
Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.)

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chemicals
2.2. Protein Solubility Curve
2.3. Preparation of Kiwicha Protein Isolate (KPI)
2.4. Hydrolysis of Kiwicha Protein Isolate
2.5. Evaluation of Hydrolysis Degree (HD)
2.6. Compositional Analysis of Kiwicha Protein Products
2.7. Determination of Amino Acid Composition by Ultra-High-Performance Liquid Chromatography (HPLC)
2.8. Pepsin-Pancreactic Digestibility
2.9. Molecular Weight (MW) Profile by Fast Protein Liquid Chromatography (FPLC)
2.10. Determination of Antioxidant Activity
2.10.1. DPPH Radical-Scavenging Activity
2.10.2. Reducing Power
2.11. Statistical Analysis
3. Results and Discussion
3.1. Chemical Characterization of Kiwicha Protein Products and Protein Digestibility
3.2. Amino Acid Composition of Kiwicha Protein Products
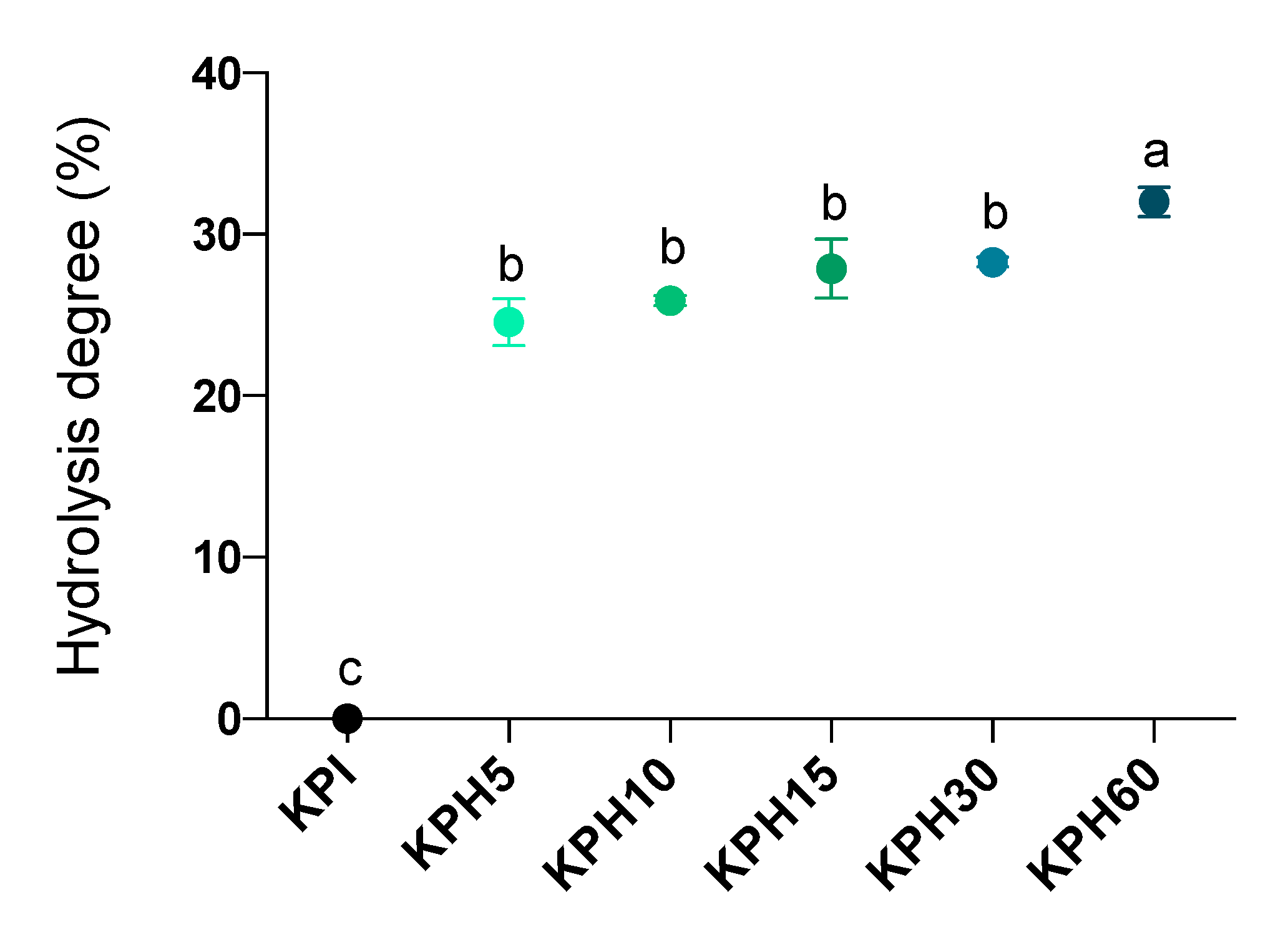
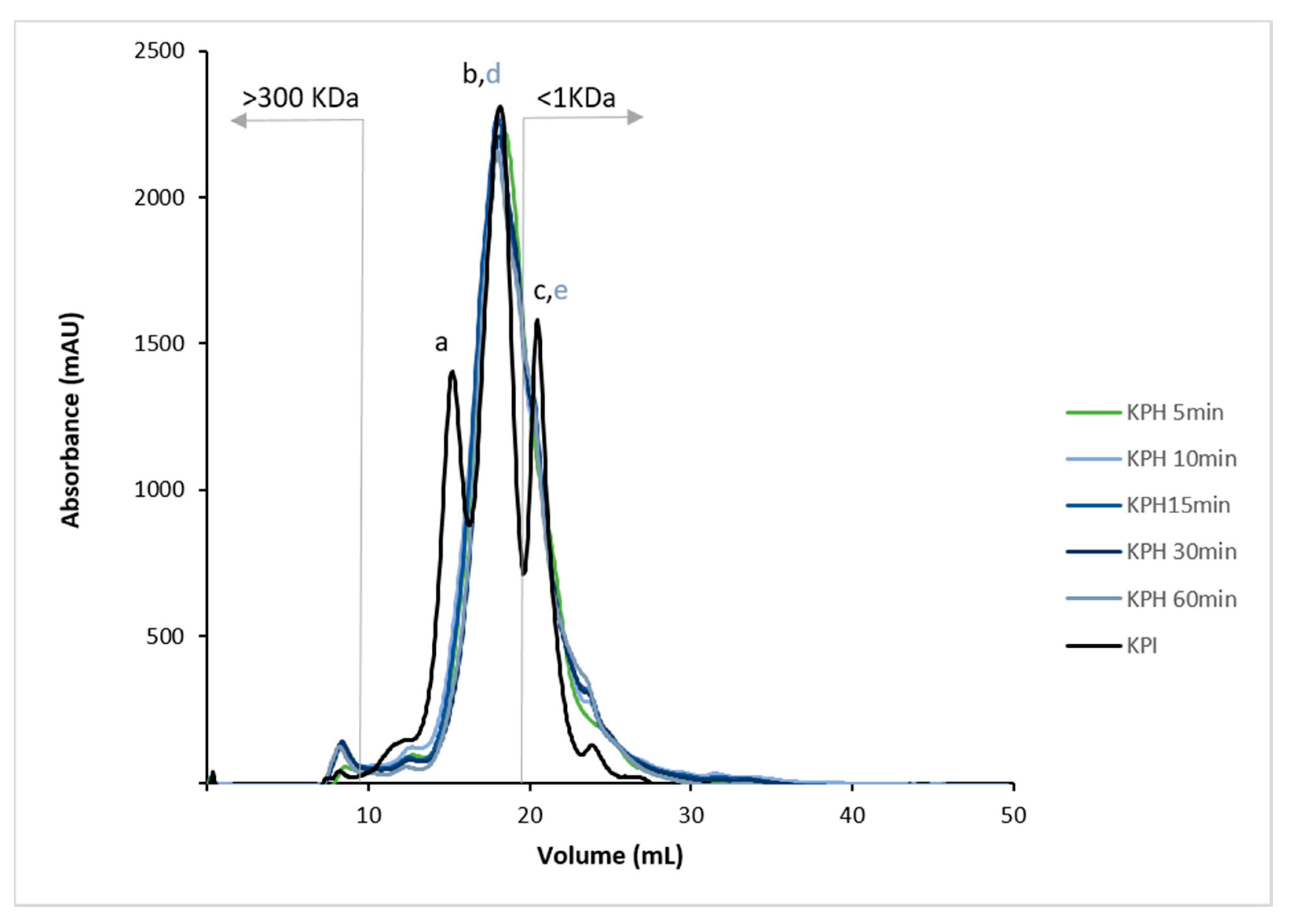
3.3. Analysis of the Hydrolysis Degree and the Molecular Profile of Kiwicha Protein Products
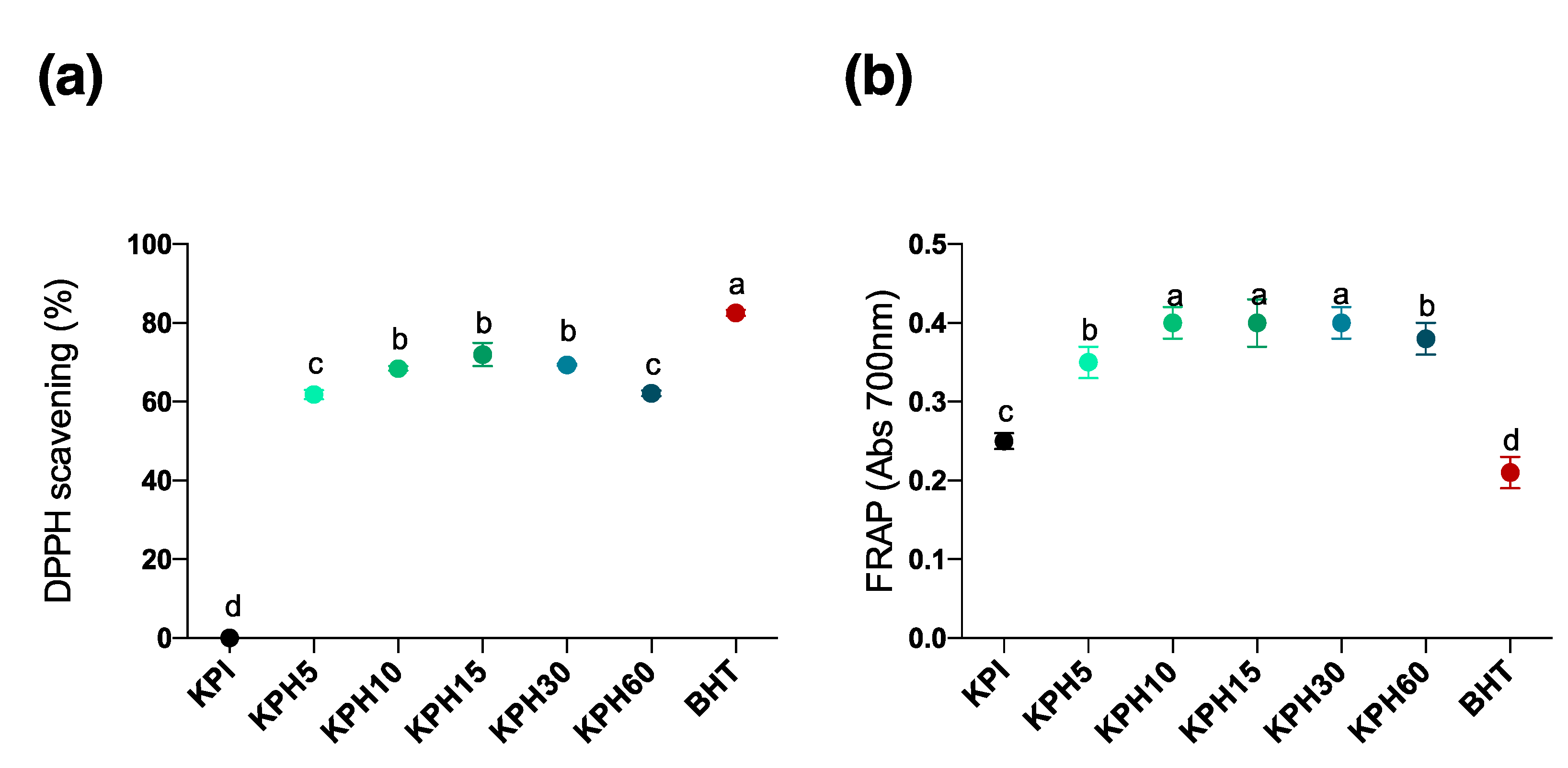
3.4. Determination of the Antioxidant Activity of Kiwicha Protein Products
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rastogi, A.; Shukla, S. Amaranth: A new millennium crop of nutraceutical values. Crit. Rev. Food Sci. Nutr. 2013, 53, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.M.; Chen, L.-W.; Heude, B.; Bernard, J.Y.; Harvey, N.C.; Duijts, L.; Mensink-Bout, S.M.; Polanska, K.; Mancano, G.; Suderman, M.; et al. Dietary inflammatory index and non-communicable disease risk: A narrative review. Nutrients 2019, 11, 1873. [Google Scholar] [CrossRef] [Green Version]
- Jha, J.C.; Ho, F.; Dan, C.; Jandeleit-Dahm, K. A causal link between oxidative stress and inflammation in cardiovascular and renal complications of diabetes. Clin. Sci. 2018, 132, 1811–1836. [Google Scholar] [CrossRef] [PubMed]
- Das, S. Amaranths: The crop of great prospect. In Amaranthus: A Promising Crop of Future, 1st ed.; Springer: Singapore, 2016; p. 48. [Google Scholar] [CrossRef]
- Cheng, A. Shaping a sustainable food future by rediscovering long-forgotten ancient grains. Plant Sci. 2018, 269, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.M.; Silva, P.M.; Martins, J.T.; Pinheiro, A.C.; Vicente, A.A. Emerging opportunities in exploring the nutritional/functional value of amaranth. Food Funct. 2018, 9, 5499–5512. [Google Scholar] [CrossRef]
- Valcárcel-Yamani, B.; Lannes, S.D.S. Applications of quinoa (Chenopodium quinoa Willd.) and amaranth (Amaranthus spp.) and their influence in the nutritional value of cereal based foods. Food Public Health 2012, 2, 265–275. [Google Scholar] [CrossRef]
- Martinez-Lopez, A.; Millan-Linares, M.C.; Rodriguez-Martin, N.M.; Millan, F.; Montserrat-de la Paz, S. Nutraceutical value of kiwicha (Amaranthus caudatus L.). J. Funct. Foods 2020, 65, 103735. [Google Scholar] [CrossRef]
- Marcone, M.F. Evidence confirming the existence of a 7S globulin-like storage protein in Amaranthus hypochondriacus seed. Food Chem. 1999, 65, 533–542. [Google Scholar] [CrossRef]
- Singhal, R.S.; Kulkarni, P.R. Composition of the seeds of some Amaranthus species. J. Sci. Food Agric. 1988, 42, 325–331. [Google Scholar] [CrossRef]
- Singhal, R.S.; Kulkarni, P.R. Amaranths—An underutilized resource. Int. J. Food Sci. Technol. 1988, 23, 125–139. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Miralles, B.; Hernández-Ledesma, B. Release of multifunctional peptides from kiwicha. (Amaranthus caudatus) protein under in vitro gastrointestinal digestion. J. Sci. Food Agric. 2019, 99, 1225–1232. [Google Scholar] [CrossRef] [Green Version]
- Chirinos, R.; Pedrechis, R.; Velásquez-Sánchez, M.; Aquilar-Gálvez, A.; Campos, D. In vitro antioxidant and angiotensin I-converting enzyme inhibitory properties of enzymatically hydrolyzed quinoa (Chenopodium quinoa) and kiwicha (Amaranthus caudatus) proteins. Cereal. Chem. 2020, 97, 949–957. [Google Scholar] [CrossRef]
- Mayne, S.T.; Playdon, M.C.; Rock, C.L. Diet, nutrition, and cancer: Past, present and future. Nat. Rev. Clin. Oncol. 2016, 13, 504. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Dietary and policy priorities for cardiovascular disease, diabetes, and obesity: A comprehensive review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- Montoya-Rodríguez, A.; Gómez-Favela, M.A.; Reyes-Moreno, C.; Milán-Carrillo, J.; González de Mejía, E. Identification of bioactive peptide sequences from amaranth (Amaranthus hypochondriacus) seed proteins and their potential role in the prevention of chronic diseases. Compr. Rev. Food Sci. Food Saf. 2015, 14, 139–158. [Google Scholar] [CrossRef]
- Sangameswaran, B.; Jayakar, B. Anti-diabetic, anti-hyperlipidemic and spermatogenic effects of Amaranthus spinosus Linn. on streptozotocin-induced diabetic rats. J. Nat. Med. 2008, 62, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Girija, K.; Lakshman, K.; Udaya, C.; Sachi, G.S.; Divya, T. Anti–diabetic and anti–cholesterolemic activity of methanol extracts of three species of Amaranthus. Asian Pac. J. Trop. Biomed. 2011, 1, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Jubete, L.; Wijngaard, H.; Arendt, E.K.; Gallagher, E. Polyphenol composition and in vitro antioxidant activity of amaranth, quinoa buckwheat and wheat as affected by sprouting and baking. Food Chem. 2010, 119, 770–778. [Google Scholar] [CrossRef]
- Tironi, V.A.; Añón, M.C. Amaranth proteins as a source of antioxidant peptides: Effect of proteolysis. Food Res. Int. 2010, 43, 315–322. [Google Scholar] [CrossRef]
- Maiyo, Z.C.; Ngure, R.M.; Matasyoh, J.C.; Chepkorir, R. Phytochemical constituents and antimicrobial activity of leaf extracts of three Amaranthus plant species. Afr. J. Biotechnol. 2010, 9, 3178–3182. [Google Scholar] [CrossRef]
- Loaiza, M.A.P.P.; López-Malo, A.; Jiménez-Munguía, M.T. Nutraceutical Properties of Amaranth and Chia Seeds. In Functional Properties of Traditional Foods, 1st ed.; Springer: Boston, MA, USA, 2016; pp. 189–198. [Google Scholar] [CrossRef]
- Rojas-Rivas, E.; Espinoza-Ortega, A.; Thomé-Ortiz, H.; Moctezuma-Pérez, S. Consumers’ perception of amaranth in Mexico. A traditional food with characteristics of functional foods. Br. Food J. 2019, 121, 1190–1202. [Google Scholar] [CrossRef]
- Rojas-Rivas, E.; Espinoza-Ortega, A.; Thomé-Ortíz, H.; Moctezuma-Pérez, S.; Cuffia, F. Understanding consumers’ perception and consumption motives towards amaranth in Mexico using the Pierre Bourdieu’s theoretical concept of Habitus. Appetite 2019, 139, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef] [PubMed]
- Esfandi, R.; Walters, M.E.; Tsopmo, A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon 2019, 5, e01538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lqari, H.; Vioque, J.; Pedroche, J.; Millán, F. Lupinus angustifolius protein isolates: Chemical composition, functional properties and protein characterization. Food Chem. 2002, 76, 349–356. [Google Scholar] [CrossRef]
- Tapan, K.M.; Abdullatif, K.; Abeer, H.; Elsayed, F.A.A.; Ahmed, A.-H. The molecular mass and isoelectric point of plant proteomes. BMC Genom. 2019, 20, 631. [Google Scholar] [CrossRef] [Green Version]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Prosky, L.; Asp, N.G.; Schweizer, T.F.; Devries, J.W.; Furda, I. Determination of insoluble, soluble, and total dietary fiber in foods and food products: Interlaboratory study. J. Assoc. Off. Anal. Chem. 1988, 71, 1017–1023. [Google Scholar] [CrossRef]
- Moores, R.G.; McDermott, D.L.; Wood, T.R. Determination of chlorogenic acid in coffee. Anal. Chem. 1948, 20, 620–624. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Alaiz, M.; Navarro, J.L.; Girón, J.; Vioque, E. Amino acid analysis by high-performance liquid chromatography after derivatization with diethyl ethoxymethylenemalonate. J. Chromatogr. A 1992, 591, 181–186. [Google Scholar] [CrossRef]
- Yust, M.M.; Pedroche, J.; Girón-Calle, J.; Vioque, J.; Millán, F.; Alaiz, M. Determination of tryptophan by high-performance liquid chromatography of alkaline hydrolysates with spectrophotometric detection. Food Chem. 2004, 85, 317–320. [Google Scholar] [CrossRef]
- Sindayikengera, S.; Xia, W.-S. Nutritional evaluation of caseins and whey proteins and their hydrolysates from Protamex. Zhejiang Univ. Sci. B 2006, 7, 90–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.C.; Chen, H.M.; Shiau, C.Y. Free amino acids and peptides as related to antioxidant properties in protein hydrolysates of mackerel (Scomber austriasicus). Food Res. Int. 2003, 36, 949–957. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reactions: Antioxidative activities of product of browning reaction prepared from glucosamine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Montserrat-de la Paz, S.; Rodriguez-Martin, N.M.; Villanueva, A.; Pedroche, J.; Cruz-Chamorro, I.; Millan, F.; Millan-Linares, M. Evaluation of anti-inflammatory and atheroprotective properties of wheat gluten protein hydrolysates in primary human monocytes. Foods 2020, 9, 854. [Google Scholar] [CrossRef]
- FAO FINNUT. Dietary Protein Quality Evaluation in Human Nutrition; FAO of the UN: Auckland, New Zealand, 2017. [Google Scholar]
- Hertzler, S.R.; Lieblen-Boff, C.; Weiler, M.; Allgeier, C. Plant proteins: Assessing their nutritional quality and effects on health and physical function. Nutrients 2020, 12, 3704. [Google Scholar] [CrossRef]
- Joye, I. Protein digestibility of cereal products. Foods 2019, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Shumoy, H.; Pattyn, S.; Raes, K. Tef protein: Solubility characterization, in vitro digestibility and its suitability as a gluten free ingredient. LWT Food Sci. Technol. 2018, 89, 697–703. [Google Scholar] [CrossRef]
- Tapal, A.; Vegarud, G.E.; Sreedharab, A.; Kaul-Tico, P. Nutraceutical protein isolate from pigeon pea (Cajanus cajan) milling waste by-product: Functional aspects and digestibility. Food Funct. 2019, 10, 2710. [Google Scholar] [CrossRef]
- Nelson, D.L.; Cox, M.M. Principles of Biochemistry, 7th ed.; Lehninger, W.H., Ed.; Freeman & Company: New York, NY, USA, 2017. [Google Scholar]
- Colovic, M.B.; Vesna, M.V.; Dragan, M.D.; Krstict, D.Z. Sulfur-containing amino acids: Protective role against free radicals and heavy metals. Curr. Med. Chem. 2018, 25, 324–335. [Google Scholar] [CrossRef]
- Hong, G.P.; Min, S.G.; Jo, Y.J. Anti-Oxidative and anti-aging activities of porcine by-product collagen hydrolysates produced by commercial proteases: Effect of hydrolysis and ultrafiltration. Molecules 2019, 24, 1104. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; Santos-Sánchez, G.; Pedroche, J.; Fernández-Pachón, M.S.; Millán, F.; Carrillo-Vico, A. Immunomodulatory and antioxidant properties of wheat gluten protein hydrolysates in human peripheral blood mononuclear cells. Nutrients 2020, 12, 1673. [Google Scholar] [CrossRef]
- Hou, C.; Wu, L.; Wang, Z.; Saguer, E.; Zhang, D. Purification and Identification of Antioxidant Alcalase-Derived Peptides from Sheep Plasma Proteins. Antioxidants 2019, 8, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millán-Linares, M.C.; Bermúdez, B.; Yust, M.M.; Millán, F.; Pedroche, J. Anti-inflammatory activity of lupine (Lupinus angustifolius L.) protein hydrolysates in THP-1- derived macrophages. J. Funct. Foods 2014, 8, 224–233. [Google Scholar] [CrossRef] [Green Version]
- Millán-Linares, M.C.; Millán, F.; Pedroche, J.; Yust, M.M. GPETAFLR: A new anti-inflammatory peptide from Lupinus angustifolius L. protein hydrolysate. J. Funct. Foods 2015, 18, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Martin, N.M.; Toscano, R.; Villanueva, A.; Pedroche, J.; Millan, F.; Montserrat-de la Paz, S.; Millan-Linares, M.C. Neuroprotective protein hydrolysates from hemp (Cannabis sativa L.) seeds. Food Funct. 2019, 10, 6732–6739. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Martin, N.M.; Montserrat-de la Paz, S.; Toscano, R.; Grao-Cruces, E.; Villanueva, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Hemp (Cannabis sativa L.) protein hydrolysates promote anti-inflammatory response in primary human monocytes. Biomolecules 2020, 10, 803. [Google Scholar] [CrossRef] [PubMed]
- Snigdha, G.; Kaustav, M. Structural-features of food-derived bioactive peptides with anti-inflammatory activity: A brief review. J. Nutr. Biochem. 2018, 2019, e12531. [Google Scholar] [CrossRef]
- Linlin, C.; Huidan, D.; Hengmin, C.; Jing, F.; Zhicai, Z.; Junliang, D.; Yinglun, L.; Xun, W.; Ling, Z. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [Green Version]
- Lujuan, X.; Qingquan, F.; Guanghong, Z.; Wangang, Z. A review of antioxidant peptides derived from meat muscle and by-products. Antioxidants 2016, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Hambraeus, L. Protein and Amino Acids in Human Nutrition. In Reference Module in Biomedical Sciences. Amaranthus: A Promising Crop of Future, 1st ed.; Elsevier: Amsterdam, The Netherland, 2014; p. 48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fat (Seed Flour, %) | 5.73 ± 0.12 | ||||||
|---|---|---|---|---|---|---|---|
| KDF | KPI | KPH5 | KPH10 | KPH15 | KPH30 | KPH60 | |
| Proteins | 15.88 ± 0.13 | 84.52 ± 0.55 | 80.93 ± 0.57 | 80.77 ± 0.12 | 80.30 ± 0.36 | 76.89 ± 0.01 | 78.50 ± 0.17 |
| Moisture | 7.01 ± 0.17 | 0.96 ± 0.10 | 2.86 ± 0.13 | 1.59 ± 0.27 | 1.29 ± 0.09 | 4.54 ± 0.04 | 1.93 ± 0.19 |
| Ash | 2.18 ± 0.09 | 1.75 ± 0.05 | 7.17 ± 0.25 | 7.54 ± 0.36 | 8.40 ± 0.16 | 9.13 ± 0.59 | 9.31 ± 0.05 |
| Fiber | 10.63 ± 0.16 | 7.86 ± 0.08 | 6.58 ± 0.02 | 7.37 ± 0.09 | 8.31 ± 0.31 | 8.09 ± 0.24 | 8.04 ± 0.18 |
| Sugar | 1.73 ± 0.22 | 0.78 ± 0.12 | 0.37 ± 0.01 | 0.52 ± 0.06 | 0.46 ± 0.02 | 0.30 ± 0.04 | 0.29 ± 0.02 |
| Polyphenols | 0.01 ± 0.00 | 0.03 ± 0.00 | 0.04 ± 0.00 | 0.05 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 | 0.04 ± 0.00 |
| Others 1 | 62.56 | 4.1 | 2.05 | 2.16 | 1.2 | 1.01 | 1.89 |
| In vitro protein digestibility 2 | 60.74 ± 0.04 | 77.97 ± 2.93 | 79.18 ± 0.66 | 67.00 ± 3.23 | 69.58 ± 0.63 | 71.74 ± 1.66 | 73.28 ± 0.51 |
| AA (%) | KDF | KPI | KPH5 | KPH10 | KPH15 | KPH30 | KPH60 | FAO 1,2 |
|---|---|---|---|---|---|---|---|---|
| Indispensable Amino Acids | ||||||||
| His | 24.30 ± 0.0 | 27.60 ± 0.0 | 30.80 ± 0.1 | 30.80 ± 0.1 | 31.20 ± 0.1 | 31.10 ± 0.0 | 31.50 ± 0.0 | 15 |
| Ile | 30.80 ± 0.3 | 43.80 ± 0.0 | 42.70 ± 0.1 | 44.60 ± 0.1 | 44.90 ± 0.2 | 44.90 ± 0.2 | 45.20 ± 0.1 | 30 |
| Leu | 61.20 ± 0.0 | 72.00 ± 0.1 | 71.90 ± 0.2 | 73.90 ± 0.1 | 74.00 ± 0.1 | 74.30 ± 0.1 | 75.60 ± 0.0 | 59 |
| Lys | 62.30 ± 0.1 | 53.10 ± 0.1 | 55.00 ± 0.1 | 55.20 ± 0.1 | 55.00 ± 0.1 | 55.00 ± 0.0 | 55.90 ± 0.0 | 45 |
| Met + Cys | 32.50 ± 0.1 | 34.60 ± 0.1 | 28.20 ± 0.1 | 35.70 ± 0.1 | 34.5 ± 0.1 | 37.10 ± 0.1 | 36.60 ± 0.1 | 22 |
| Met | 13.1 ± 0.1 | 20.90 ± 0.2 | 13.3 ± 0.1 | 21.5 ± 0.1 | 20.0 ± 0.2 | 22.7 ± 0.2 | 20.50 ± 0.2 | 16 |
| Cys | 19.40 ± 0.1 | 13.70 ± 0.0 | 14.90 ± 0.1 | 14.20 ± 0.1 | 14.50 ± 0.1 | 14.40 ± 0.1 | 15.10 ± 0.1 | 6 |
| Phe + Tyr | 77.90 ± 0.1 | 92.60 ± 0.1 | 93.10 ± 0.2 | 94.80 ± 0.1 | 93.70 ± 0.1 | 95.50 ± 0.0 | 95.40 ± 0.2 | 38 |
| Thr | 39.8 ± 0.1 | 41.0 ± 0.0 | 42.9 ± 0.1 | 43.1 ± 0.1 | 43.0 ± 0.0 | 43.0 ± 0.0 | 43.7 ± 0.0 | 23 |
| Trp | 9.2 ± 0.0 | 11.5 ± 0.0 | 16.7 ± 0.0 | 16.0 ± 0.0 | 15.2 ± 0.0 | 12.6 ± 0.1 | 8.7 ± 0.0 | 6 |
| Val | 35.3 ± 0.3 | 46.10 ± 0.1 | 45.2 ± 0.2 | 47.4 ± 0.1 | 48.1 ± 0.2 | 47.30 ± 0.2 | 48.20 ± 0.2 | 39 |
| Total | 373.50 | 422.30 | 426.50 | 441.50 | 439.60 | 440.80 | 440.80 | 277 |
| Dispensable Amino Acids | ||||||||
| Asp + Asn | 94.3 ± 0.4 | 92.6 ± 0.2 | 99.7 ± 0.2 | 97.7 ± 0.2 | 97.1 ± 0.1 | 96.8 ± 0.0 | 98.3 ± 0.0 | n.a |
| Glu + Gln | 183.0 ± 0.6 | 155.1 ± 0.1 | 168.4 ± 0.3 | 163.3 ± 0.4 | 162.4 ± 0.2 | 160.8 ± 0.0 | 162.7 ± 0.1 | n.a |
| Ser | 79.1 ± 0.4 | 55.4 ± 0.1 | 60.3 ± 0.1 | 59.0 ± 0.1 | 58.5 ± 0.1 | 58.4 ± 0.1 | 59.4 ± 0.1 | n.a |
| Gly | 83.1 ± 0.4 | 49.3 ± 0.1 | 52.8 ± 0.1 | 51.6 ± 0.1 | 52.5 ± 0.1 | 51.4 ± 0.0 | 53.0 ± 0.1 | n.a |
| Arg | 87.3 ± 0.2 | 89.4 ± 0.1 | 95.2 ± 0.2 | 94.2 ± 0.2 | 93.8 ± 0.1 | 93.7 ± 0.1 | 94.8 ± 0.1 | n.a |
| Ala | 41.4 ± 0.2 | 41.1 ± 0.0 | 43.0 ± 0.1 | 43.1 ± 0.1 | 43.3 ± 0.1 | 43.0 ± 0.0 | 43.6 ± 0.0 | n.a |
| Pro | 58.5 ± 0.7 | 94.9 ± 0.6 | 54.4 ± 1.9 | 49.7 ± 1.4 | 52.7 ± 0.5 | 54.8 ± 0.6 | 48.3 ± 1.0 | n.a |
| Tyr | 34.9 ± 0.1 | 40.8 ± 0.2 | 41.0 ± 0.2 | 41.5 ± 0.1 | 40.4 ± 0.2 | 42.3 ± 0.0 | 41.1 ± 0.3 | n.a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paz, S.M.-d.l.; Martinez-Lopez, A.; Villanueva-Lazo, A.; Pedroche, J.; Millan, F.; Millan-Linares, M.C. Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.). Antioxidants 2021, 10, 645. https://doi.org/10.3390/antiox10050645
Paz SM-dl, Martinez-Lopez A, Villanueva-Lazo A, Pedroche J, Millan F, Millan-Linares MC. Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.). Antioxidants. 2021; 10(5):645. https://doi.org/10.3390/antiox10050645
Chicago/Turabian StylePaz, Sergio Montserrat-de la, Alicia Martinez-Lopez, Alvaro Villanueva-Lazo, Justo Pedroche, Francisco Millan, and Maria C. Millan-Linares. 2021. "Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.)" Antioxidants 10, no. 5: 645. https://doi.org/10.3390/antiox10050645
APA StylePaz, S. M.-d. l., Martinez-Lopez, A., Villanueva-Lazo, A., Pedroche, J., Millan, F., & Millan-Linares, M. C. (2021). Identification and Characterization of Novel Antioxidant Protein Hydrolysates from Kiwicha (Amaranthus caudatus L.). Antioxidants, 10(5), 645. https://doi.org/10.3390/antiox10050645