
Modal Analysis of Cerebrovascular Effects for Digital Health Integration of Neurostimulation Therapies—A Review of Technology Concepts



Abstract
:1. Introduction
2. Materials and Methods
3. Results
| Pole | Damping | Frequency (rad/seconds) | Time Constant (seconds) |
| (−0.245 + 0 j) | 1 | 0.245 | 4.09 |
| (−3.3 + 0 j) | 1 | 3.3 | 0.303 |
| (−4.9 + 8.44 j) | 0.502 | 9.76 | 0.204 |
| (−4.9 − 8.44 j) | 0.502 | 9.76 | 0.204 |
| (−20.7 + 0 j) | 1 | 20.7 | 0.0483 |
| (−9,590,000 + 0 j) | 1 | 9,590,000 | 1.04 × 10−7 |
| Pole | Damping | Frequency (rad/seconds) | Time Constant (seconds) |
| (−0.245 + 0 j) | 1 | 0.245 | 4.09 |
| (−1 + 0 j) | 1 | 1 | 1 |
| (−3.3 + 0 j) | 1 | 3.3 | 0.303 |
| (−4.9 + 8.44 j) | 0.502 | 9.76 | 0.204 |
| (−4.9 − 8.44j) | 0.502 | 9.76 | 0.204 |
| (−20.7+0j) | 1 | 20.7 | 0.0483 |
| (−29700 + 0 j) | 1 | 29,700 | 3.36 × 10−5 |
| (−9,590,000 + 0 j) | 1 | 9,590,000 | 1.04 × 10−7 |
| Pole | Damping | Frequency (rad/seconds) | Time Constant (seconds) |
| (−0.245 + 0 j) | 1 | 0.245 | 4.09 |
| (−1 + 0 j) | 1 | 1 | 1 |
| (−1.97 + 0 j) | 1 | 1.97 | 0.509 |
| (−3.3 + 0 j) | 1 | 3.3 | 0.303 |
| (−4.9 + 8.44 j) | 0.502 | 9.76 | 0.204 |
| (−4.9−8.44 j) | 0.502 | 9.76 | 0.204 |
| (−15.1 + 0 j) | 1 | 15.1 | 0.0663 |
| (−20.7 + 0 j) | 1 | 20.7 | 0.0483 |
| (−29700 + 0 j) | 1 | 29,700 | 3.36 × 10−5 |
| (−9,590,000 + 0 j) | 1 | 9,590,000 | 1.04 × 10−7 |
| Pole | Damping | Frequency (rad/seconds) | Time Constant (seconds) |
| (−0.245 + 0 j) | 1 | 0.245 | 4.09 |
| (−0.4 + 0 j) | 1 | 0.4 | 2.5 |
| (−1 + 0 j) | 1 | 1 | 1 |
| (−1.97 + 0 j) | 1 | 1.97 | 0.509 |
| (−3.3 + 0 j) | 1 | 3.3 | 0.303 |
| (−4.9 + 8.44 j) | 0.502 | 9.76 | 0.204 |
| (−4.9−8.44 j) | 0.502 | 9.76 | 0.204 |
| (−15.1 + 0 j) | 1 | 15.1 | 0.0663 |
| (−20.7 + 0 j) | 1 | 20.7 | 0.0483 |
| (−29,700 + 0 j) | 1 | 29700 | 3.36 × 10−5 |
| (−9,590,000 + 0 j) | 1 | 9,590,000 | 1.04 × 10−7 |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zheng, X.; Alsop, D.C.; Schlaug, G. Effects of Transcranial Direct Current Stimulation (tDCS) on Human Regional Cerebral Blood Flow. NeuroImage 2011, 58, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Arora, Y.; Dutta, A. Human-in-the-Loop Optimization of Transcranial Electrical Stimulation at the Point of Care: A Computational Perspective. Brain Sci. 2022, 12, 1294. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.J.; Bouret, S. Orienting and Reorienting: The Locus Coeruleus Mediates Cognition through Arousal. Neuron 2012, 76, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Witts, E.C.; Mathews, M.A.; Murray, A.J. The Locus Coeruleus Directs Sensory-Motor Reflex Amplitude across Environmental Contexts. Curr. Biol. CB 2023, 33, 4679–4688.e3. [Google Scholar] [CrossRef] [PubMed]
- Ogoh, S.; Tarumi, T. Cerebral Blood Flow Regulation and Cognitive Function: A Role of Arterial Baroreflex Function. J. Physiol. Sci. 2019, 69, 813–823. [Google Scholar] [CrossRef] [PubMed]
- Silvani, A.; Calandra-Buonaura, G.; Benarroch, E.E.; Dampney, R.A.L.; Cortelli, P. Bidirectional Interactions between the Baroreceptor Reflex and Arousal: An Update. Sleep Med. 2015, 16, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Silva-Filho, E.; Albuquerque, J.; Bikson, M.; Pegado, R.; da Cruz Santos, A.; do Socorro Brasileiro-Santos, M. Effects of Transcranial Direct Current Stimulation Associated with an Aerobic Exercise Bout on Blood Pressure and Autonomic Modulation of Hypertensive Patients: A Pilot Randomized Clinical Trial. Auton. Neurosci. 2021, 235, 102866. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, B.; Barboza, C.A.; Moura, E.G.; Ministro, G.; Ferreira-Melo, S.E.; Castaño, J.B.; Nunes, W.M.S.; Mostarda, C.; Coca, A.; Vianna, L.C.; et al. Acute and Short-Term Autonomic and Hemodynamic Responses to Transcranial Direct Current Stimulation in Patients With Resistant Hypertension. Front. Cardiovasc. Med. 2022, 9, 853427. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.Y.; Lee, M. Autonomic Dysfunction, Diabetes and Metabolic Syndrome. J. Diabetes Investig. 2021, 12, 2108–2111. [Google Scholar] [CrossRef]
- Pillai, J.A.; Bena, J.; Bekris, L.; Kodur, N.; Kasumov, T.; Leverenz, J.B.; Kashyap, S.R.; on behalf of the Alzheimer’s Disease Neuroimaging Initiative. Metabolic Syndrome Biomarkers Relate to Rate of Cognitive Decline in MCI and Dementia Stages of Alzheimer’s Disease. Alzheimers Res. Ther. 2023, 15, 54. [Google Scholar] [CrossRef]
- Giorgi, F.S.; Galgani, A.; Puglisi-Allegra, S.; Limanaqi, F.; Busceti, C.L.; Fornai, F. Locus Coeruleus and Neurovascular Unit: From Its Role in Physiology to Its Potential Role in Alzheimer’s Disease Pathogenesis. J. Neurosci. Res. 2020, 98, 2406–2434. [Google Scholar] [CrossRef] [PubMed]
- Arora, Y.; Walia, P.; Hayashibe, M.; Muthalib, M.; Chowdhury, S.R.; Perrey, S.; Dutta, A. Grey-Box Modeling and Hypothesis Testing of Functional near-Infrared Spectroscopy-Based Cerebrovascular Reactivity to Anodal High-Definition tDCS in Healthy Humans. PLOS Comput. Biol. 2021, 17, e1009386. [Google Scholar] [CrossRef] [PubMed]
- Modal Analysis—1st Edition. Available online: https://shop.elsevier.com/books/modal-analysis/fu/978-0-7506-5079-3 (accessed on 19 December 2023).
- Modal Analysis for Damage Detection in Structures—Semantic Scholar. Available online: https://www.semanticscholar.org/paper/Modal-Analysis-for-Damage-Detection-in-Structures-Hearn-Testa/38a4691979855d965640f076820b6acc406feae9 (accessed on 19 December 2023).
- Rogers, G. Modal Analysis for Control. In Power System Oscillations; Rogers, G., Ed.; The Springer International Series in Engineering and Computer Science; Springer US: Boston, MA, USA, 2000; pp. 75–100. ISBN 978-1-4615-4561-3. [Google Scholar]
- Dutta, A.; Jacob, A.; Chowdhury, S.R.; Das, A.; Nitsche, M.A. EEG-NIRS Based Assessment of Neurovascular Coupling during Anodal Transcranial Direct Current Stimulation—A Stroke Case Series. J. Med. Syst. 2015, 39, 205. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Besson, P.; Muthalib, M.; Jindal, U.; Perrey, S.; Dutta, A.; Hayashibe, M. NIRS-EEG Joint Imaging during Transcranial Direct Current Stimulation: Online Parameter Estimation with an Autoregressive Model. J. Neurosci. Methods 2016, 274, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Arora, Y.; Dutta, A. Perspective: Disentangling the Effects of tES on Neurovascular Unit. Front. Neurol. 2023, 13, 1038700. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.-R.; Colenbier, N.; Van Den Bossche, S.; Clauw, K.; Johri, A.; Tandon, M.; Marinazzo, D. rsHRF: A Toolbox for Resting-State HRF Estimation and Deconvolution. NeuroImage 2021, 244, 118591. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.W.; Fan, J.; Luu, E.; Khalid, W.; Xia, Y.; Khadka, N.; Bikson, M.; Fu, B.M. In Vivo Modulation of the Blood-Brain Barrier Permeability by Transcranial Direct Current Stimulation (tDCS). Ann. Biomed. Eng. 2020, 48, 1256–1270. [Google Scholar] [CrossRef]
- Hertz, L.; Lovatt, D.; Goldman, S.A.; Nedergaard, M. Adrenoceptors in Brain: Cellular Gene Expression and Effects on Astrocytic Metabolism and [Ca2+]i. Neurochem. Int. 2010, 57, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Moshkforoush, A.; Ashenagar, B.; Harraz, O.F.; Dabertrand, F.; Longden, T.A.; Nelson, M.T.; Tsoukias, N.M. The Capillary Kir Channel as Sensor and Amplifier of Neuronal Signals: Modeling Insights on K+-Mediated Neurovascular Communication. Proc. Natl. Acad. Sci. 2020, 117, 16626–16637. [Google Scholar] [CrossRef]
- Reitman, M.E.; Tse, V.; Mi, X.; Willoughby, D.D.; Peinado, A.; Aivazidis, A.; Myagmar, B.-E.; Simpson, P.C.; Bayraktar, O.A.; Yu, G.; et al. Norepinephrine Links Astrocytic Activity to Regulation of Cortical State. Nat. Neurosci. 2023, 26, 579–593. [Google Scholar] [CrossRef]
- Vanneste, S.; Mohan, A.; Yoo, H.B.; Huang, Y.; Luckey, A.M.; McLeod, S.L.; Tabet, M.N.; Souza, R.R.; McIntyre, C.K.; Chapman, S.; et al. The Peripheral Effect of Direct Current Stimulation on Brain Circuits Involving Memory. Sci. Adv. 2020, 6, eaax9538. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Tomita, M.; Lahiri, U.; Das, A. NeuroMind: Eye Tracking and Portable Neuroimaging to Monitor Multidomain Lifestyle Intervention for Healthy Ageing, In Proceedings of the Dementia Research at UCLan and Beyond: An Event for Scientists and the Public, Preston, UK, 21 July 2023.
- Sheng, Y.; Zhu, L. The Crosstalk between Autonomic Nervous System and Blood Vessels. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 17–28. [Google Scholar] [PubMed]
- Kuo, H.; Paulus, W.; Batsikadze, G.; Jamil, A.; Kuo, M.; Nitsche, M.A. Acute and Chronic Effects of Noradrenergic Enhancement on Transcranial Direct Current Stimulation-induced Neuroplasticity in Humans. J. Physiol. 2017, 595, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Khadka, N.; Bikson, M. Neurocapillary-Modulation. Neuromodul. J. Int. Neuromodul. Soc. 2020, 25, 1299–1311. [Google Scholar] [CrossRef] [PubMed]
- Bleys, R.L.A.W.; Cowen, T.; Groen, G.J.; Hillen, B.; Ibrahim, N.B.N. Perivascular Nerves of the Human Basal Cerebral Arteries: I. Topographical Distribution. J. Cereb. Blood Flow Metab. 1996, 16, 1034–1047. [Google Scholar] [CrossRef] [PubMed]
- Grizzanti, J.; Moritz, W.R.; Pait, M.C.; Stanley, M.; Kaye, S.D.; Carroll, C.M.; Constantino, N.J.; Deitelzweig, L.J.; Snipes, J.A.; Kellar, D.; et al. KATP Channels Are Necessary for Glucose-Dependent Increases in Amyloid-β and Alzheimer’s Disease–Related Pathology. JCI Insight 2023, 8, e162454. [Google Scholar] [CrossRef] [PubMed]
- Bekar, L.K.; Wei, H.S.; Nedergaard, M. The Locus Coeruleus-Norepinephrine Network Optimizes Coupling of Cerebral Blood Volume with Oxygen Demand. J. Cereb. Blood Flow Metab. 2012, 32, 2135–2145. [Google Scholar] [CrossRef]
- Korte, N.; James, G.; You, H.; Hirunpattarasilp, C.; Christie, I.; Sethi, H.; Attwell, D. Noradrenaline Released from Locus Coeruleus Axons Contracts Cerebral Capillary Pericytes via A2 Adrenergic Receptors. J. Cereb. Blood Flow Metab. Off. J. Int. Soc. Cereb. Blood Flow Metab. 2023, 43, 1142–1152. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Fang, Z.; Chen, W. Quick and Effective Hyperpolarization of the Membrane Potential in Intact Smooth Muscle Cells of Blood Vessels by Synchronization Modulation Electric Field. J. Bioenerg. Biomembr. 2012, 44, 385–395. [Google Scholar] [CrossRef]
- Koep, J.L.; Taylor, C.E.; Coombes, J.S.; Bond, B.; Ainslie, P.N.; Bailey, T.G. Autonomic Control of Cerebral Blood Flow: Fundamental Comparisons between Peripheral and Cerebrovascular Circulations in Humans. J. Physiol. 2022, 600, 15–39. [Google Scholar] [CrossRef]
- Claassen, J.A.H.R.; Thijssen, D.H.J.; Panerai, R.B.; Faraci, F.M. Regulation of Cerebral Blood Flow in Humans: Physiology and Clinical Implications of Autoregulation. Physiol. Rev. 2021, 101, 1487–1559. [Google Scholar] [CrossRef] [PubMed]
- LONGDEN, T.A.; NELSON, M.T. Vascular Inward Rectifier K+ Channels as External K+ Sensors in the Control of Cerebral Blood Flow. Microcirc. N. Y. N 1994 2015, 22, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Tomita, M.; Dutta, A. Operational Modal Analysis of Near-Infrared Spectroscopy Measure of 2-Month Exercise Intervention Effects in Sedentary Older Adults with Diabetes and Cognitive Impairment. Brain Sci. 2023, 13, 1099. [Google Scholar] [CrossRef]
- Mauri, P.; Miniussi, C.; Balconi, M.; Brignani, D. Bursts of Transcranial Electrical Stimulation Increase Arousal in a Continuous Performance Test. Neuropsychologia 2015, 74, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, G.; Ottaviani, C.; Fiorucci, C.; Petrocchi, N.; Moscucci, F.; Di Iorio, C.; Mastropietri, F.; Parrotta, I.; Pascucci, M.; Magrì, D. Transcranial Direct Current Stimulation Improves the QT Variability Index and Autonomic Cardiac Control in Healthy Subjects Older than 60 Years. Clin. Interv. Aging 2016, 11, 1687–1695. [Google Scholar] [CrossRef] [PubMed]
- Sherwood, M.S.; Madaris, A.T.; Mullenger, C.R.; McKinley, R.A. Repetitive Transcranial Electrical Stimulation Induces Quantified Changes in Resting Cerebral Perfusion Measured from Arterial Spin Labeling. Neural Plast. 2018, 2018, 5769861. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A. Bidirectional Interactions between Neuronal and Hemodynamic Responses to Transcranial Direct Current Stimulation (tDCS): Challenges for Brain-State Dependent tDCS. Front. Syst. Neurosci. 2015, 9, 107. [Google Scholar] [CrossRef]
- Dutta, A. Simultaneous Functional Near-Infrared Spectroscopy (fNIRS) and Electroencephalogram (EEG) to Elucidate Neurovascular Modulation by Transcranial Electrical Stimulation (tES). Brain Stimulat. 2021, 14, 1093–1094. [Google Scholar] [CrossRef]
- Steenland, K.; Karnes, C.; Seals, R.; Carnevale, C.; Hermida, A.; Levey, A. Late-Life Depression as a Risk Factor for Mild Cognitive Impairment or Alzheimer’s Disease in 30 US Alzheimer’s Disease Centers. J. Alzheimers Dis. JAD 2012, 31, 265–275. [Google Scholar] [CrossRef]
- Najjar, S.; Pearlman, D.M.; Devinsky, O.; Najjar, A.; Zagzag, D. Neurovascular Unit Dysfunction with Blood-Brain Barrier Hyperpermeability Contributes to Major Depressive Disorder: A Review of Clinical and Experimental Evidence. J. Neuroinflammation 2013, 10, 142. [Google Scholar] [CrossRef]
- Practical Guide to Transcranial Direct Current Stimulation: Principles, Procedures and Applications; Knotkova, H.; Nitsche, M.A.; Bikson, M.; Woods, A.J. (Eds.) Springer International Publishing: Cham, Switzerland, 2019; ISBN 978-3-319-95947-4. [Google Scholar]
- Bergmann, T.O.; Groppa, S.; Seeger, M.; Mölle, M.; Marshall, L.; Siebner, H.R. Acute Changes in Motor Cortical Excitability during Slow Oscillatory and Constant Anodal Transcranial Direct Current Stimulation. J. Neurophysiol. 2009, 102, 2303–2311. [Google Scholar] [CrossRef] [PubMed]
- Vieira, P.G.; Krause, M.R.; Pack, C.C. tACS Entrains Neural Activity While Somatosensory Input Is Blocked. PLoS Biol. 2020, 18, e3000834. [Google Scholar] [CrossRef] [PubMed]
- Taira, K.; Brunton, S.L.; Dawson, S.T.M.; Rowley, C.W.; Colonius, T.; McKeon, B.J.; Schmidt, O.T.; Gordeyev, S.; Theofilis, V.; Ukeiley, L.S. Modal Analysis of Fluid Flows: An Overview. AIAA J. 2017, 55, 4013–4041. [Google Scholar] [CrossRef]
- Gruszecki, M.; Nuckowska, M.K.; Szarmach, A.; Radkowski, M.; Szalewska, D.; Waskow, M.; Szurowska, E.; Frydrychowski, A.F.; Demkow, U.; Winklewski, P.J. Oscillations of Subarachnoid Space Width as a Potential Marker of Cerebrospinal Fluid Pulsatility. Adv. Exp. Med. Biol. 2018, 1070, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Svinkunaite, L.; Horschig, J.M.; Floor-Westerdijk, M.J. Employing Cardiac and Respiratory Features Extracted from fNIRS Signals for Mental Workload Classification. In Proceedings of the Biophotonics in Exercise Science, Sports Medicine, Health Monitoring Technologies, and Wearables II, Online, 5 March 2021; SPIE: Bellingham, DC, USA, 2021; Volume 11638, pp. 53–61. [Google Scholar]
- Mestre, H.; Tithof, J.; Du, T.; Song, W.; Peng, W.; Sweeney, A.M.; Olveda, G.; Thomas, J.H.; Nedergaard, M.; Kelley, D.H. Flow of Cerebrospinal Fluid Is Driven by Arterial Pulsations and Is Reduced in Hypertension. Nat. Commun. 2018, 9, 4878. [Google Scholar] [CrossRef] [PubMed]
- Vijayakrishnan Nair, V.; Kish, B.R.; Inglis, B.; Yang, H.-C.; Wright, A.M.; Wu, Y.-C.; Zhou, X.; Schwichtenberg, A.J.; Tong, Y. Human CSF Movement Influenced by Vascular Low Frequency Oscillations and Respiration. Front. Physiol. 2022, 13, 940140. [Google Scholar] [CrossRef]
- Murdock, M.H.; Yang, C.-Y.; Sun, N.; Pao, P.-C.; Blanco-Duque, C.; Kahn, M.C.; Kim, T.; Lavoie, N.S.; Victor, M.B.; Islam, M.R.; et al. Multisensory Gamma Stimulation Promotes Glymphatic Clearance of Amyloid. Nature 2024, 627, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Jiang-Xie, L.-F.; Drieu, A.; Bhasiin, K.; Quintero, D.; Smirnov, I.; Kipnis, J. Neuronal Dynamics Direct Cerebrospinal Fluid Perfusion and Brain Clearance. Nature 2024, 1–8. [Google Scholar] [CrossRef]
- Zhang, D.; Ruan, J.; Peng, S.; Li, J.; Hu, X.; Zhang, Y.; Zhang, T.; Ge, Y.; Zhu, Z.; Xiao, X.; et al. Synaptic-like Transmission between Neural Axons and Arteriolar Smooth Muscle Cells Drives Cerebral Neurovascular Coupling. Nat. Neurosci. 2024. [Google Scholar] [CrossRef]
- Faghih, M.M.; Sharp, M.K. Is Bulk Flow Plausible in Perivascular, Paravascular and Paravenous Channels? Fluids Barriers CNS 2018, 15, 17. [Google Scholar] [CrossRef]
- Silver, I.A.; Erecińska, M. Extracellular Glucose Concentration in Mammalian Brain: Continuous Monitoring of Changes during Increased Neuronal Activity and upon Limitation in Oxygen Supply in Normo-, Hypo-, and Hyperglycemic Animals. J. Neurosci. Off. J. Soc. Neurosci. 1994, 14, 5068–5076. [Google Scholar] [CrossRef] [PubMed]
- Pinti, P.; Tachtsidis, I.; Hamilton, A.; Hirsch, J.; Aichelburg, C.; Gilbert, S.; Burgess, P.W. The Present and Future Use of Functional Near-Infrared Spectroscopy (fNIRS) for Cognitive Neuroscience. Ann. N. Y. Acad. Sci. 2020, 1464, 5–29. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Nam, K.I.; Song, J. The Glymphatic System in Diabetes-Induced Dementia. Front. Neurol. 2018, 9, 867. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Kang, H.; Xu, Q.; Chen, M.J.; Liao, Y.; Thiyagarajan, M.; O’Donnell, J.; Christensen, D.J.; Nicholson, C.; Iliff, J.J.; et al. Sleep Drives Metabolite Clearance from the Adult Brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Mukli, P.; Csipo, T.; Lipecz, A.; Stylianou, O.; Racz, F.S.; Owens, C.D.; Perry, J.W.; Tarantini, S.; Sorond, F.A.; Kellawan, J.M.; et al. Sleep Deprivation Alters Task-Related Changes in Functional Connectivity of the Frontal Cortex: A near-Infrared Spectroscopy Study. Brain Behav. 2021, 11, e02135. [Google Scholar] [CrossRef] [PubMed]
- Lanza, G.; DelRosso, L.M.; Ferri, R. Sleep and Homeostatic Control of Plasticity. Handb. Clin. Neurol. 2022, 184, 53–72. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, M.A.; Fricke, K.; Henschke, U.; Schlitterlau, A.; Liebetanz, D.; Lang, N.; Henning, S.; Tergau, F.; Paulus, W. Pharmacological Modulation of Cortical Excitability Shifts Induced by Transcranial Direct Current Stimulation in Humans. J. Physiol. 2003, 553, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilpour, Z.; Kronberg, G.; Reato, D.; Parra, L.C.; Bikson, M. Temporal Interference Stimulation Targets Deep Brain Regions by Modulating Neural Oscillations. Brain Stimulat. 2020, 14, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Doiron, B.; Goswami, C.; Grover, P. The Mechanics of Temporal Interference Stimulation. BioRxiv 2020. [Google Scholar] [CrossRef]
- Nishimura, N.; Schaffer, C.B.; Friedman, B.; Lyden, P.D.; Kleinfeld, D. Penetrating Arterioles Are a Bottleneck in the Perfusion of Neocortex. Proc. Natl. Acad. Sci. USA 2007, 104, 365–370. [Google Scholar] [CrossRef]
- Cheung, M.C. Hemodynamics Due to Calf Muscle Activity–Biophysical Modeling and Experiments Using Frequency Domain near Infrared Spectroscopy in Healthy Humans. Master’s Thesis, State University of New York at Buffalo, Buffalo, NY, USA, 2018. [Google Scholar]
- Battista, N.A.; Strickland, W.C.; Barrett, A.; Miller, L.A. IB2d Reloaded: A More Powerful Python and MATLAB Implementation of the Immersed Boundary Method. Math. Methods Appl. Sci. 2018, 41, 8455–8480. [Google Scholar] [CrossRef]
- Liebetanz, D.; Nitsche, M.A.; Tergau, F.; Paulus, W. Pharmacological Approach to the Mechanisms of Transcranial DC-Stimulation-Induced after-Effects of Human Motor Cortex Excitability. Brain J. Neurol. 2002, 125, 2238–2247. [Google Scholar] [CrossRef]
- Hariharan, A.; Robertson, C.D.; Garcia, D.C.G.; Longden, T.A. Brain Capillary Pericytes Are Metabolic Sentinels That Control Blood Flow through a KATP Channel-Dependent Energy Switch. Cell Rep. 2022, 41. [Google Scholar] [CrossRef] [PubMed]
- Arora, Y.; Chowdhury, S.R.; Dutta, A. Physiological Neurovascular Modeling of Cerebrovascular Effects of Transcranial Electrical Current Stimulation. Brain Stimul. Basic Transl. Clin. Res. Neuromodul. 2021, 14, 1597–1598. [Google Scholar] [CrossRef]
- Development of Point of Care Testing Device for Neurovascular Coupling From Simultaneous Recording of EEG and NIRS During Anodal Transcranial Direct Current Stimulation. IEEE J. Transl. Eng. Health Med. 2015, 3, 2000112. [CrossRef] [PubMed]
- Wiehler, A.; Branzoli, F.; Adanyeguh, I.; Mochel, F.; Pessiglione, M. A Neuro-Metabolic Account of Why Daylong Cognitive Work Alters the Control of Economic Decisions. Curr. Biol. 2022, 32, 3564–3575.e5. [Google Scholar] [CrossRef] [PubMed]
- Tarasoff-Conway, J.M.; Carare, R.O.; Osorio, R.S.; Glodzik, L.; Butler, T.; Fieremans, E.; Axel, L.; Rusinek, H.; Nicholson, C.; Zlokovic, B.V.; et al. Clearance Systems in the Brain—Implications for Alzheimer Disease. Nat. Rev. Neurol. 2015, 11, 457–470. [Google Scholar] [CrossRef] [PubMed]
- Tykocki, N.R.; Boerman, E.M.; Jackson, W.F. Smooth Muscle Ion Channels and Regulation of Vascular Tone in Resistance Arteries and Arterioles. Compr. Physiol. 2017, 7, 485–581. [Google Scholar] [CrossRef] [PubMed]
- Yassine, H.N.; Solomon, V.; Thakral, A.; Sheikh-Bahaei, N.; Chui, H.C.; Braskie, M.N.; Schneider, L.S.; Talbot, K. Brain Energy Failure in Dementia Syndromes: Opportunities and Challenges for Glucagon-like Peptide-1 Receptor Agonists. Alzheimers Dement. 2022, 18, 478–497. [Google Scholar] [CrossRef]
- Mallucci, G. Dementia Therapy: Time for an Energy Boost. Brain 2024, 147, 1593–1594. [Google Scholar] [CrossRef]
- Mukli, P.; Pinto, C.B.; Owens, C.D.; Csipo, T.; Lipecz, A.; Szarvas, Z.; Peterfi, A.; Langley, A.C.d.C.P.; Hoffmeister, J.; Racz, F.S.; et al. Impaired Neurovascular Coupling and Increased Functional Connectivity in the Frontal Cortex Predict Age-Related Cognitive Dysfunction. Adv. Sci. 2024, 11, 2303516. [Google Scholar] [CrossRef] [PubMed]
- Shahdadian, S.; Wang, X.; Liu, H. Directed Physiological Networks in the Human Prefrontal Cortex at Rest and Post Transcranial Photobiomodulation. Sci. Rep. 2024, 14, 10242. [Google Scholar] [CrossRef] [PubMed]
- Manippa, V.; Palmisano, A.; Filardi, M.; Vilella, D.; Nitsche, M.A.; Rivolta, D.; Logroscino, G. An Update on the Use of Gamma (Multi)Sensory Stimulation for Alzheimer’s Disease Treatment. Front. Aging Neurosci. 2022, 14, 1095081. [Google Scholar] [CrossRef] [PubMed]
- Kann, O.; Papageorgiou, I.E.; Draguhn, A. Highly Energized Inhibitory Interneurons Are a Central Element for Information Processing in Cortical Networks. J. Cereb. Blood Flow Metab. 2014, 34, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Beinlich, F.R.M.; Asiminas, A.; Untiet, V.; Bojarowska, Z.; Plá, V.; Sigurdsson, B.; Timmel, V.; Gehrig, L.; Graber, M.H.; Hirase, H.; et al. Oxygen Imaging of Hypoxic Pockets in the Mouse Cerebral Cortex. Science 2024, 383, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Dutta, A.; Zhao, F.; Cheung, M.; Das, A.; Tomita, M.; Chatterjee, K. Cerebral and Muscle Near-Infrared Spectroscopy during Lower-Limb Muscle Activity – Volitional and Neuromuscular Electrical Stimulation. In Proceedings of the 2021 43rd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Virtual, 1–5 November 2021; pp. 6577–6580. [Google Scholar]
- Guleyupoglu, B.; Schestatsky, P.; Edwards, D.; Fregni, F.; Bikson, M. Classification of Methods in Transcranial Electrical Stimulation (tES) and Evolving Strategy from Historical Approaches to Contemporary Innovations. J. Neurosci. Methods 2013, 219, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Paulus, W. Transcranial Electrical Stimulation (tES—tDCS; tRNS, tACS) Methods. Neuropsychol. Rehabil. 2011, 21, 602–617. [Google Scholar] [CrossRef]
- Rezaee, Z.; Dutta, A. Transcranial Direct Current Stimulation of the Leg Motor Area—Is It Partly Somatosensory? In Proceedings of the 2018 40th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC), Honolulu, HI, USA, 18–21 July 2018; pp. 4764–4767. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kozalakis, K.; Baftizadeh, F.; Campagnola, L.; Jarsky, T.; Koch, C.; Anastassiou, C.A. Cell-Class-Specific Electric Field Entrainment of Neural Activity. Neuron 2024, 0. [Google Scholar] [CrossRef]
- Dutta, A.; Dutta, A. Using Electromagnetic Reciprocity and Magnetic Resonance Current Density Imaging to Fit Multi-Electrode Montage for Non-Invasive Brain Stimulation. In Proceedings of the 2013 6th International IEEE/EMBS Conference on Neural Engineering (NER), San Diego, CA, USA, 6–8 November 2013; pp. 447–451. [Google Scholar]
- Antal, A.; Paulus, W. Transcranial Alternating Current Stimulation (tACS). Front. Hum. Neurosci. 2013, 7, 317. [Google Scholar] [CrossRef]
- Helfrich, R.F.; Schneider, T.R.; Rach, S.; Trautmann-Lengsfeld, S.A.; Engel, A.K.; Herrmann, C.S. Entrainment of Brain Oscillations by Transcranial Alternating Current Stimulation. Curr. Biol. 2014, 24, 333–339. [Google Scholar] [CrossRef]
- Machado, D.G.d.S.; Unal, G.; Andrade, S.M.; Moreira, A.; Altimari, L.R.; Brunoni, A.R.; Perrey, S.; Mauger, A.R.; Bikson, M.; Okano, A.H. Effect of Transcranial Direct Current Stimulation on Exercise Performance: A Systematic Review and Meta-Analysis. Brain Stimulat. 2019, 12, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Roberto, S.; Milia, R.; Doneddu, A.; Pinna, V.; Palazzolo, G.; Serra, S.; Orrù, A.; Hosseini Kakhak, S.A.; Ghiani, G.; Mulliri, G.; et al. Hemodynamic Abnormalities during Muscle Metaboreflex Activation in Patients with Type 2 Diabetes Mellitus. J. Appl. Physiol. Bethesda Md 1985 2019, 126, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Doneddu, A.; Roberto, S.; Pinna, V.; Magnani, S.; Ghiani, G.; Sainas, G.; Mulliri, G.; Serra, S.; Kakhak, S.A.H.; Milia, R.; et al. Effect of Combined Mental Task and Metaboreflex Activation on Hemodynamics and Cerebral Oxygenation in Patients With Metabolic Syndrome. Front. Physiol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Broggini, T.; Duckworth, J.; Ji, X.; Liu, R.; Xia, X.; Mächler, P.; Shaked, I.; Munting, L.P.; Iyengar, S.; Kotlikoff, M.; et al. Long-Wavelength Traveling Waves of Vasomotion Modulate the Perfusion of Cortex. Neuron 2024. ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, T.; Tamura, Y. A Frequency–Spatial Domain Decomposition (FSDD) Method for Operational Modal Analysis. Mech. Syst. Signal Process. 2010, 24, 1227–1239. [Google Scholar] [CrossRef]
- Kutz, J.N.; Brunton, S.L.; Brunton, B.W.; Proctor, J.L. Dynamic Mode Decomposition; Other Titles in Applied Mathematics; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 2016; ISBN 978-1-61197-449-2. [Google Scholar]
- Brunton, S.L.; Kutz, J.N. Data-Driven Science and Engineering: Machine Learning, Dynamical Systems, and Control; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Kamdem, C.D.B.; Yomi, P.A.N.; Tabi, C.B.; Mohamadou, A. Modulated Blood Waves in the Coupled Complex Ginzburg–Landau Equations of Jeffrey Fluids in Arteries. Eur. Phys. J. Plus 2023, 138, 176. [Google Scholar] [CrossRef]
- Cole, J.D. Perturbation Methods in Fluid Mechanics (Milton Van Dyke). SIAM Rev. 1977, 19, 572. [Google Scholar] [CrossRef]
- Fraenkel, L.E. Perturbation Methods in Fluid Mechanics. By MILTON VAN DYKE. Academic Press, 1964. 229 Pp. 22 16s. J. Fluid Mech. 1965, 23, 822–825. [Google Scholar] [CrossRef]
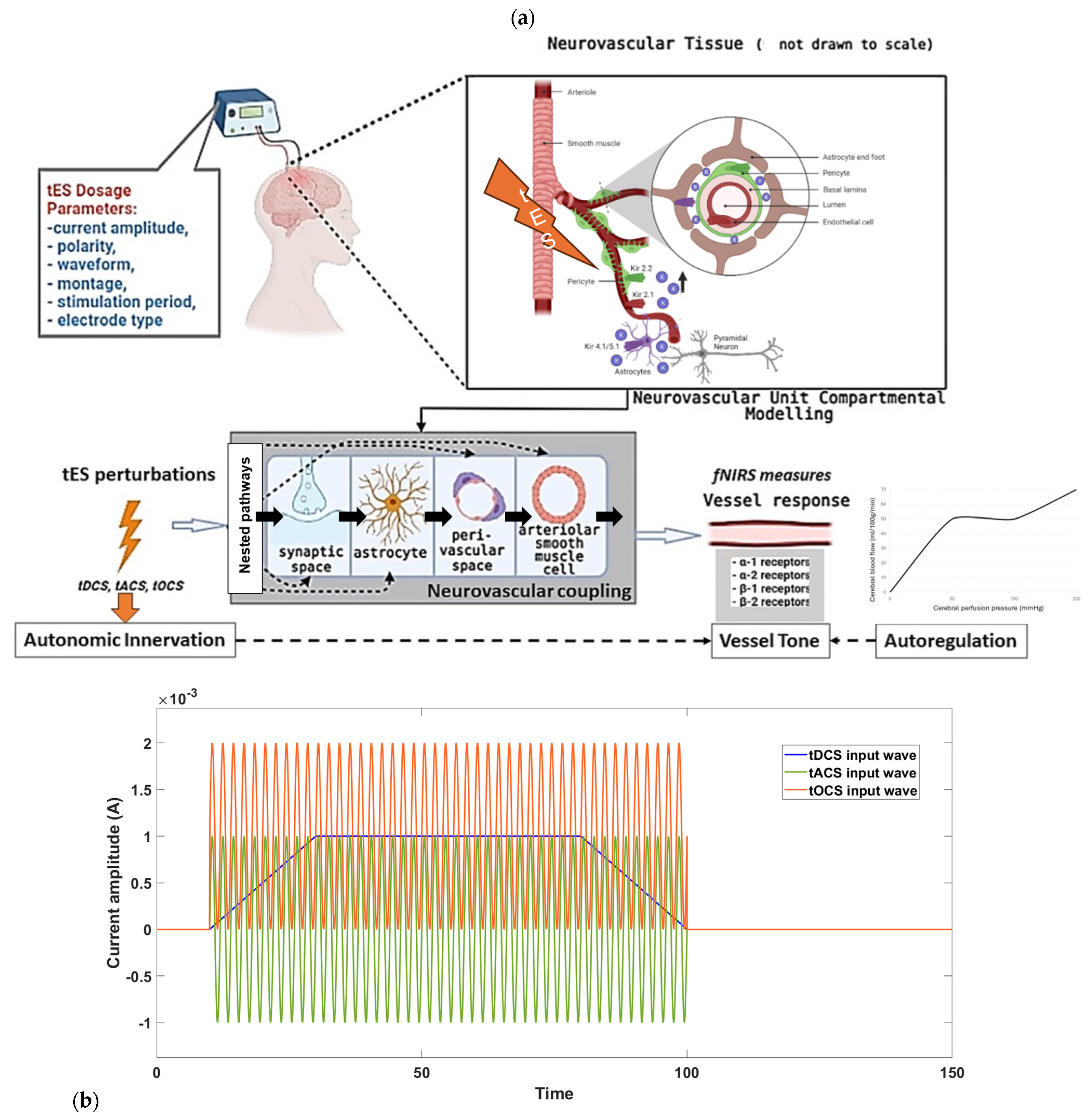
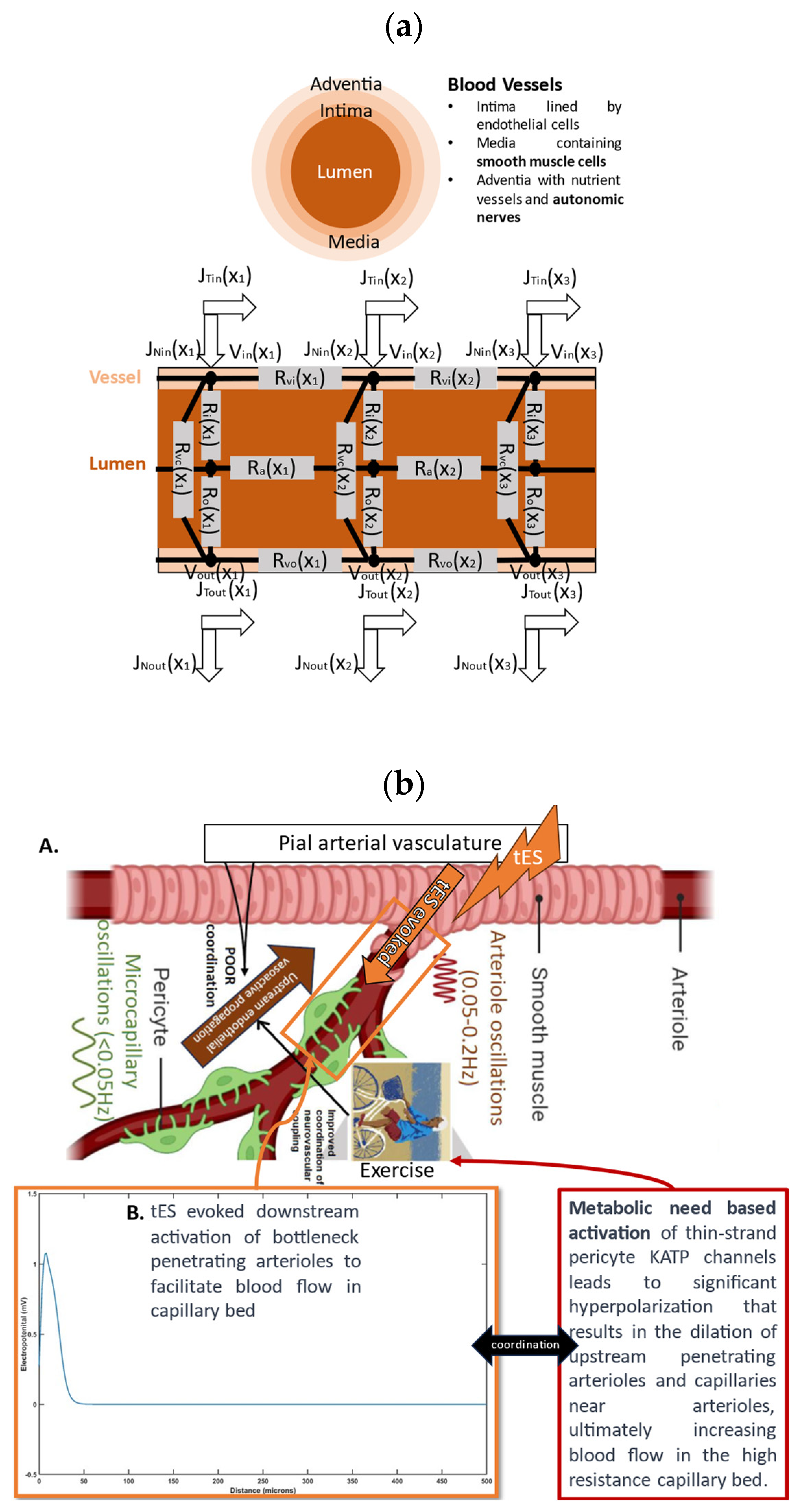

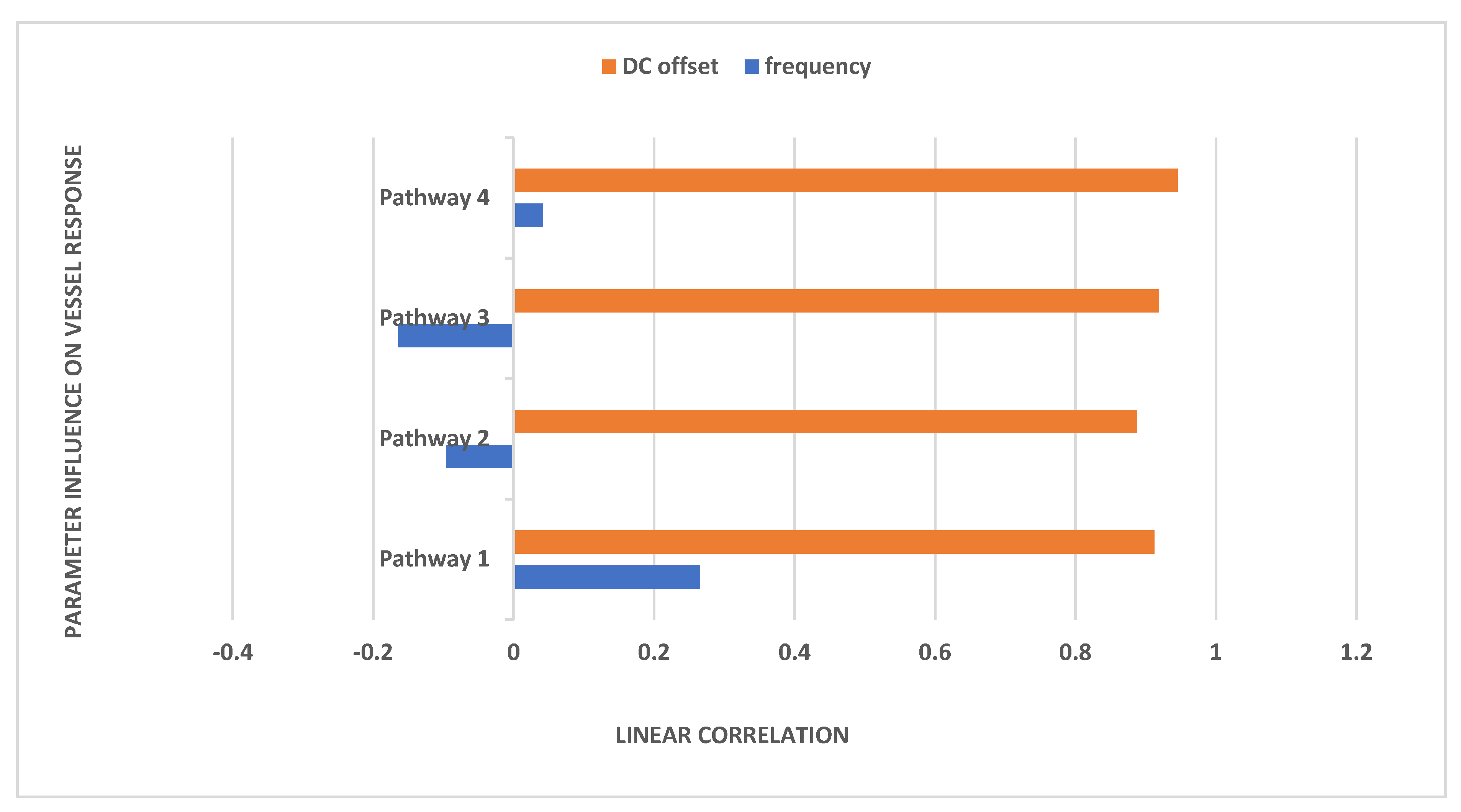



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway/Bandpass-Filtered White Noise Input | P1 (K1 = 0.000001) Hz | P2 (K2 = 0.00000000001) Hz | P3 (K3 = 0.001) Hz | P4 (K4 = 0.000000000001) Hz |
|---|---|---|---|---|
| 1* | 0.0193 | 0.0247 | 0.0052, 0.0157, 0.0265 | 0.0069, 0.0201 |
| 2 | 0.0189, 0.0201 | 0.0003, 0.0156, 0.0167 | 0.0025, 0.0231, 0.0235 | 0.0128, 0.0141 |
| 3 | 0.0153, 0.0195 | 0.0109, 0.0191, 0.0288, 0.0476 | 0.0027, 0.0081, 0.0196, 0.0236 | 0.0067, 0.0090, 0.0144 |
| 4 | 0.0115, 0.0245 | - | 0.0050 | 0.0020, 0.0168 |
| 5 | 0.0007, 0.0077, 0.0200, 0.0276 | 0.0131 | 0.0005, 0.0128, 0.0176, | 0.0192, 0.0318 |
| 6 | 0.0179, 0.0363 | 0.0237, 0.0418 | 0.0055 | 0.0104, 0.0171 |
| 7 | 0.0107, 0.0131 | 0.0128, 0.0200, 0.0380, 0.0659 | 0.0036, 0.0386 | 0.0381 |
| 8 | 0.0225, 0.0298 | 0.0048, 0.0100 | 0.0011, 0.0209, | 0.0043, 0.0223 |
| 9 | 0.0215 | 0.0038, 0.0285 | 0.0033, 0.0083, 0.0184 | 0.0099, 0.0164 |
| 10 | 0.0198, 0.0319, 0.0360 | 0.0189, 0.0229, 0.0242 | 0.0058, 0.0188 | 0.0387 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefanski, M.; Arora, Y.; Cheung, M.; Dutta, A. Modal Analysis of Cerebrovascular Effects for Digital Health Integration of Neurostimulation Therapies—A Review of Technology Concepts. Brain Sci. 2024, 14, 591. https://doi.org/10.3390/brainsci14060591
Stefanski M, Arora Y, Cheung M, Dutta A. Modal Analysis of Cerebrovascular Effects for Digital Health Integration of Neurostimulation Therapies—A Review of Technology Concepts. Brain Sciences. 2024; 14(6):591. https://doi.org/10.3390/brainsci14060591
Chicago/Turabian StyleStefanski, Marcel, Yashika Arora, Mancheung Cheung, and Anirban Dutta. 2024. "Modal Analysis of Cerebrovascular Effects for Digital Health Integration of Neurostimulation Therapies—A Review of Technology Concepts" Brain Sciences 14, no. 6: 591. https://doi.org/10.3390/brainsci14060591
APA StyleStefanski, M., Arora, Y., Cheung, M., & Dutta, A. (2024). Modal Analysis of Cerebrovascular Effects for Digital Health Integration of Neurostimulation Therapies—A Review of Technology Concepts. Brain Sciences, 14(6), 591. https://doi.org/10.3390/brainsci14060591