Fukuda, K.; Vogel, E.K. Visual Short-Term Memory Capacity Predicts the “Bandwidth” of Visual Long-Term Memory Encoding. Mem. Cogn. 2019, 47, 1481–1497. [Google Scholar] [CrossRef] [Green Version]
- Khader, P.; Ranganath, C.; Seemüller, A.; Rösler, F. Working Memory Maintenance Contributes to Long-Term Memory Formation: Evidence from Slow Event-Related Brain Potentials. Cogn. Affect. Behav. Neurosci. 2007, 7, 212–224. [Google Scholar] [CrossRef]
- Oberauer, K. Access to Information in Working Memory: Exploring the Focus of Attention. J. Exp. Psychol. Learn. Mem. Cogn. 2002, 28, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Ranganath, C.; Cohen, M.X.; Brozinsky, C.J. Working Memory Maintenance Contributes to Long-Term Memory Formation: Neural and Behavioral Evidence. J. Cogn. Neurosci. 2005, 17, 994–1010. [Google Scholar] [CrossRef] [PubMed]
- Schurgin, M.W.; Wixted, J.T.; Brady, T.F. Psychophysical Scaling Reveals a Unified Theory of Visual Memory Strength. Nat. Hum. Behav. 2020, 4, 1156–1172. [Google Scholar] [CrossRef]
- Gorbach, T.; Pudas, S.; Lundquist, A.; Orädd, G.; Josefsson, M.; Salami, A.; de Luna, X.; Nyberg, L. Longitudinal Association between Hippocampus Atrophy and Episodic-Memory Decline. Neurobiol. Aging 2017, 51, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Cam, C.; Henson, R.N.; Campbell, K.L.; Davis, S.W.; Taylor, J.R.; Emery, T.; Erzinclioglu, S.; Kievit, R.A. Multiple Determinants of Lifespan Memory Differences. Sci. Rep. 2016, 6, 32527. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.-G.; BÄCkman, L.; Erngrund, K.; Nyberg, L.; Adolfsson, R.; Bucht, G.; Karlsson, S.; Widing, M.; Winblad, B. The Betula Prospective Cohort Study: Memory, Health, and Aging. Aging Neuropsychol. Cogn. 1997, 4, 1–32. [Google Scholar] [CrossRef]
- Peich, M.-C.; Husain, M.; Bays, P.M. Age-Related Decline of Precision and Binding in Visual Working Memory. Psychol. Aging 2013, 28, 729–743. [Google Scholar] [CrossRef]
- Pertzov, Y.; Heider, M.; Liang, Y.; Husain, M. Effects of Healthy Ageing on Precision and Binding of Object Location in Visual Short Term Memory. Psychol. Aging 2015, 30, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Rönnlund, M.; Nyberg, L.; Bäckman, L.; Nilsson, L.-G. Stability, Growth, and Decline in Adult Life Span Development of Declarative Memory: Cross-Sectional and Longitudinal Data From a Population-Based Study. Psychol. Aging 2005, 20, 3–18. [Google Scholar] [CrossRef]
- Zokaei, N.; Grogan, J.; Fallon, S.J.; Slavkova, E.; Hadida, J.; Manohar, S.; Nobre, A.C.; Husain, M. Short-Term Memory Advantage for Brief Durations in Human APOE Ε4 Carriers. Sci. Rep. 2020, 10, 9503. [Google Scholar] [CrossRef]
- Pudas, S.; Persson, J.; Josefsson, M.; de Luna, X.; Nilsson, L.-G.; Nyberg, L. Brain Characteristics of Individuals Resisting Age-Related Cognitive Decline over Two Decades. J. Neurosci. 2013, 33, 8668–8677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogalski, E.J.; Gefen, T.; Shi, J.; Samimi, M.; Bigio, E.; Weintraub, S.; Geula, C.; Mesulam, M.-M. Youthful Memory Capacity in Old Brains: Anatomic and Genetic Clues from the Northwestern SuperAging Project. J. Cogn. Neurosci. 2013, 25, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, F.W.; Stepanovic, M.R.; Andreano, J.; Barrett, L.F.; Touroutoglou, A.; Dickerson, B.C. Youthful Brains in Older Adults: Preserved Neuroanatomy in the Default Mode and Salience Networks Contributes to Youthful Memory in Superaging. J. Neurosci. 2016, 36, 9659–9668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, B.; Corkin, S.; Teuber, H.-L. Further Analysis of the Hippocampal Amnesic Syndrome: 14-Year Follow-up Study of H.M. Neuropsychologia 1968, 6, 215–234. [Google Scholar] [CrossRef]
- Scoville, W.B.; Milner, B. Loss Of Recent Memory After Bilateral Hippocampal Lesions. J. Neurol. Neurosurg. Psychiatry 1957, 20, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Squire, L.R. The Legacy of Patient H.M. for Neuroscience. Neuron 2009, 61, 6–9. [Google Scholar] [CrossRef] [Green Version]
- Olson, I.R. Working Memory for Conjunctions Relies on the Medial Temporal Lobe. J. Neurosci. 2006, 26, 4596–4601. [Google Scholar] [CrossRef] [Green Version]
- Pertzov, Y.; Miller, T.D.; Gorgoraptis, N.; Caine, D.; Schott, J.M.; Butler, C.; Husain, M. Binding Deficits in Memory Following Medial Temporal Lobe Damage in Patients with Voltage-Gated Potassium Channel Complex Antibody-Associated Limbic Encephalitis. Brain 2013, 136, 2474–2485. [Google Scholar] [CrossRef]
- Watson, P.D.; Voss, J.L.; Warren, D.E.; Tranel, D.; Cohen, N.J. Spatial Reconstruction by Patients with Hippocampal Damage Is Dominated by Relational Memory Errors: Relational Errors in Hippocampal Amnesia. Hippocampus 2013, 23, 570–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zokaei, N.; Nour, M.M.; Sillence, A.; Drew, D.; Adcock, J.; Stacey, R.; Voets, N.; Sen, A.; Husain, M. Binding Deficits in Visual Short-Term Memory in Patients with Temporal Lobe Lobectomy. Hippocampus 2019, 29, 63–67. [Google Scholar] [CrossRef]
- Olson, I.R.; Moore, K.S.; Stark, M.; Chatterjee, A. Visual Working Memory Is Impaired When the Medial Temporal Lobe Is Damaged. J. Cogn. Neurosci. 2006, 18, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Baddeley, A.; Jarrold, C.; Vargha-Khadem, F. Working Memory and the Hippocampus. J. Cogn. Neurosci. 2011, 23, 3855–3861. [Google Scholar] [CrossRef] [PubMed]
- Jeneson, A.; Squire, L.R. Working Memory, Long-Term Memory, and Medial Temporal Lobe Function. Learn. Mem. 2012, 19, 15–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugtmeijer, S.; de Haan, E.H.F.; Kessels, R.P.C. A Comparison of Visual Working Memory and Episodic Memory Performance in Younger and Older Adults. Aging Neuropsychol. Cogn. 2019, 26, 387–406. [Google Scholar] [CrossRef] [Green Version]
- van Geldorp, B.; Heringa, S.M.; van den Berg, E.; Olde Rikkert, M.G.M.; Biessels, G.J.; Kessels, R.P.C. Working Memory Binding and Episodic Memory Formation in Aging, Mild Cognitive Impairment, and Alzheimer’s Dementia. J. Clin. Exp. Neuropsychol. 2015, 37, 538–548. [Google Scholar] [CrossRef]
- Korkki, S.M.; Richter, F.R.; Jeyarathnarajah, P.; Simons, J.S. Healthy Ageing Reduces the Precision of Episodic Memory Retrieval. Psychol. Aging 2020, 35, 124–142. [Google Scholar] [CrossRef]
- Kiyonaga, A.; Scimeca, J.M.; Bliss, D.P.; Whitney, D. Serial Dependence across Perception, Attention, and Memory. Trends Cogn. Sci. 2017, 21, 493–497. [Google Scholar] [CrossRef]
- Makovski, T.; Jiang, Y.V. Proactive Interference from Items Previously Stored in Visual Working Memory. Mem. Cogn. 2008, 36, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.Z.; Fidalgo, C.; Barense, M.D.; Lee, A.C.H.; Cant, J.S.; Ferber, S. Erasing and Blurring Memories: The Differential Impact of Interference on Separate Aspects of Forgetting. J. Exp. Psychol. Gen. 2017, 146, 1606–1630. [Google Scholar] [CrossRef]
- Forsberg, A.; Guitard, D.; Greene, N.R.; Naveh-Benjamin, M.; Cowan, N. The Proportion of Working Memory Items Recoverable from Long-Term Memory Remains Fixed despite Adult Aging. Psychol. Aging 2022, 37, 777–786. [Google Scholar] [CrossRef]
- Peterson, D.J.; Schmidt, N.E.; Naveh-Benjamin, M. The Role of Schematic Support in Age-Related Associative Deficits in Short-Term and Long-Term Memory. J. Mem. Lang. 2017, 92, 79–97. [Google Scholar] [CrossRef]
- Zokaei, N.; Čepukaitytė, G.; Board, A.G.; Mackay, C.E.; Husain, M.; Nobre, A.C. Dissociable Effects of the Apolipoprotein-E (APOE) Gene on Short- and Long-Term Memories. Neurobiol. Aging 2019, 73, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Bohbot, V.D.; Kalina, M.; Stepankova, K.; Spackova, N.; Petrides, M.; Nadel, L. Spatial Memory Deficits in Patients with Lesions to the Right Hippocampus and to the Right Parahippocampal Cortex. Neuropsychologia 1998, 36, 1217–1238. [Google Scholar] [CrossRef] [PubMed]
- Stepankova, K.; Fenton, A.A.; Pastalkova, E.; Kalina, M.; Bohbot, V.D. Object–Location Memory Impairment in Patients with Thermal Lesions to the Right or Left Hippocampus. Neuropsychologia 2004, 42, 1017–1028. [Google Scholar] [CrossRef]
- Brady, T.F.; Konkle, T.; Alvarez, G.A.; Oliva, A. Visual Long-Term Memory Has a Massive Storage Capacity for Object Details. Proc. Natl. Acad. Sci. USA 2008, 105, 14325–14329. [Google Scholar] [CrossRef] [Green Version]
- Brodeur, M.B.; Dionne-Dostie, E.; Montreuil, T.; Lepage, M. The Bank of Standardized Stimuli (BOSS), a New Set of 480 Normative Photos of Objects to Be Used as Visual Stimuli in Cognitive Research. PLoS ONE 2010, 5, e10773. [Google Scholar] [CrossRef] [Green Version]
- Brunner, E.; Puri, M.L. Nonparametric Methods in Factorial Designs. Stat. Pap. 2001, 42, 1–52. [Google Scholar] [CrossRef]
- Noguchi, K.; Gel, Y.R.; Brunner, E.; Konietschke, F. NparLD: An R Software Package for the Nonparametric Analysis of Longitudinal Data in Factorial Experiments. J. Stat. Soft. 2012, 50, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, S.; Schubert, S.; Hoon, C.; Mioshi, E.; Hodges, J.R. Validation of the Addenbrooke’s Cognitive Examination III in Frontotemporal Dementia and Alzheimer’s Disease. Dement. Geriatr. Cogn. Disord. 2013, 36, 242–250. [Google Scholar] [CrossRef]
- Li, Q.; Joo, S.J.; Yeatman, J.D.; Reinecke, K. Controlling for Participants’ Viewing Distance in Large-Scale, Psychophysical Online Experiments Using a Virtual Chinrest. Sci. Rep. 2020, 10, 904. [Google Scholar] [CrossRef]
- Kolarik, B.S.; Baer, T.; Shahlaie, K.; Yonelinas, A.P.; Ekstrom, A.D. Close but No Cigar: Spatial Precision Deficits Following Medial Temporal Lobe Lesions Provide Novel Insight into Theoretical Models of Navigation and Memory. Hippocampus 2018, 28, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Yonelinas, A.P. The Hippocampus Supports High-Resolution Binding in the Service of Perception, Working Memory and Long-Term Memory. Behav. Brain Res. 2013, 254, 34–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamberlain, J.D.; Gagnon, H.; Lalwani, P.; Cassady, K.E.; Simmonite, M.; Seidler, R.D.; Taylor, S.F.; Weissman, D.H.; Park, D.C.; Polk, T.A. GABA Levels in Ventral Visual Cortex Decline with Age and Are Associated with Neural Distinctiveness. Neurobiol. Aging 2021, 102, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Jorge, L.; Canário, N.; Quental, H.; Bernardes, R.; Castelo-Branco, M. Is the Retina a Mirror of the Aging Brain? Aging of Neural Retina Layers and Primary Visual Cortex Across the Lifespan. Front. Aging Neurosci. 2020, 11, 360. [Google Scholar] [CrossRef] [Green Version]
- Pitchaimuthu, K.; Wu, Q.; Carter, O.; Nguyen, B.N.; Ahn, S.; Egan, G.F.; McKendrick, A.M. Occipital GABA Levels in Older Adults and Their Relationship to Visual Perceptual Suppression. Sci. Rep. 2017, 7, 14231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grogan, J.P.; Fallon, S.J.; Zokaei, N.; Husain, M.; Coulthard, E.J.; Manohar, S.G. A New Toolbox to Distinguish the Sources of Spatial Memory Error. J. Vis. 2020, 20, 6. [Google Scholar] [CrossRef]
- Schurgin, M.W. Visual Memory, the Long and the Short of It: A Review of Visual Working Memory and Long-Term Memory. Atten. Percept. Psychophys. 2018, 80, 1035–1056. [Google Scholar] [CrossRef] [Green Version]
- Babu, R.J.; Leat, S.J.; Irving, E.L. Effect of Age and Pop out Distracter on Attended Field of View. J. Optom. 2014, 7, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Owsley, C. Visual Processing Speed. Vis. Res. 2013, 90, 52–56. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young | Middle-Aged | Older | p | |
|---|---|---|---|---|
| n | 20 | 20 | 20 | - |
| Age | 25.2 (4.9) | 49.6 (7.0) | 70.8 (4.4) | - |
| Male/Female | 11/9 | 6/14 | 9/11 | 0.28 |
| Years in education | 17.8 (2.4) | 17.5 (3.4) | 16.8 (4.6) | 0.66 |
| ACE-III Total | - | 96.5 (2.8) | 96.2 (2.1) | 0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čepukaitytė, G.; Thom, J.L.; Kallmayer, M.; Nobre, A.C.; Zokaei, N. The Relationship between Short- and Long-Term Memory Is Preserved across the Age Range. Brain Sci. 2023, 13, 106. https://doi.org/10.3390/brainsci13010106
Čepukaitytė G, Thom JL, Kallmayer M, Nobre AC, Zokaei N. The Relationship between Short- and Long-Term Memory Is Preserved across the Age Range. Brain Sciences. 2023; 13(1):106. https://doi.org/10.3390/brainsci13010106
Chicago/Turabian StyleČepukaitytė, Giedrė, Jude L. Thom, Melvin Kallmayer, Anna C. Nobre, and Nahid Zokaei. 2023. "The Relationship between Short- and Long-Term Memory Is Preserved across the Age Range" Brain Sciences 13, no. 1: 106. https://doi.org/10.3390/brainsci13010106
APA StyleČepukaitytė, G., Thom, J. L., Kallmayer, M., Nobre, A. C., & Zokaei, N. (2023). The Relationship between Short- and Long-Term Memory Is Preserved across the Age Range. Brain Sciences, 13(1), 106. https://doi.org/10.3390/brainsci13010106