
Can Rice Growth Substrate Substitute Rapeseed Growth Substrate in Rapeseed Blanket Seedling Technology? Lesson from Reactive Oxygen Species Production and Scavenging Analysis



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experiment Design
2.2. Sampling
2.3. Seedling Quality Assessment
2.4. ROS Production and Enzymatic Activity Analysis
2.4.1. ROS Production and Scavenging Capacity Analysis
2.4.2. Enzymatic Analysis for ROS Scavenging
2.4.3. Ascorbic Acid Pathway Analysis
2.4.4. Glutathione Pathway Analysis
2.4.5. Other Antioxidants
2.5. RNA-Seq Analysis
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR) Analysis
2.7. Statistics
3. Results
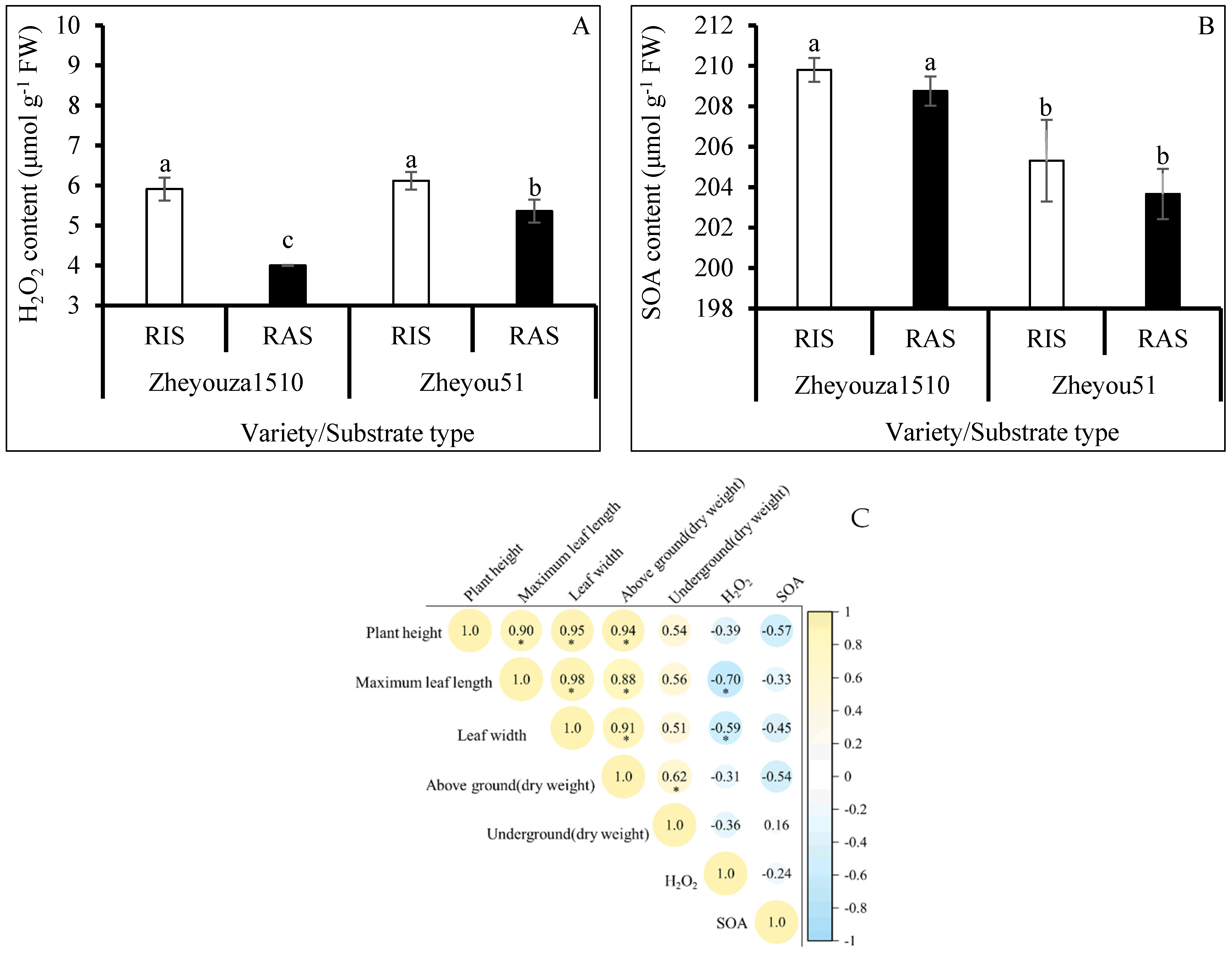
3.1. Seedling Quality and ROS Production under Different Crop Growth Substrates and Genotypes
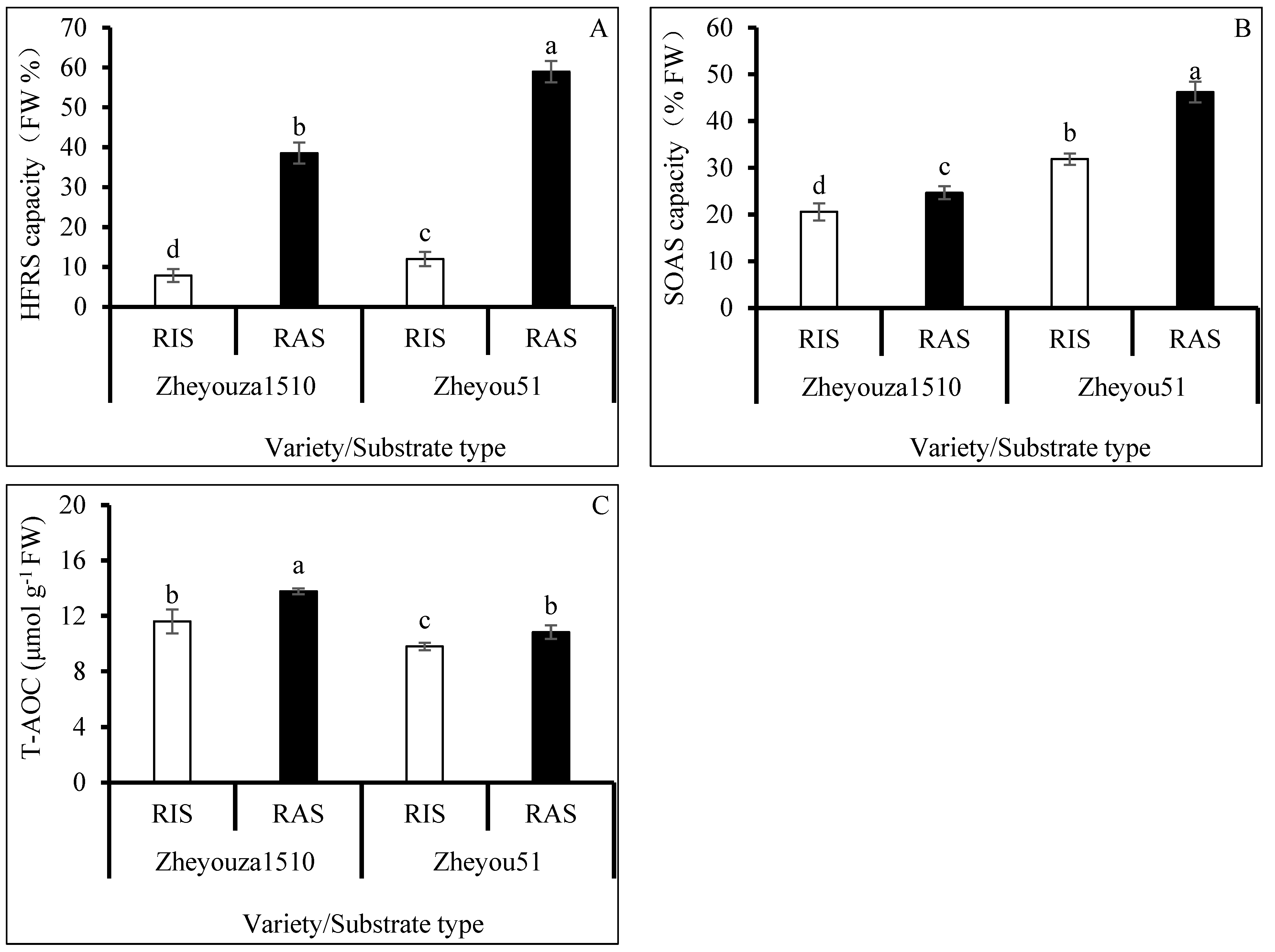
3.2. ROS Scavenging Capacity under Different Crop Growth Substrates and Genotypes
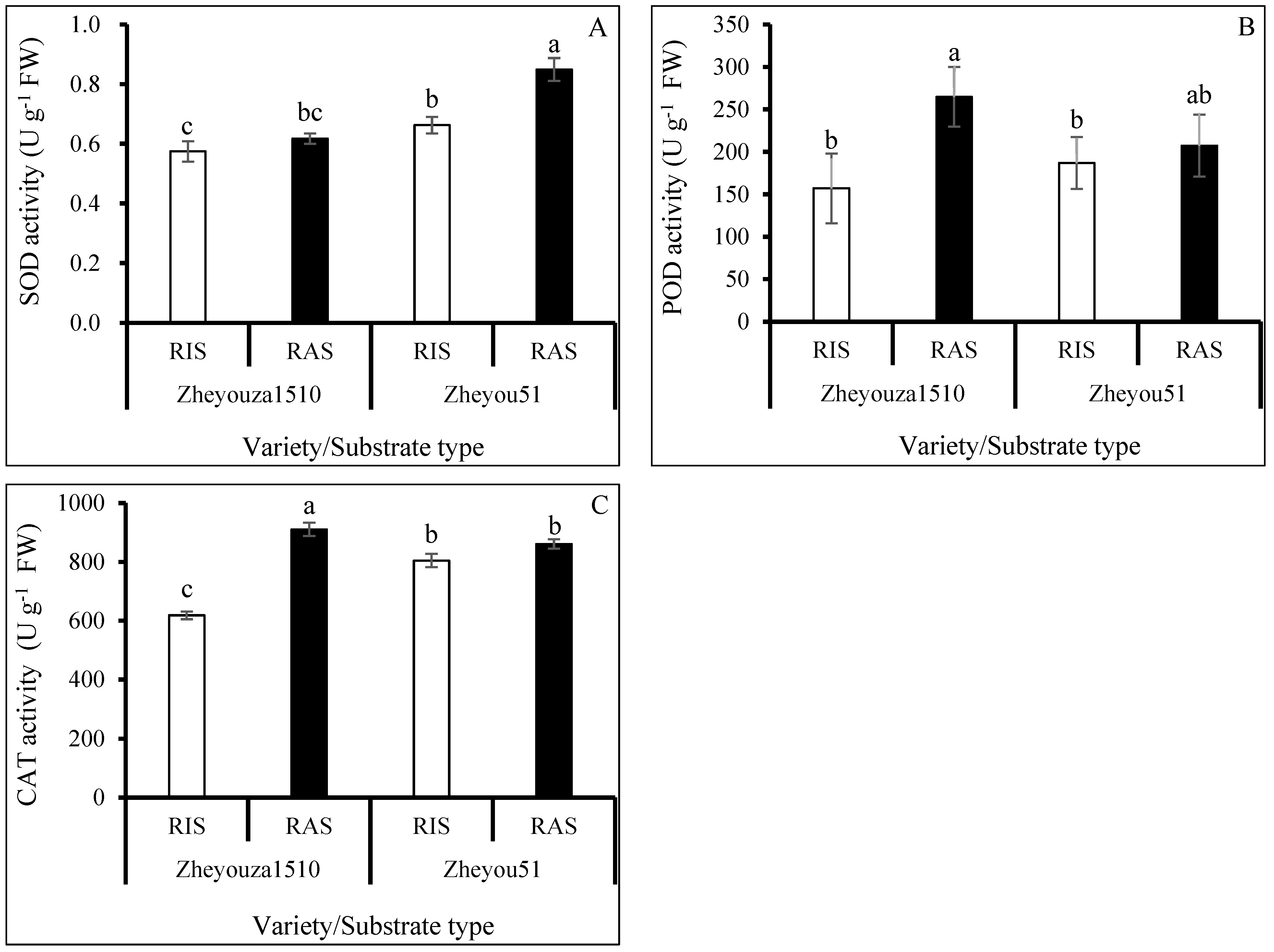
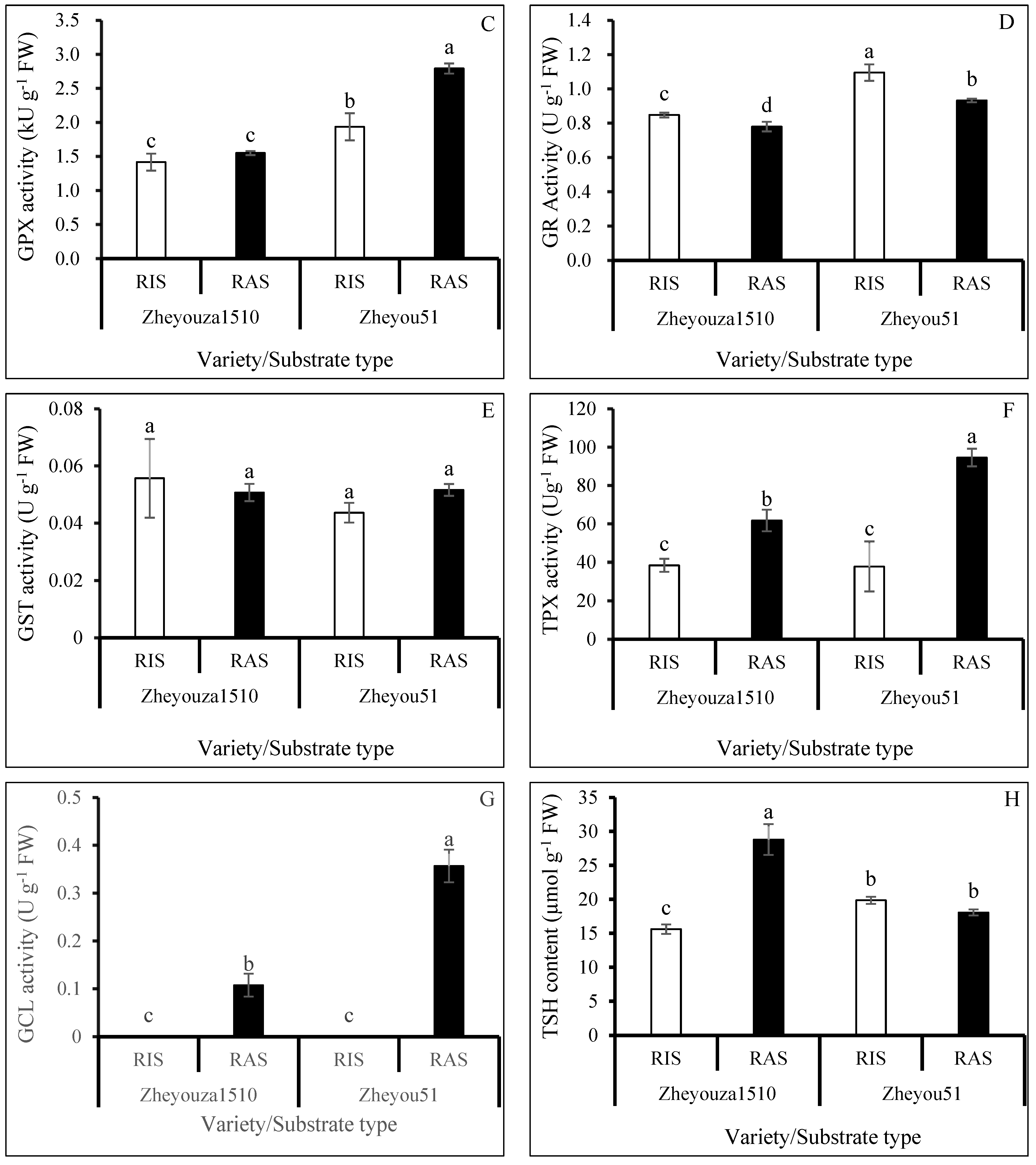
3.3. Enzymatic Activities Analysis on ROS Scavenging Capacity under Different Crop Growth Substrates and Genotypes
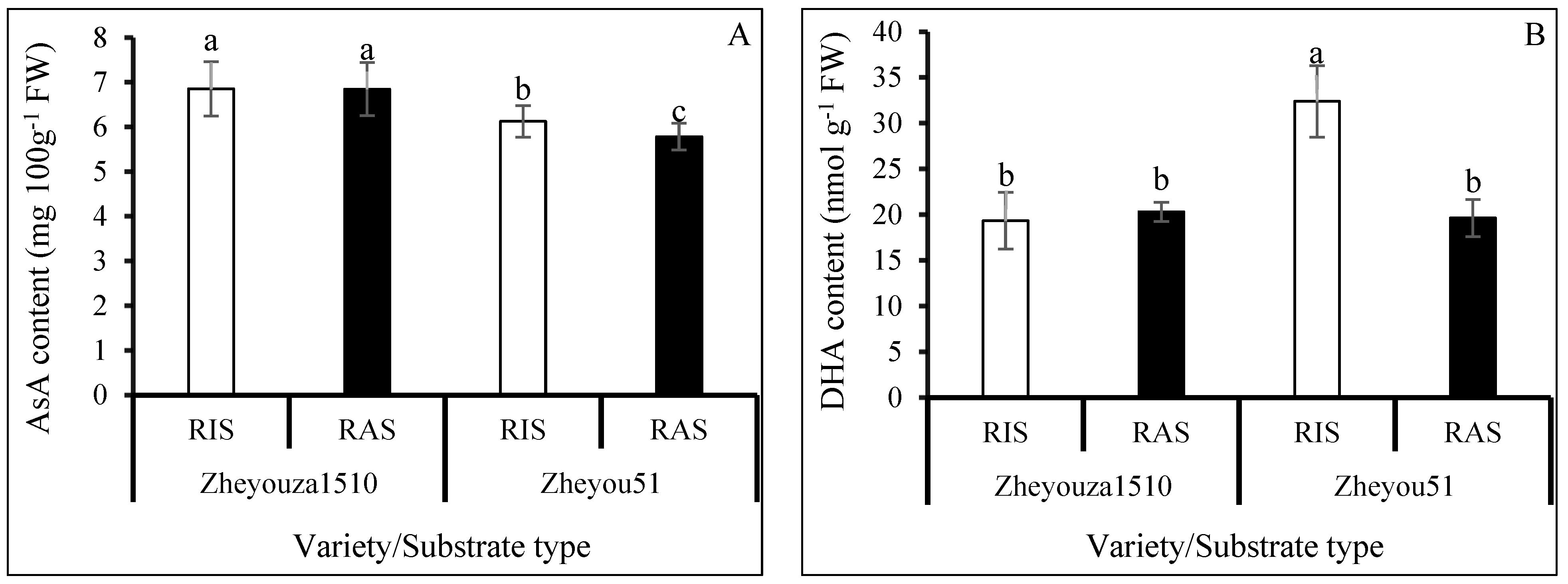
3.4. Ascorbic Acid Metabolism under Different Crop Growth Substrates and Genotypes
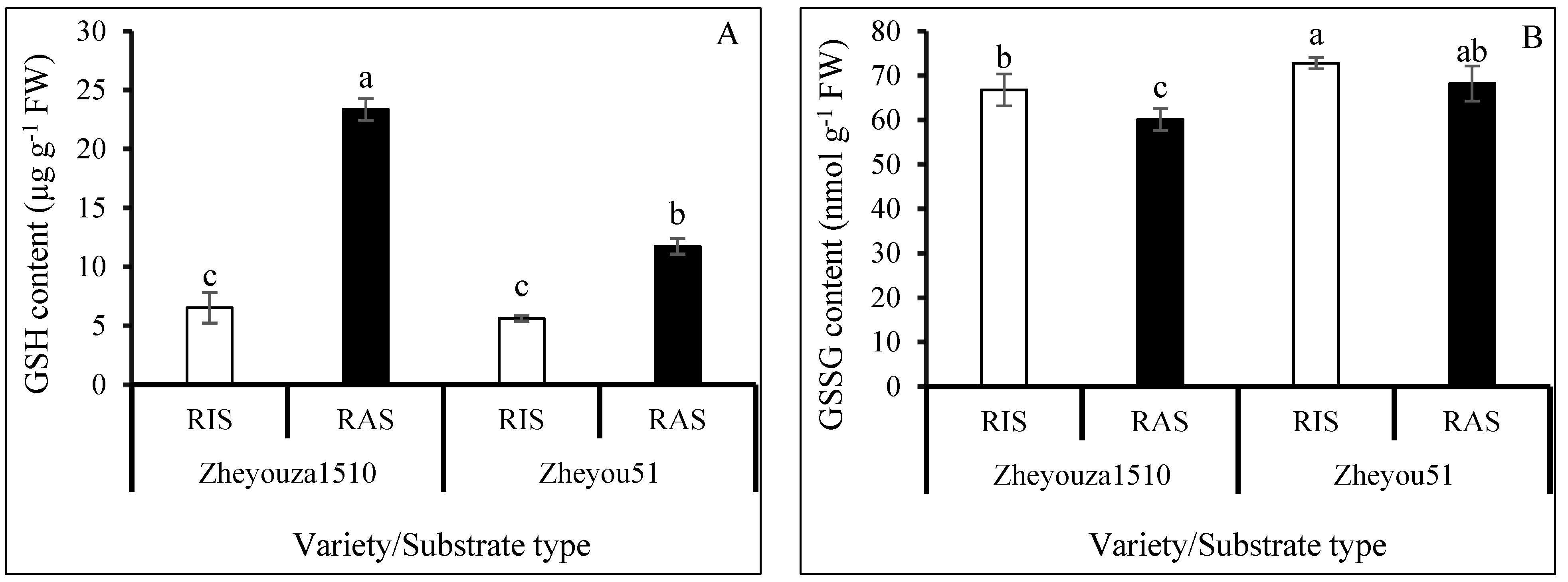
3.5. Glutathione Metabolism for ROS Scavenging under Different Crop Growth Substrates and Genotypes
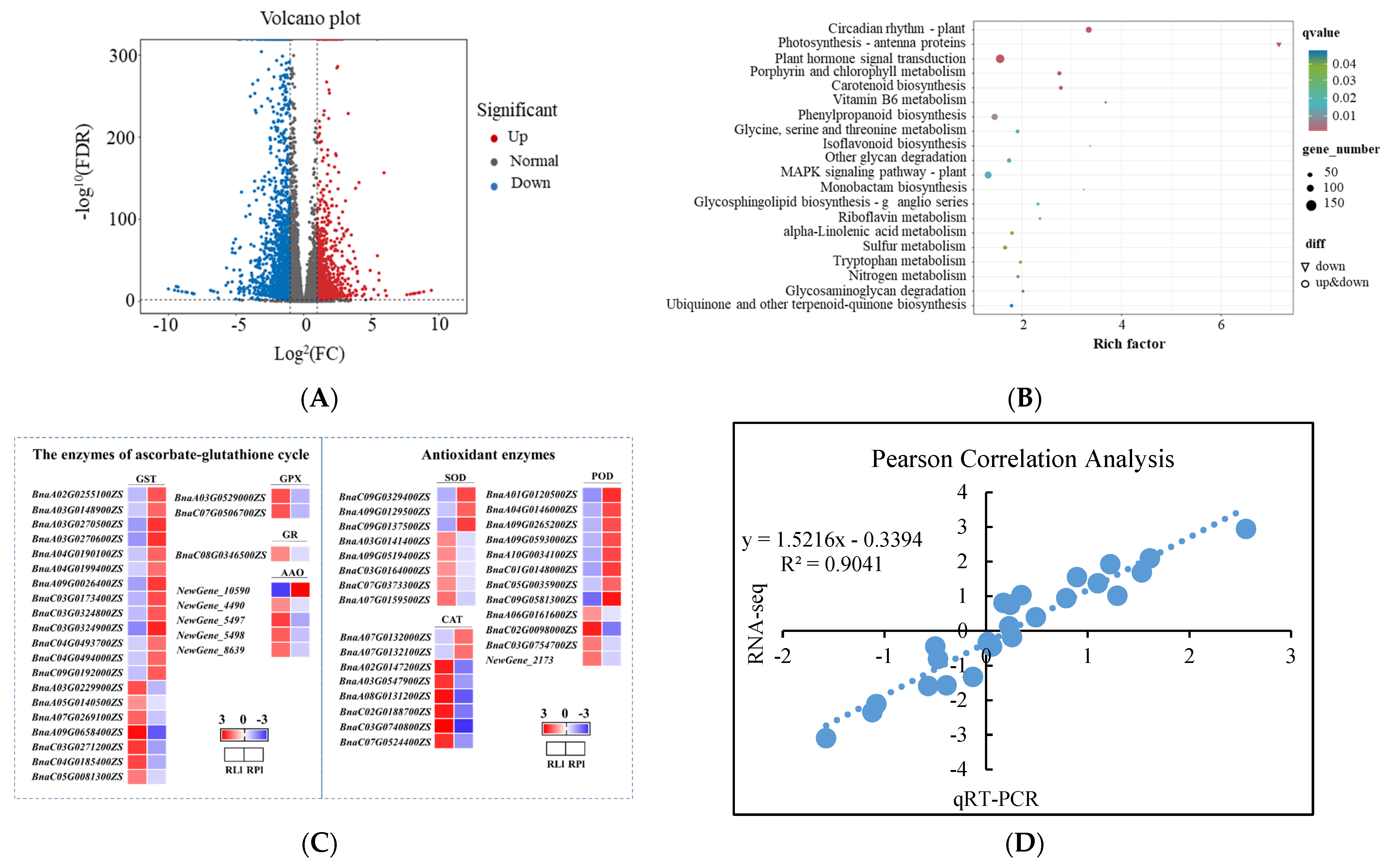
3.6. RNA-Seq Analysis of the ROS Scavenging Capacity under Different Crop Growth Substrates and Genotypes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhen, M.; Terzaghi, W.; Wang, H.Z.; Hua, W. Integrated strategies for increasing rapeseed yield. Trends Plant Sci. 2022, 27, 742–745. [Google Scholar] [CrossRef]
- Shah, S.; Weinholdt, C.; Jedrusik, N.; Molina, C.; Zou, J.; Große, I.; Schiessl, S.; Jung, C.; Emrani, N. Whole-transcriptome analysis reveals genetic factors underlying flowering time regulation in rapeseed (Brassica napus L.). Plant Cell Envion. 2018, 41, 1935–1947. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, S.L.; Ye, X.S.; Xu, F.S. BnaA4.BOR2 contributes the tolerance of rapeseed to boron deficiency by improving the transport of boron from root to shoot. Plant Physiol. Biochem. 2024, 208, 108508. [Google Scholar] [CrossRef]
- Ye, J.; Liang, H.B.; Zhao, X.Y.; Li, N.; Song, D.J.; Zhan, J.P.; Liu, J.; Wang, X.F.; Tu, J.X.; Varshney, R.K.; et al. A systematic dissection in oilseed rape provides insight into the genetic architecture and molecular mechanism of yield heterosis. Plant Biotechnol. J. 2023, 21, 1479–1495. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; El-Badri, A.M.; Li, M.L.; Batool, M.; Wang, C.Y.; Shao, D.L.; Kuai, J.; Wang, B.; Xu, Z.H.; Zhao, J.; et al. Micro-ridge-furrow soil moisture regulation technology improves seedling quality and yield of winter rapeseed. Soil Tillage Res. 2024, 237, 105960. [Google Scholar] [CrossRef]
- Rivelli, G.M.; Calderini, D.F.; Abeledo, L.G.; Miralles, D.J.; Rondanini, D.P. Yield and quality traits of wheat and rapeseed response to source-sink ratio and heat stress in post-flowering. Eur. J. Agron. 2024, 152, 127028. [Google Scholar] [CrossRef]
- Li, Z.J.; Liu, F.C.; Wu, W. Optimising nitrogen management strategies to minimise lodging risk while sustaining high seed yield in rapeseed. Eur. J. Agron. 2023, 142, 126671. [Google Scholar] [CrossRef]
- Geng, G.T.; Cakmak, I.; Ren, T.; Lu, Z.F.; Lu, J.W. Effect of magnesium fertilization on seed yield, seed quality, carbon assimilation and nutrient uptake of rapeseed plants. Field Crop Res. 2021, 264, 108082. [Google Scholar] [CrossRef]
- Zuo, Q.S.; Wang, L.; Zheng, J.D.; You, J.J.; Yang, G.; Leng, S.H.; Liu, J.Y. Effects of uniconazole rate on agronomic traits and physiological indexes of rapeseed blanket seedling. Oil Crop Sci. 2020, 5, 198–204. [Google Scholar] [CrossRef]
- Zhang, K.; Wen, Y.X.; Tang, J.Q.; Zhang, Y.B.B.; Peng, X.X.; Ji, Y.W.; Sun, J.; Liu, X.Y. The regulation of photosynthesis and growth of rapeseed seedling by the interaction of red and yellow lights with blue light. Environ. Exp. Bot. 2024, 225, 105869. [Google Scholar] [CrossRef]
- Zhi, X.M.; Bian, X.H.; Yu, J.L.; Xiao, X.L.; Duan, B.; Huang, F.Y.; Jiang, Z.; Zhou, G.S.; Ma, N. Comparative metabolomics analysis of tolerant and sensitive genotypes of rapeseed (Brassica napus L.) seedlings under drought stress. Agric. Water Manag. 2024, 296, 108797. [Google Scholar] [CrossRef]
- Xu, J.S.; Zhan, H.; Xie, Y.P.; Tian, G.; Xie, L.L.; Xu, B.B.; Zhao, Y.G.; Lu, G.Y.; Zhang, X.K. Associative transcriptomics study dissects the genetic architecture of seedling biomass-related traits in rapeseed (Brassica napus L.). Plant Breed. 2021, 140, 285–293. [Google Scholar] [CrossRef]
- Hooks, T.; Niu, G.H.; Ganjegune, G. Seedling emergence and seedling growth of mustard and rapeseed genotypes under salt stress. Agrosyst. Geosci. Environ. 2018, 2, 1–8. [Google Scholar] [CrossRef]
- Wang, X.C.; Li, Z.H.; Tan, S.Y.; Li, H.W.; Qi, L.; Wang, Y.W.; Chen, J.T.; Yang, C.Y.; Chen, J.Y.; Qin, Y.J.; et al. Research on density grading of hybrid rice machine-transplanted blanket-seedlings based on multi-source unmanned aerial vehicle data and mechanized transplanting test. Comput. Electron. Agric. 2024, 222, 109070. [Google Scholar] [CrossRef]
- Ke, J.; Xin, X.M.; Li, G.H.; Ding, Y.F.; Dou, F.G.; Wang, S.H.; Liu, Z.H.; Tang, S.; Ding, D.Q.; Chen, L. Effect of different controlled-release nitrogen fertilisers on ammonia volatilisation, nitrogen use efficiency and yield of blanket-seedling machine-transplanted rice. Field Crop Res. 2017, 205, 147–156. [Google Scholar] [CrossRef]
- Alotaibi, N.M.; Alotaibi, M.M.; Younis, U.; Hussain, G.S.; Dawar, K.; Hareem, M.; Ghoneim, A.M.; Fahad, S.; Danish, S. Zn-quantum dot biochar regulates antioxidants and nutrient uptake to improve rapeseed growth and yield in drought stress. Plant Stress 2024, 11, 100286. [Google Scholar] [CrossRef]
- Ding, X.Y.; Xu, J.S.; Huang, H.; Qiao, X.; Shen, M.Z.; Chen, Y.; Zhang, X.K. Unraveling waterlogging tolerance-related traits with QTL analysis in reciprocal intervarietal introgression lines using genotyping by sequencing in rapeseed (Brassica napus L.). J. Integr. Agric. 2020, 19, 1974–1983. [Google Scholar] [CrossRef]
- Mohamed, I.A.A.; Shalby, N.; El-Badri, A.M.; Batool, M.; Wang, C.Y.; Wang, Z.K.; Salah, A.; Rady, M.M.; Kuai, J.; Wang, B.; et al. RNA-Seq analysis revealed key genes associated with salt tolerance in rapeseed germination through carbohydrate metabolism, hormone, and MAPK signaling pathways. Ind. Crop Prod. 2022, 176, 114262. [Google Scholar] [CrossRef]
- Brooker, R.; Brown, L.K.; George, T.S.; Pakeman, R.J.; Palmer, S.; Ramsay, L.; Schöb, C.; Schurch, N.; Wilkinson, M.J. Active and adaptive plasticity in a changing climate. Trends Plant Sci. 2022, 57, 717–728. [Google Scholar] [CrossRef]
- Ali, S.; Tyagi, A.; Bae, H. ROS interplay between plant growth and stress biology: Challenges and future perspectives. Plant Physiol. Biochem. 2023, 203, 108032. [Google Scholar] [CrossRef]
- Qin, Q.Q. RO: Important factor in plant stem cell fate regulation. J. Plant Physiol. 2023, 289, 154082. [Google Scholar] [CrossRef]
- Peláez-Vico, M.Á.; Fichman, Y.; Zandalinas, S.I.; Van Breusigem, F.; Karpiński, S.W.; Mittler, R. ROS and redox regulation of cell-to-cell and systematic signaling in plants during stress. Free. Radic. Biol. Med. 2022, 193, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, A.; Kumar, A.; Kaur, N. ROS and oxidative burst: Roots in plant development. Plant Divers. 2020, 42, 33–43. [Google Scholar] [CrossRef]
- Trevisan, S.; Trentin, A.R.; Ghisi, R.; Masi, A.; Quaggiotti, S. Nitrate affects transcriptional regulation of UPBEAT1 and ROS localisation in roots of Zea mays L. Physiol. Plant. 2019, 166, 794–811. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Li, Y.; Sun, J.; Du, L.; Zhang, Y.; Yu, Q.; Liu, X. Comparative physiological and proteomic response to abrupt low temperature stress between two winter wheat cultivars differing in low temperature tolerance. Plant Biol. 2013, 15, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Pignatello, J.J. Analytical methods for selectively determining hydrogen peroxide, peroxymonosulfate and peroxydisulfate in their binary mixtures. Water Res. 2024, 253, 121256. [Google Scholar] [CrossRef]
- Ahmed, S.; Jubair, A.; Hossain, M.A.; Hossain, M.M.; Azam, M.S.; Biswas, M. Free radical-scavenging capacity and HPLC-DAD screening of phenolic compounds from pulp and seed of Syzygium claviflorum fruit. J. Agric. Food Res. 2021, 6, 100203. [Google Scholar] [CrossRef]
- Vo, Q.V.; Hoa, N.T.; Thong, N.M.; Mechler, A. The hydroperoxyl and superoxide anion radical scavenging activity of anthocyanidins in physiological environments: Theoretical insights into mechanisms and kinetics. Phytochemistry 2021, 192, 112968. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin-Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
- Gao, W.D.; Liu, B.C.; Phetmany, S.; Li, J.H.; Wang, D.N.; Liu, Z.Y.; Gao, C.Q. ThDIV2, an R-R-type MYB transcription factor of Tamarix hispida, negatively regulates cadmium stress by modulating ROS hemeostasis. Environ. Exp. Bot. 2023, 214, 105453. [Google Scholar] [CrossRef]
- Wang, L.R.; Li, R.; Shi, X.J.; Wei, L.; Li, W.; Shao, Y.Z. Ripening patterns (off tree and on tree) affect physiology, quality, and ascorbic acid metabolism of mango fruit (cv. Guifei). Sci. Hortic. 2023, 315, 111971. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Xing, M.L.; Guo, X.R.; Wu, M.; Liu, F.; Sun, L.L.; Zhang, P.; Duan, M.; Fan, W.X.; Xu, J. Physiological and transcriptomic analyses reveal that phytohormone pathways and glutathione metabolism are involved in the arsenite toxicity response in tomatoes. Sci. Total Environ. 2023, 899, 165676. [Google Scholar] [CrossRef]
- Kumar, S.; Kayastha, A.M. Soybean (Glycine max) urease: Significance of sulfhydryl groups in urea catalysis. Plant Physiol. Biochem. 2020, 48, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Georgiou, C.D.; Zisimopoulos, D.; Argyropoulou, V.; Kalaitzopoulou, E.; Ioannou, P.V.; Salachas, G.; Grune, T. Protein carbonyl determination by a rhodamine B hydrazide-based fluorometric assay. Rodox Biol. 2018, 17, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.Q.; Chen, D.Z.; Pan, Q.; Li, F.F.; Zhao, Z.G.; Ge, X.H.; Li, Z.Y. Production of red-flowered oilseed rape via the ectopic 616 expression of Orychophragmus violaceus OvPAP2. Plant Biotechnol. J. 2018, 16, 367–380. [Google Scholar] [CrossRef]
- Liu, Y.J.; Ye, S.H.; Yuan, G.G.; Ma, X.W.; Heng, S.P.; Yi, B.; Ma, C.Z.; Shen, J.X.; Tu, J.X.; Fu, T.D.; et al. Gene silencing of BnaA09.ZEP and BnaC09.ZEP confers orange color in Brassica napus flowers. Plant J. 2020, 4, 932–949. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Marín-Guirao, J.I.; Rodríguez-Romera, P.; Lupión-Rodríguez, B.; Camacho-Ferre, F.; Tello-Marquina, J.C. Effect of trichoderma on horticultural seedlings’ growth promotion depending on inoculum and substrate type. J. Appl. Microbiol. 2016, 16, 1095–1102. [Google Scholar] [CrossRef]
- Wang, X.Y.; Liu, M.J.; Ciampitti, I.A.; Cui, J.W.; Fang, K.C.; Zhao, S.C.; He, P.; Zhou, W. Benefits and trade-offs of replacing inorganic fertilizer by organic substrate in crop production: A global meta-analysis. Sci. Total Environ. 2024, 925, 171781. [Google Scholar] [CrossRef]
- Aghdam, A.M.; Sayfzadeh, S.; Rad, A.H.S.; Valadabadi, S.A.; Zakerin, H.R. The assessment of water stress and delay cropping on quantitative and qualitative traits of rapeseed genotypes. Ind. Crop Prod. 2019, 131, 160–165. [Google Scholar] [CrossRef]
- Idrissi, I.S.E.; Kettani, R.; Brhadda, N.; Louali, A.; Channaoui, S.; Gaboune, F.; Nabloussi, A. Variation of rapeseed genotype’s reaction to drought during flowering and identification of tolerant-genotypes selection index. J. Agric. Food Res. 2023, 14, 100872. [Google Scholar] [CrossRef]
- Zhang, S.T.; Shan, Y.X.; Li, Y.; He, J.X.; Jiang, Y.M. Hydrogen peroxide receptors regulate chilling injury of banana fruit during low-temperature storage. Postharvest Biol. Technol. 2024, 214, 112985. [Google Scholar] [CrossRef]
- Neto, V.G.; Riberio, P.R.; Del-Bem, L.E.; Bernal, D.T.; Lima, S.T.C.; Ligterink, W.; Fernandez, L.G.; de Castro, R.D. Characterization of the superoxide dismutase gene family in seeds of two Ricinus communis L. genotypes submitted to germination under water restriction conditions. Environ. Exp. Bot. 2018, 155, 453–463. [Google Scholar] [CrossRef]
- Heydari, M.; Asadi-Rad, H.; Hosseinzadeh, J.; Hajinia, S.; Wait, D.A.; Prevosto, B. Managing semi-arid oak forests (Quercus brantii Lindl.): Mature oak trees of different dimensions create contrasted microhabitats influencing seedling quality. J. Environ. Manag. 2022, 304, 114269. [Google Scholar] [CrossRef] [PubMed]
- Lei, W.S.; Ding, Y.F.; Li, G.H.; Tang, S.; Wang, S.H. Effects of soilless substrates on seedling quality and the growth of transplanted super japonica rice. J. Integr. Agric. 2017, 16, 1053–1063. [Google Scholar] [CrossRef]
- Chakraborty, K.; Bishi, S.L.; Goswami, N.; Singh, A.L.; Zala, P.V. Differential fine-regulation of enzyme driven ROS detoxification network imparts salt tolerance in contrasting peanut genotypes. Environ. Exp. Bot. 2016, 128, 79–90. [Google Scholar] [CrossRef]
- Forghani, A.H.; Aghaie, P. The comparison expression of Rare cold inducible2 (RIC2), some enzymatic antioxidant, and proline in two tolerant and sensitive genotypes chickpea under drought stress. Plant Stress 2024, 11, 100419. [Google Scholar] [CrossRef]
- Singh, A.; Parihar, S.; Shekhawat, G.S. Cd-induced cytotoxicity and its HO-1 and ROS quenching enzyme-mediated regulation in 2-3 leaf stage seedlings of Sorghum bicolor: An important millet crop of the arid & semi-arid regions. J. Trace Elem. Miner. 2024, 9, 100165. [Google Scholar]
- Wu, W.B.; Miao, Y.M.; Kong, J.; Lindsey, K.; Zhang, X.L.; Min, L. ROS signaling and its involvement in abiotic stress with emphasis on heat stress-driven anther sterility in plants. Crop Envion. 2024, 3, 65–74. [Google Scholar]
- Alwutayd, K.M.; Esmail, S.M.; Saad-El-Din, H.I.; El-moneim, D.A.; Draz, I.S. Metabolic and genotypic mechanisms involved in early defensive response of powdery mildew-infected wheat. Physiol. Mol. Plant Pathol. 2023, 126, 102035. [Google Scholar] [CrossRef]
- Rajonandraina, T.; Rakotoson, T.; Wissuwa, M.; Ueda, Y.; Razafimbelo, T.; Andriamananjara, A.; Kirk, G.J.D. Mechanism of genotypic differences in tolerance of iron toxicity in field-grown rice. Field Crop Res. 2023, 298, 108953. [Google Scholar] [CrossRef]
- Hashemi, S.; Razaghi-Moghadam, Z.; Laitinen, R.A.E.; Nikoloski, Z. Relative flux trade-offs and optimization of metabolic network functionalities. Comput. Struct. Biotechnol. 2022, 20, 3963–3971. [Google Scholar] [CrossRef] [PubMed]
- Cornejo-Castillo, F.M.; Inomura, K.; Zehr, J.P.; Follows, M.J. Metabolic trade-offs constrain the cell size ratio in a nitrogen-fixing symbiosis. Cell 2024, 187, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Mishra, B.; Chand, S.; Sangwan, N.S. ROS management is mediated by ascorbate-glutathione-α-tocopherol triad in coordination with secondary metabolic pathway under cadmium stress in Withania somnifera. Plant Physiol. Biochem. 2019, 139, 620–629. [Google Scholar] [CrossRef]
- Zhou, C.Y.; Zhang, X.; Chen, Y.X.; Liu, X.P.; Sun, Y.X.; Xiong, B. Glutathione alleviate the cadmium exposure-caused porcine oocyte meiotic defects via eliminating the excessive ROS. Environ. Pollut. 2019, 255, 113194. [Google Scholar] [CrossRef] [PubMed]
- Somegowda, V.K.; Reddy, S.E.D.; Gaddameedi, A.; Kiranmayee, K.N.S.U.; Naravula, J.; Kishor, P.B.K.; Penna, S. Genomic breeding approaches for developing Sorghum bicolor lines with stress resilience and other agronomic traits. Curr. Plant Biol. 2024, 37, 100314. [Google Scholar] [CrossRef]
- Bazyklo, A.; Parzonko, A.; Jeż, W.; Osińska, E.; Kiss, A.K. Inhibition of ROS production, photoprotection, and total phenolic, flavonoids and ascorbic acid content of fresh herb juice and extracts from the leaves and flowers of Tropaeolum majus. Ind. Crop Prod. 2014, 55, 19–24. [Google Scholar]
- Lan, X.G.; Yang, J.; Abhinandan, K.; Nie, Y.Z.; Li, X.Y.; Li, Y.H.; Samuel, M.A. Flavonoids and ROS play opposing roles in mediating pollination in ornamental kale (Brassica oleracea var. acephala). Mol. Plant 2017, 10, 1361–1364. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Zhang, Z.N.; Guo, S.J.; Qu, P.Y.; Liu, J.P.; Cheng, C.Z. Characterization of blueberry glutathione S-transferase (GST) genes and functional analysis of VcSGTF8 reveal the role of ‘MYB/bHLH-GSTF’ module in anthocyanin accumulation. Ind. Crop Prod. 2024, 218, 119006. [Google Scholar] [CrossRef]
- Park, H.; Kim, H.; Abassi, S.; Bui, Q.T.N.; Ki, J. Two novel glutathione S-transferase (GST) in the toxic marine dinoflagellate Alexandrium pacificum and their transcriptional responses to environmental contaminants. Sci. Total Environ. 2024, 915, 169983. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rapeseed Variety | Substrate | Plant Height (cm) | Maximum Leaf Size (cm) | Dry Matter (g plant−1) | ||
|---|---|---|---|---|---|---|
| Length | Width | Aboveground | Underground | |||
| Zheyouza 1510 | RIS | 1.47 ± 0.78 c | 3.97 ± 0.24 c | 2.97 ± 0.41 c | 0.42 ± 0.04 c | 0.10 ± 0.01 a |
| RAS | 3.23 ± 0.19 b | 6.15 ± 0.17 b | 4.84 ± 0.19 b | 0.52 ± 0.02 b | 0.11 ± 0.01 a | |
| Zheyou 51 | RIS | 1.61 ± 0.48 c | 3.87 ± 0.43 c | 3.16 ± 0.30 c | 0.38 ± 0.06 c | 0.07 ± 0.01 b |
| RAS | 5.63 ± 0.07 a | 6.70 ± 0.23 a | 5.78 ± 0.26 a | 0.79 ± 0.04 a | 0.13 ± 0.01 a | |
| Cultivar (C) | ** | ns | * | ** | ** | |
| Substrate (S) | ** | ** | ** | * | ns | |
| C × S | ns | ns | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, K.; Ren, Y.; Zhang, H.; Lin, B.; Hao, P.; Hua, S. Can Rice Growth Substrate Substitute Rapeseed Growth Substrate in Rapeseed Blanket Seedling Technology? Lesson from Reactive Oxygen Species Production and Scavenging Analysis. Antioxidants 2024, 13, 1022. https://doi.org/10.3390/antiox13081022
Yi K, Ren Y, Zhang H, Lin B, Hao P, Hua S. Can Rice Growth Substrate Substitute Rapeseed Growth Substrate in Rapeseed Blanket Seedling Technology? Lesson from Reactive Oxygen Species Production and Scavenging Analysis. Antioxidants. 2024; 13(8):1022. https://doi.org/10.3390/antiox13081022
Chicago/Turabian StyleYi, Kaige, Yun Ren, Hui Zhang, Baogang Lin, Pengfei Hao, and Shuijin Hua. 2024. "Can Rice Growth Substrate Substitute Rapeseed Growth Substrate in Rapeseed Blanket Seedling Technology? Lesson from Reactive Oxygen Species Production and Scavenging Analysis" Antioxidants 13, no. 8: 1022. https://doi.org/10.3390/antiox13081022
APA StyleYi, K., Ren, Y., Zhang, H., Lin, B., Hao, P., & Hua, S. (2024). Can Rice Growth Substrate Substitute Rapeseed Growth Substrate in Rapeseed Blanket Seedling Technology? Lesson from Reactive Oxygen Species Production and Scavenging Analysis. Antioxidants, 13(8), 1022. https://doi.org/10.3390/antiox13081022