
Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Sample Collection and Laboratory Analysis
2.3. SNP Selection and Genotyping
2.4. Statistical Analysis
3. Results
3.1. Characteristics of Obese and Non-Obese Cohorts
3.2. Allelic Discrimination Analysis in Obese and Non-Obese Subjects
3.3. Monogenic Risk of Obesity with Each Polymorphism
3.4. Polygenic Risk of Obesity in Association with Gene Variants
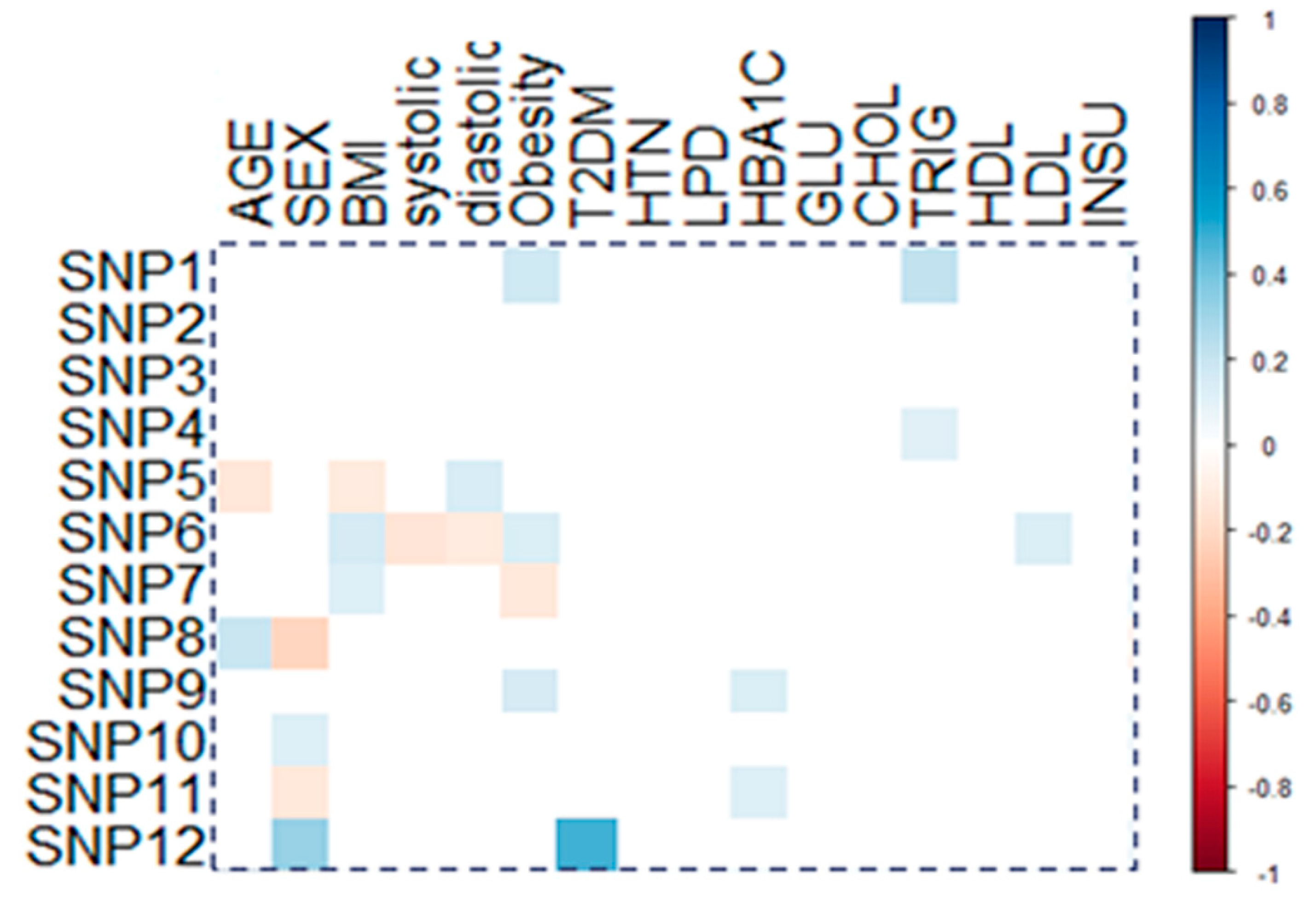
3.5. Association of Gene Variants with Clinical and Biochemical Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dai, H.; Alsalhe, T.A.; Chalghaf, N.; Riccò, M.; Bragazzi, N.L.; Wu, J. The Global Burden of Disease Attributable to High Body Mass Index in 195 Countries and Territories, 1990-2017: An Analysis of the Global Burden of Disease Study. PLoS Med. 2020, 17, e1003198. [Google Scholar] [CrossRef]
- Williams, E.P.; Mesidor, M.; Winters, K.; Dubbert, P.M.; Wyatt, S.B. Overweight and Obesity: Prevalence, Consequences, and Causes of a Growing Public Health Problem. Curr. Obes. Rep. 2015, 4, 363–370. [Google Scholar] [CrossRef]
- Sherif, S.; Sumpio, B.E. Economic Development and Diabetes Prevalence in MENA Countries: Egypt and Saudi Arabia Comparison. World J. Diabetes 2015, 6, 304–311. [Google Scholar] [CrossRef]
- Al-Ghamdi, S.; Shubair, M.M.; Aldiab, A.; Al-Zahrani, J.M.; Aldossari, K.K.; Househ, M.; Nooruddin, S.; Razzak, H.A.; El-Metwally, A. Prevalence of Overweight and Obesity Based on the Body Mass Index; a Cross-Sectional Study in Alkharj, Saudi Arabia. Lipids Health Dis. 2018, 17, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzeh, F.S.; Bukhari, H.M.; Header, E.A.; Ghabashi, M.A.; Al-Mashi, S.S.; Noorwali, N.M. Trends in Overweight or Obesity and other Anthropometric Indices in Adults Aged 18-60 Years in Western Saudi Arabia. Ann. Saudi. Med. 2017, 37, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawzy, M.S.; AlSel, B.T.A. Association of Serum Uric Acid Levels with Components of Metabolic Syndrome: A Cross-Sectional Analysis in a Saudi Adult Population. Int. J. Biomed. 2020, 10, 457–466. [Google Scholar] [CrossRef]
- Fawzy, M.S.; Alhadramy, O.; Hussein, M.H.; Ismail, H.M.; Ismail, N.M.; Biomy, N.M.; Toraih, E.A. Functional and Structural Impact of ATP-Binding Cassette Transporter A1 R219K and I883M Gene Polymorphisms in Obese Children and Adolescents. Mol. Diagn. Ther. 2015, 19, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [Green Version]
- Vincent, H.K.; Taylor, A.G. Biomarkers and Potential Mechanisms of Obesity-Induced Oxidant Stress in Humans. Int. J. Obes. 2006, 30, 400–418. [Google Scholar] [CrossRef] [Green Version]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased Oxidative Stress in Obesity and its Impact on Metabolic Syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Lee, H.; Lee, Y.J.; Choi, H.; Ko, E.H.; Kim, J.W. Reactive Oxygen Species Facilitate Adipocyte Differentiation by Accelerating Mitotic Clonal Expansion. J. Biol. Chem. 2009, 284, 10601–10609. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, M.; Dusting, G.J.; Peshavariya, H.; Jiang, F.; Hsiao, S.T.; Chan, E.C.; Liu, G.S. Differentiation of Human Adipose-Derived Stem Cells Into Fat Involves Reactive Oxygen Species and Forkhead box O1 Mediated Upregulation of Antioxidant Enzymes. Stem Cells Dev. 2013, 22, 878–888. [Google Scholar] [CrossRef] [Green Version]
- Horvath, T.L.; Andrews, Z.B.; Diano, S. Fuel Utilization by Hypothalamic Neurons: Roles for ROS. Trends Endocrinol. Metab. 2009, 20, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Savini, I.; Catani, M.V.; Evangelista, D.; Gasperi, V.; Avigliano, L. Obesity-Associated Oxidative Stress: Strategies Finalized to Improve Redox State. Int. J. Mol. Sci 2013, 14, 10497–10538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čolak, E.; Pap, D. The Role of Oxidative Stress in the Development of Obesity and Obesity-Related Metabolic Disorders. J. Med. Biochem. 2021, 40, 1–9. [Google Scholar] [CrossRef]
- Picklo, M.J.; Long, E.K.; Vomhof-DeKrey, E.E. Glutathionyl Systems and Metabolic Dysfunction in Obesity. Nutr. Rev. 2015, 73, 858–868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozata, M.; Mergen, M.; Oktenli, C.; Aydin, A.; Sanisoglu, S.Y.; Bolu, E.; Yilmaz, M.I.; Sayal, A.; Isimer, A.; Ozdemir, I.C. Increased Oxidative Stress and Hypozincemia in Male Obesity. Clin. Biochem. 2002, 35, 627–631. [Google Scholar] [CrossRef]
- Olusi, S.O. Obesity is an Independent Risk Factor for Plasma Lipid Peroxidation and Depletion of Erythrocyte Cytoprotectic Enzymes in Humans. Int. J. Obes. Relat. Metab. Disord. 2002, 26, 1159–1164. [Google Scholar] [CrossRef] [Green Version]
- Bełtowski, J.; Wójcicka, G.; Górny, D.; Marciniak, A. The Effect of Dietary-Induced Obesity on Lipid Peroxidation, Antioxidant Enzymes and Total Plasma Antioxidant Capacity. J. Physiol. Pharmacol. 2000, 51, 883–896. [Google Scholar]
- Lee, Y.S.; Kim, A.Y.; Choi, J.W.; Kim, M.; Yasue, S.; Son, H.J.; Masuzaki, H.; Park, K.S.; Kim, J.B. Dysregulation of Adipose Glutathione Peroxidase 3 in Obesity Contributes to Local and Systemic Oxidative Stress. Mol. Endocrinol. 2008, 22, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Deignan, J.L.; Qi, H.; Zhu, J.; Qian, S.; Zhong, J.; Torosyan, G.; Majid, S.; Falkard, B.; Kleinhanz, R.R.; et al. Validation of Candidate Causal Genes for Obesity that Affect Shared Metabolic Pathways and Networks. Nat. Genet. 2009, 41, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baez-Duarte, B.G.; Zamora-Ginez, I.; Mendoza-Carrera, F.; Ruiz-Vivanco, G.; Torres-Rasgado, E.; Gonzalez-Mejia, M.E.; Garcia-Zapien, A.; Flores-Martinez, S.E.; Perez-Fuentes, R. Serum Levels of Glutathione Peroxidase 3 in Overweight and Obese Subjects from Central Mexico. Arch. Med. Res. 2012, 43, 541–547. [Google Scholar] [CrossRef]
- Guerra, R.C.; Zuñiga-Muñoz, A.; Guarner Lans, V.; Díaz-Díaz, E.; Tena Betancourt, C.A.; Pérez-Torres, I. Modulation of the Activities of Catalase, cu-zn, mn Superoxide Dismutase, and Glutathione Peroxidase in Adipocyte from Ovariectomised Female Rats with Metabolic Syndrome. Int. J. Endocrinol. 2014, 2014, 175080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elizalde, M.; Rydén, M.; van Harmelen, V.; Eneroth, P.; Gyllenhammar, H.; Holm, C.; Ramel, S.; Olund, A.; Arner, P.; Andersson, K. Expression of Nitric Oxide Synthases in Subcutaneous Adipose Tissue of Non-Obese and Obese Humans. J. Lipid Res. 2000, 41, 1244–1251. [Google Scholar] [CrossRef]
- Rydén, M.; Elizalde, M.; van Harmelen, V.; Ohlund, A.; Hoffstedt, J.; Bringman, S.; Andersson, K. Increased Expression of eNOS Protein in Omental Versus Subcutaneous Adipose Tissue in Obese Human Subjects. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 811–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvin, V.B.; Barakat, H.; Kemeny, G.; Macdonald, K.G.; Pories, W.J.; Hickner, R.C. Endothelial Nitric Oxide Synthase Content in Adipose Tissue from Obese and Lean African American and white American Women. Metabolism 2005, 54, 1368–1373. [Google Scholar] [CrossRef] [PubMed]
- Probst-Hensch, N.M.; Imboden, M.; Felber Dietrich, D.; Barthélemy, J.C.; Ackermann-Liebrich, U.; Berger, W.; Gaspoz, J.M.; Schwartz, J. Glutathione S-Transferase Polymorphisms, Passive Smoking, Obesity, and Heart Rate Variability in Nonsmokers. Environ. Health Perspect. 2008, 116, 1494–1499. [Google Scholar] [CrossRef] [Green Version]
- Afrand, M.; Bashardoost, N.; Sheikhha, M.H.; Afkhami-Ardekani, M. Association Between Glutathione S-Transferase GSTM1-T1 and P1 Polymorphisms with Metabolic Syndrome in Zoroastrians in Yazd, Iran. Iran. J. Public Health 2015, 44, 673–682. [Google Scholar]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. The Copper-Zinc Superoxide Dismutase Activity in Selected Diseases. Eur. J. Clin. Investig. 2019, 49, e13036. [Google Scholar] [CrossRef] [Green Version]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide Dismutase Multigene Family: A Comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) Gene Structures, Evolution, and Expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Perry, J.J.; Shin, D.S.; Getzoff, E.D.; Tainer, J.A. The Structural Biochemistry of the Superoxide Dismutases. Biochim. Biophys Acta 2010, 1804, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Kinnula, V.L.; Lehtonen, S.; Koistinen, P.; Kakko, S.; Savolainen, M.; Kere, J.; Ollikainen, V.; Laitinen, T. Two Functional Variants of the Superoxide Dismutase Genes in Finnish Families with Asthma. Thorax 2004, 59, 116–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Alterations in Concentration/Activity of Superoxide Dismutases in Context of Obesity and Selected Single Nucleotide Polymorphisms in Genes. Int. J. Mol. Sci. 2020, 21, 5069. [Google Scholar] [CrossRef]
- Hernández-Guerrero, C.; Hernández-Chávez, P.; Romo-Palafox, I.; Blanco-Melo, G.; Parra-Carriedo, A.; Pérez-Lizaur, A. Genetic Polymorphisms in SOD (rs2070424, rs7880) and CAT (rs7943316, rs1001179) Enzymes Are Associated with Increased Body Fat Percentage and Visceral Fat in an Obese Population from Central Mexico. Arch. Med. Res. 2016, 47, 331–339. [Google Scholar] [CrossRef]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione Peroxidases. Biochim. Biophys. Acta 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Ghasemi, H.; Roustazadeh, A.; Alipoor, B. Phenotype and Genotype Relationship of Glutathione Peroxidase1 (GPx1) and rs 1800668 Variant: The Homozygote Effect on Kinetic Parameters. Gene 2012, 505, 19–22. [Google Scholar] [CrossRef]
- Perez, R.V.; Machado, C.G.; Santos-Bezerra, D.P.; Admoni, S.N.; Patente, T.A.; Monteiro, M.B.; Cavaleiro, A.M.; Queiroz, M.S.; Nery, M.; Corrêa-Giannella, M.L. Allelic Variations in Genes Belonging to Glutathione System Increase Proliferative Retinopathy Risk in Type 1 Diabetes Individuals. Gene 2019, 703, 120–124. [Google Scholar] [CrossRef]
- Admoni, S.N.; Santos-Bezerra, D.P.; Perez, R.V.; Patente, T.A.; Monteiro, M.B.; Cavaleiro, A.M.; Parisi, M.C.; Moura Neto, A.; Pavin, E.J.; Queiroz, M.S.; et al. Glutathione Peroxidase 4 Functional Variant rs713041 Modulates the Risk for Cardiovascular Autonomic Neuropathy in Individuals with Type 1 Diabetes. Diab. Vasc. Dis. Res. 2019, 16, 297–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sansbury, B.E.; Hill, B.G. Regulation of Obesity and Insulin Resistance by Nitric Oxide. Free Radic. Biol. Med. 2014, 73, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Joost, H.-G.; Tschop, M.H. NO to Obesity: Does Nitric Oxide Regulate Fat Oxidation and Insulin Sensitivity? Endocrinology 2007, 148, 4545–4547. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Li, Y.M.; Yang, L.Q.; Zhong, C.G.; Zhuang, Z.X. Association of NOS2 and NOS3 Gene Polymorphisms with Susceptibility to Type 2 Diabetes Mellitus and Diabetic Nephropathy in the Chinese Han Population. IUBMB Life 2016, 68, 516–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasr, H.B.; Dimassi, S.; M’hadhbi, R.; Debbabi, H.; Kortas, M.; Tabka, Z.; Chahed, K. Functional G894T (rs1799983) Polymorphism and Intron-4 VNTR Variant of Nitric Oxide Synthase (NOS3) Gene are Susceptibility Biomarkers of Obesity Among Tunisians. Obes. Res. Clin. Pract. 2016, 10, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Pawlik, A.; Błaszczyk, H.; Rać, M.; Maciejewska-Skrendo, A.; Safranow, K.; Dziedziejko, V. NOS3 Gene rs1799983 and rs2070744 Polymorphisms in Patients with Unstable Angina. J. Vasc. Res. 2020, 57, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2018, 71, e127–e248. [Google Scholar] [CrossRef] [PubMed]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the Concentration of Low-Density Lipoprotein Cholesterol in Plasma, Without Use of the Preparative Ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Health, N.I.O. ATP III Guidelines at-a-Glance Quick Desk Reference; NIH Publication: Bethesda, MD, USA, 2001; pp. 1–3305. [Google Scholar]
- Fawzy, M.S.; Ibrahiem, A.T.; AlSel, B.T.A.; Alghamdi, S.A.; Toraih, E.A. Analysis of Microrna-34a Expression Profile and rs2666433 Variant in Colorectal Cancer: A Pilot Study. Sci. Rep. 2020, 10, 16940. [Google Scholar] [CrossRef] [PubMed]
- Janssens, A.C.; Moonesinghe, R.; Yang, Q.; Steyerberg, E.W.; van Duijn, C.M.; Khoury, M.J. The Impact of Genotype Frequencies on the Clinical Validity of Genomic Profiling for Predicting Common Chronic Diseases. Genet. Med. 2007, 9, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haschenburger, J.K.; Spinelli, J.J. Assessing the Goodness-of-Fit of Statistical Distributions when Data are Grouped. Math. Geol. 2005, 37, 261–276. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A Web Tool for the Analysis of Association Studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [Green Version]
- Horita, N.; Kaneko, T. Genetic Model Selection for a Case–Control Study and a Meta-Analysis. Meta. Gene. 2015, 5, 1–8. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package’ Corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- Găman, M.A.; Epîngeac, M.E.; Diaconu, C.C.; Găman, A.M. Evaluation of Oxidative Stress Levels in Obesity and Diabetes by the Free Oxygen Radical Test and Free Oxygen Radical Defence Assays and Correlations with Anthropometric and Laboratory Parameters. World J. Diabetes 2020, 11, 193–201. [Google Scholar] [CrossRef]
- Villareal, D.T.; Apovian, C.M.; Kushner, R.F.; Klein, S.; Nutrition, A.S.F.; NAASO, T.E.O.S. Obesity in Older Adults: Technical Review and Position Statement of the American Society for Nutrition and NAASO, The Obesity Society. Am. J. Clin. Nutr. 2005, 82, 923–934. [Google Scholar] [CrossRef]
- Martyn, J.A.; Kaneki, M.; Yasuhara, S. Obesity-Induced Insulin Resistance and Hyperglycemia: Etiologic Factors and Molecular Mechanisms. Anesthesiology 2008, 109, 137–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klop, B.; Elte, J.W.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218–1240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.K.; Reaven, G.M.; Kim, S.H. Dissecting the Relationship Between Obesity and Hyperinsulinemia: Role of Insulin Secretion and Insulin Clearance. Obesity (Silver Spring) 2017, 25, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Leggio, M.; Lombardi, M.; Caldarone, E.; Severi, P.; D’Emidio, S.; Armeni, M.; Bravi, V.; Bendini, M.G.; Mazza, A. The Relationship Between Obesity and Hypertension: An Updated Comprehensive Overview on Vicious Twins. Hypertens Res. 2017, 40, 947–963. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.; Mooney, K.; Mc Auley, M. Obesity and the Dysregulation of Fatty Acid Metabolism: Implications for Healthy Aging. Expert Rev. Endocrinol. Metab. 2016, 11, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tareen, S.H.K.; Adriaens, M.E.; Arts, I.C.W.; de Kok, T.M.; Vink, R.G.; Roumans, N.J.T.; van Baak, M.A.; Mariman, E.C.M.; Evelo, C.T.; Kutmon, M. Profiling Cellular Processes in Adipose Tissue during Weight Loss Using Time Series Gene Expression. Genes 2018, 9, 525. [Google Scholar] [CrossRef] [Green Version]
- Le Lay, S.; Simard, G.; Martinez, M.C.; Andriantsitohaina, R. Oxidative Stress and Metabolic Pathologies: From an Adipocentric Point of View. Oxid Med. Cell Longev. 2014, 2014, 908539. [Google Scholar] [CrossRef] [Green Version]
- Vávrová, L.; Kodydková, J.; Zeman, M.; Dušejovská, M.; Macášek, J.; Staňková, B.; Tvrzická, E.; Zák, A. Altered Activities of Antioxidant Enzymes in Patients with Metabolic Syndrome. Obes. Facts 2013, 6, 39–47. [Google Scholar] [CrossRef]
- Mattson, M.P. Roles of the Lipid Peroxidation Product 4-Hydroxynonenal in Obesity, the Metabolic Syndrome, and Associated Vascular and Neurodegenerative Disorders. Exp. Gerontol. 2009, 44, 625–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.A. Lack of Association Between Glutathione s-Transferase Mu 1 (GSTM1) Gene Polymorphisms and Obesity. J. Exerc. Rehabil. 2017, 13, 608–612. [Google Scholar] [CrossRef] [Green Version]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the Contribution of Synonymous Mutations to Human Disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Flekac, M.; Skrha, J.; Hilgertova, J.; Lacinova, Z.; Jarolimkova, M. Gene Polymorphisms of Superoxide Dismutases and Catalase in Diabetes Mellitus. BMC Med. Genet. 2008, 9, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montano, M.A.; Barrio Lera, J.P.; Gottlieb, M.G.; Schwanke, C.H.; da Rocha, M.I.; Manica-Cattani, M.F.; dos Santos, G.F.; da Cruz, I.B. Association Between Manganese Superoxide Dismutase (MnSOD) Gene Polymorphism and Elderly Obesity. Mol. Cell Biochem. 2009, 328, 33–40. [Google Scholar] [CrossRef]
- Echtay, K.S.; Murphy, M.P.; Smith, R.A.; Talbot, D.A.; Brand, M.D. Superoxide Activates Mitochondrial Uncoupling Protein 2 from the Matrix side. Studies Using Targeted Antioxidants. J. Biol. Chem. 2002, 277, 47129–47135. [Google Scholar] [CrossRef] [Green Version]
- Sutton, A.; Imbert, A.; Igoudjil, A.; Descatoire, V.; Cazanave, S.; Pessayre, D.; Degoul, F. The Manganese Superoxide Dismutase Ala16Val Dimorphism Modulates Both Mitochondrial Import and mRNA Stability. Pharm. Genom. 2005, 15, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Becer, E.; Çırakoğlu, A. Association of the Ala16Val MnSOD Gene Polymorphism with Plasma Leptin Levels and Oxidative Stress Biomarkers in Obese Patients. Gene 2015, 568, 35–39. [Google Scholar] [CrossRef]
- Decharatchakul, N.; Settasatian, C.; Settasatian, N.; Komanasin, N.; Kukongviriyapan, U.; Intharaphet, P.; Senthong, V. Association of Genetic Polymorphisms in SOD2, SOD3, GPX3, and GSTT1 with Hypertriglyceridemia and Low HDL-C Level in Subjects with High Risk of Coronary Artery Disease. PeerJ 2019, 7, e7407. [Google Scholar] [CrossRef] [Green Version]
- Masschelin, P.M.; Cox, A.R.; Chernis, N.; Hartig, S.M. The Impact of Oxidative Stress on Adipose Tissue Energy Balance. Front. Physiol. 2019, 10, 1638. [Google Scholar] [CrossRef]
- Han, Y.H.; Buffolo, M.; Pires, K.M.; Pei, S.; Scherer, P.E.; Boudina, S. Adipocyte-Specific Deletion of Manganese Superoxide Dismutase Protects from Diet-Induced Obesity Through Increased Mitochondrial Uncoupling and Biogenesis. Diabetes 2016, 65, 2639–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, X.; Li, D.; Liu, H.; Zhao, Y. SOD3 and eNOS Genotypes are Associated with SOD Activity and NO. Exp. Ther. Med. 2014, 8, 328–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.M.; Xie, X.R.; Jin, A.L. Genetic Polymorphisms in Extracellular Superoxide Dismutase Leu53Leu, Arg213Gly, and Ala40Thr and Susceptibility to Type 2 Diabetes Mellitus. Genet. Mol. Res. 2016, 15. [Google Scholar] [CrossRef]
- Stancill, J.S.; Happ, J.T.; Broniowska, K.A.; Hogg, N.; Corbett, J.A. Peroxiredoxin 1 Plays a Primary Role in Protecting Pancreatic β-Cells from Hydrogen Peroxide and Peroxynitrite. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020, 318, R1004–R1013. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, M.; Hong, K.; Cope, E.L.; Daneva, Z.; Johnstone, S.R.; Isakson, B.; Sonkusare, S.K. Locally formed Peroxynitrite Inhibits Endothelial TRPV4 Channels and Elevates Blood Pressure in Obesity. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Cui, R.; Gao, M.; Qu, S.; Liu, D. Overexpression of Superoxide Dismutase 3 Gene Blocks High-Fat Diet-Induced Obesity, Fatty Liver and Insulin Resistance. Gene Ther. 2014, 21, 840–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irfan, S.; Rani, A.; Sameem, M.; Nawaz, S.K.; Liaqat, I.; Arshad, M. Association of rs1800668 Polymorphism in Glutathione Peroxidase- 1 Gene and Risk of Rheumatoid Arthritis in Pakistani Population. Pak. J. Med. Sci. 2016, 32, 1204–1207. [Google Scholar] [CrossRef]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M.; et al. Reactive Oxygen Species Enhance Insulin Sensitivity. Cell Metab. 2009, 10, 260–272. [Google Scholar] [CrossRef] [Green Version]
- Merry, T.L.; Tran, M.; Stathopoulos, M.; Wiede, F.; Fam, B.C.; Dodd, G.T.; Clarke, I.; Watt, M.J.; Andrikopoulos, S.; Tiganis, T. High-Fat-Fed Obese Glutathione Peroxidase 1-Deficient Mice Exhibit Defective Insulin Secretion but Protection from Hepatic Steatosis and Liver Damage. Antioxid. Redox Signal. 2014, 20, 2114–2129. [Google Scholar] [CrossRef]
- Dosenko, V.E.; Zagoriy, V.Y.; Haytovich, N.V.; Gordok, O.A.; Moibenko, A.A. Allelic Polymorphism of Endothelial NO-Synthase Gene and its Functional Manifestations. Acta Biochim. Pol. 2006, 53, 299–302. [Google Scholar] [CrossRef] [PubMed]
- Wiltshire, E.J.; Peña, A.S.; MacKenzie, K.; Bose-Sundernathan, T.; Gent, R.; Couper, J.J. A NOS3 Polymorphism Determines Endothelial Response to Folate in Children with Type 1 Diabetes or Obesity. J. Pediatr. 2015, 166, 319–325.e311. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, S.; Poole, J.; Davis, H.; Dong, Y.; Pollock, J.; Snieder, H.; Treiber, F. Effects of NOS3 Glu298Asp Polymorphism on Hemodynamic Reactivity to Stress: Influences of Ethnicity and Obesity. Hypertension 2004, 44, 866–871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bressler, J.; Pankow, J.S.; Coresh, J.; Boerwinkle, E. Interaction Between the NOS3 Gene and Obesity as a Determinant of Risk of Type 2 Diabetes: The Atherosclerosis Risk in Communities Study. PLoS ONE 2013, 8, e79466. [Google Scholar] [CrossRef] [Green Version]
- Tanus-Santos, J.E.; Desai, M.; Flockhart, D.A. Effects of Ethnicity on the Distribution of Clinically Relevant Endothelial Nitric Oxide Variants. Pharmacogenetics 2001, 11, 719–725. [Google Scholar] [CrossRef]
- Marroni, A.S.; Metzger, I.F.; Souza-Costa, D.C.; Nagassaki, S.; Sandrim, V.C.; Correa, R.X.; Rios-Santos, F.; Tanus-Santos, J.E. Consistent Interethnic Differences in the Distribution of Clinically Relevant Endothelial Nitric Oxide Synthase Genetic Polymorphisms. Nitric. Oxide 2005, 12, 177–182. [Google Scholar] [CrossRef]
- Dhillon, S.S.; Mastropaolo, L.A.; Murchie, R.; Griffiths, C.; Thöni, C.; Elkadri, A.; Xu, W.; Mack, A.; Walters, T.; Guo, C.; et al. Higher Activity of the Inducible Nitric Oxide Synthase Contributes to Very Early Onset Inflammatory Bowel Disease. Clin. Transl. Gastroenterol. 2014, 5, e46. [Google Scholar] [CrossRef]
- Qidwai, T.; Jamal, F. Inducible Nitric Oxide Synthase (iNOS) Gene Polymorphism and Disease Prevalence. Scand. J. Immunol. 2010, 72, 375–387. [Google Scholar] [CrossRef]
- Gusti, A.M.T.; Qusti, S.Y.; Bahijri, S.M.; Toraih, E.A.; Bokhari, S.; Attallah, S.M.; Alzahrani, A.; Alshehri, W.M.A.; Alotaibi, H.; Fawzy, M.S. Glutathione S-Transferase (GSTT1 rs17856199) and Nitric Oxide Synthase (NOS2 rs2297518) Genotype Combination as Potential Oxidative Stress-Related Molecular Markers for Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. 2021, 14, 1385–1403. [Google Scholar] [CrossRef]
- Porojan, M.D.; Cătană, A.; Popp, R.A.; Dumitrascu, D.L.; Bala, C. The Role of NOS2A -954G/C and Vascular Endothelial Growth Factor +936C/T Polymorphisms in Type 2 Diabetes Mellitus and Diabetic Nonproliferative Retinopathy Risk Management. Ther. Clin. Risk Manag. 2015, 11, 1743–1748. [Google Scholar] [CrossRef] [Green Version]
- Shahid, S.U.; Hasnain, S. Use of a Gene Score of Multiple Low-Modest Effect Size Variants can Predict the Risk of Obesity Better than the Individual SNPs. Lipids Health Dis. 2018, 17, 155. [Google Scholar] [CrossRef] [Green Version]
- Hernández-Guerrero, C.; Parra-Carriedo, A.; Ruiz-de-Santiago, D.; Galicia-Castillo, O.; Buenrostro-Jáuregui, M.; Díaz-Gutiérrez, C. Genetic Polymorphisms of Antioxidant Enzymes CAT and SOD Affect the Outcome of Clinical, Biochemical, and Anthropometric Variables in People with Obesity Under a Dietary Intervention. Genes Nutr. 2018, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.Y.; Mitra, S.R. The Combined Effect of Polygenic Risk from FTO and ADRB2 Gene Variants, Odds of Obesity, and Post-Hipcref Diet Differences. Lifestyle Genom. 2020, 13, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Llanaj, E.; Pikó, P.; Nagy, K.; Rácz, G.; János, S.; Kósa, Z.; Fiatal, S.; Ádány, R. Applicability of Obesity-Related SNPs and Their Effect Size Measures Defined on Populations with European Ancestry for Genetic Risk Estimation Among Roma. Genes 2020, 11, 516. [Google Scholar] [CrossRef] [PubMed]
- Gosadi, I.M.; Goyder, E.C.; Teare, M.D. Investigating the Potential Effect of Consanguinity on Type 2 Diabetes Susceptibility in a Saudi Population. Hum. Hered. 2014, 77, 197–206. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Variables | Levels | Non-Obese | Obese | p-Value |
|---|---|---|---|---|
| Demographic Data | ||||
| Age, years | Mean ± SD | 36.96 ± 14.7 | 41.49 ± 16.2 | 0.005 * |
| Sex | Female | 78 (33.9) | 65 (42.2) | 0.10 |
| Male | 152 (66.1) | 89 (57.8) | ||
| BMI, kg/m2 | Mean ± SD | 21.36 ± 3.2 | 33.65 ± 3.9 | <0.001 * |
| Comorbidities | ||||
| T2DM | Positive | 99 (43.0) | 78 (50.6) | 0.14 |
| Dyslipidemia | Positive | 26 (11.3) | 35 (22.7) | 0.004 * |
| Hypertension | Positive | 27 (11.7) | 31 (20.1) | 0.029 * |
| SBP, mmHg | 123.62 ± 18.0 | 126.14 ± 18.6 | 0.18 | |
| DBP, mmHg | 72.5 ± 9.95 | 72.11 ± 9.60 | 0.65 | |
| Laboratory Data | ||||
| Glycemic state | Serum glucose, mmol/L | 5.7 (4.7–6.9) | 6.0 (5.24–7.7) | 0.003 * |
| Glycosylated hemoglobin (HbA1c), % | 6.0 ± 1.67 | 6.27 ± 1.77 | 0.13 | |
| Serum insulin, mIU/L | 160 (73.1–324.5) | 220 (86–448) | 0.006 * | |
| Lipid profile | Triglyceride, mmol/L | 1.51 (1.03–2.3) | 1.58 (1.04–2.5) | 0.80 |
| Total cholesterol, mmol/L | 4.83 ± 0.99 | 4.85 ± 1.07 | 0.85 | |
| HDL-cholesterol, mmol/L | 1.16 (0.97–1.35) | 1.14 (0.97–1.33) | 0.61 | |
| LDL-cholesterol, mmol/L | 2.73 (2.24–3.53) | 2.91 (2.19–3.5) | 0.74 |
| Gene | SNP ID | Alleles | Non-Obese | Obese | Genotypes | Non-Obese | Obese | Model | Adjusted OR (95%CI) | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| GSTM1 | rs1056806 | C allele | 416 (93) | 270 (88) | C/C | 195 (87) | 119 (78) | Heterozygote | 2.02 (1.15–3.55) | 0.015 * |
| T allele | 32 (7) | 36 (12) | C/T | 26 (12) | 32 (21) | Homozygote | 1.09 (0.18–6.63) | 0.05 | ||
| p-value | 0.22 | T/T | 3 (1) | 2 (1) | Dominant | 1.92 (1.11–3.31) | 0.019 * | |||
| p-value | 0.039 * | Recessive | 0.98 (0.16–5.91) | 0.98 | ||||||
| GSTT1 | rs1111875 | A allele | 259 (76) | 159 (75) | A/A | 96 (56) | 61 (58) | Heterozygote | 0.87 (0.52–1.45) | 0.42 |
| C allele | 81 (24) | 53 (25) | A/C | 67 (39) | 37 (35) | Homozygote | 1.80 (0.62–5.21) | 0.45 | ||
| p-value | 0.86 | C/C | 7 (4) | 8 (8) | Dominant | 0.96 (0.59–1.56) | 0.86 | |||
| p-value | 0.41 | Recessive | 1.90 (0.67–5.40) | 0.23 | ||||||
| GSTP1 | rs1695 | A allele | 221 (65) | 133 (62) | A/A | 73 (43) | 45 (42 | Heterozygote | 0.93 (0.55–1.58) | 0.46 |
| G allele | 117 (35) | 81 (38) | A/G | 75 (44) | 43 (40) | Homozygote | 1.47 (0.71–3.03) | 0.49 | ||
| p-value | 0.65 | G/G | 21 (12) | 19 (18) | Dominant | 1.05 (0.64–1.71) | 0.85 | |||
| p-value | 0.45 | Recessive | 1.52 (0.78–2.99) | 0.22 | ||||||
| MGST3 | rs7744724 | C allele | 339 (96) | 214 (98) | C/C | 165 (93) | 105 (96) | Heterozygote | 0.70 (0.21–2.33) | 0.20 |
| T allele | 15 (4) | 4 (2) | C/T | 9 (5) | 4 (4) | Homozygote | 0.00 (0.00–NA) | 0.57 | ||
| p-value | 0.11 | T/T | 3 (2) | 0 (0) | Dominant | 0.52 (0.16–1.67) | 0.25 | |||
| p-value | 0.33 | Recessive | 0.00 (0.00–NA) | 0.09 | ||||||
| SOD1 | rs2234694 | A allele | 338 (97) | 209 (100) | A/A | 166 (95) | 104 (99) | Heterozygote | 0.27 (0.03–2.24) | 0.15 |
| C allele | 10 (3) | 1 (0) | A/C | 6 (3) | 1 (1) | Homozygote | 0.00 (0.00–NA) | 0.17 | ||
| p-value | 0.048 * | C/C | 2 (1) | 0 (0) | Dominant | 0.20 (0.02–1.62) | 0.07 | |||
| p-value | 0.23 | Recessive | 0.00 (0.00–NA) | 0.17 | ||||||
| SOD2 | rs4880 | A allele | 191 (57) | 102 (48) | A/A | 58 (35) | 26 (25) | Heterozygote | 1.49 (0.83–2.67) | 0.13 |
| G allele | 143 (43) | 110 (52) | A/G | 75 (45) | 50 (47) | Homozygote | 1.97 (1.00–3.86) | 0.045 * | ||
| p-value | 0.038 * | G/G | 34 (20) | 30 (28) | Dominant | 1.64 (0.95–2.82) | 0.07 | |||
| p-value | 0.13 | Recessive | 1.54 (0.88–2.72) | 0.13 | ||||||
| SOD3 | rs2536512 | A allele | 170 (51) | 118 (56) | A/A | 39 (23) | 38 (36) | Heterozygote | 0.47 (0.26–0.83) | 0.033 * |
| G allele | 166 (49) | 92 (44) | A/G | 92 (55) | 42 (40) | Homozygote | 0.69 (0.35–1.36) | 0.07 | ||
| p-value | <0.001 * | G/G | 37 (22) | 25 (24) | Dominant | 0.53 (0.31–0.91) | 0.021 * | |||
| p-value | 0.033 * | Recessive | 1.11 (0.62–1.97) | 0.73 | ||||||
| CAT | rs7943316 | A allele | 116 (47) | 52 (40) | A/A | 34 (28) | 13 (20) | Heterozygote | 0.85 (0.43–1.69) | 0.46 |
| T allele | 130 (53) | 78 (60) | T/A | 48 (39) | 26 (40) | Homozygote | 0.60 (0.27–1.35) | 0.90 | ||
| p-value | 0.18 | T/T | 41 (33) | 26 (40) | Dominant | 0.75 (0.40–1.40) | 0.37 | |||
| p-value | 0.46 | Recessive | 0.65 (0.32–1.35) | 0.24 | ||||||
| GPX1 | rs1800668 | A allele | 83 (19) | 72 (25) | A/A | 9 (4) | 15 (10) | Heterozygote | 1.03 (0.64–1.65) | 0.77 |
| G allele | 345 (81) | 218 (75) | G/A | 65 (30) | 42 (29) | Homozygote | 2.65 (1.11–6.32) | 0.048 * | ||
| p-value | 0.041 * (M-H) | G/G | 140 (65) | 88 (61) | Dominant | 1.23 (0.79–1.90) | 0.36 | |||
| p-value | 0.07 | Recessive | 2.63 (1.12–6.18) | 0.024 * | ||||||
| GPX4 | rs713041 | C allele | 176 (52) | 111 (53) | C/C | 42 (25) | 27 (26) | Heterozygote | 0.96 (0.54–1.73) | 0.97 |
| T allele | 160 (48) | 97 (47) | C/T | 92 (55) | 57 (55) | Homozygote | 0.92 (0.44–1.91) | 0.99 | ||
| p-value | 0.82 | T/T | 34 (20) | 20 (19) | Dominant | 0.95 (0.54–1.67) | 0.86 | |||
| p-value | 0.48 | Recessive | 0.94 (0.51–1.74) | 0.84 | ||||||
| NOS3 | rs1799983 | G allele | 254 (75) | 176 (83) | G/G | 97 (57) | 73 (69) | Heterozygote | 0.66 (0.39–1.13) | 0.09 |
| T allele | 84 (25) | 36 (17) | T/G | 60 (36) | 30 (28) | Homozygote | 0.33 (0.09–1.22) | 0.21 | ||
| p-value | 0.029 * | T/T | 12 (7) | 3 (3) | Dominant | 0.61 (0.37–1.02) | 0.06 | |||
| p-value | 0.10 | Recessive | 0.38 (0.10–1.38) | 0.11 | ||||||
| NOS2 | rs2297518 | A allele | 123 (36) | 82 (38) | A/A | 23 (13) | 13 (12) | Heterozygote | 1.34 (0.80–2.26) | 0.51 |
| G allele | 221 (64) | 134 (62) | A/G | 77 (45) | 56 (52) | Homozygote | 1.04 (0.48-2.29) | 0.25 | ||
| p-value | 0.59 | G/G | 72 (42) | 39 (36) | Dominant | 1.27 (0.78-2.09) | 0.34 | |||
| p-value | 0.51 | Recessive | 0.89 (0.43-1.83) | 0.74 | ||||||
| No. | GSTM1 | SOD2 | SOD3 | GPX1 | Frequency | OR (95% CI) | p-Value |
|---|---|---|---|---|---|---|---|
| 1 | C | A | A | G | 0.126 | 1 | --- |
| 2 | C | A | G | G | 0.117 | 3.05 (0.66–14.14) | 0.15 |
| 3 | C | G | A | G | 0.191 | 2.95 (1.27–6.86) | 0.012 * |
| 4 | C | G | G | G | 0.111 | 2.06 (0.35–11.94) | 0.42 |
| 5 | C | G | A | G | 0.080 | 6.15 (1.46–25.87) | 0.014 * |
| 6 | C | A | A | G | 0.067 | 0.52 (0.03–8.27) | 0.64 |
| 7 | C | A | G | G | 0.064 | 0.74 (0.10–5.58) | 0.77 |
| 8 | C | A | A | A | 0.043 | 5.00 (1.23–20.33) | 0.025 * |
| 9 | C | G | G | G | 0.039 | 1.85 (0.02–209.77) | 0.80 |
| 10 | T | G | G | G | 0.030 | 3.25 (1.06–9.95) | 0.040 * |
| 11 | C | G | A | A | 0.026 | 1.45 (0.16–13.23) | 0.74 |
| 12 | C | A | A | A | 0.021 | 9.44 (0.73–122.24) | 0.09 |
| 13 | T | A | A | G | 0.019 | 1.85 (0.02–209.77) | 0.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gusti, A.M.T.; Qusti, S.Y.; Alshammari, E.M.; Toraih, E.A.; Fawzy, M.S. Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study. Antioxidants 2021, 10, 595. https://doi.org/10.3390/antiox10040595
Gusti AMT, Qusti SY, Alshammari EM, Toraih EA, Fawzy MS. Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study. Antioxidants. 2021; 10(4):595. https://doi.org/10.3390/antiox10040595
Chicago/Turabian StyleGusti, Amani M. T., Safaa Y. Qusti, Eida M. Alshammari, Eman A. Toraih, and Manal S. Fawzy. 2021. "Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study" Antioxidants 10, no. 4: 595. https://doi.org/10.3390/antiox10040595
APA StyleGusti, A. M. T., Qusti, S. Y., Alshammari, E. M., Toraih, E. A., & Fawzy, M. S. (2021). Antioxidants-Related Superoxide Dismutase (SOD), Catalase (CAT), Glutathione Peroxidase (GPX), Glutathione-S-Transferase (GST), and Nitric Oxide Synthase (NOS) Gene Variants Analysis in an Obese Population: A Preliminary Case-Control Study. Antioxidants, 10(4), 595. https://doi.org/10.3390/antiox10040595