
(Chemical) Roles of HOCl in Rheumatic Diseases

Institute for Medical Physics and Biophysics, Medical Faculty, Leipzig University, 04103 Leipzig, Germany
*
Author to whom correspondence should be addressed.
Antioxidants 2024, 13(8), 921; https://doi.org/10.3390/antiox13080921 (registering DOI)
Submission received: 27 March 2024
/
Revised: 16 July 2024
/
Accepted: 26 July 2024
/
Published: 29 July 2024
(This article belongs to the Special Issue Heme Peroxidases in (Patho)Physiological Reactions and Disease Progression)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Chronic rheumatic diseases such as rheumatoid arthritis (RA) are characterized by a dysregulated immune response and persistent inflammation. The large number of neutrophilic granulocytes in the synovial fluid (SF) from RA patients leads to elevated enzyme activities, for example, from myeloperoxidase (MPO) and elastase. Hypochlorous acid (HOCl), as the most important MPO-derived product, is a strong reactive oxygen species (ROS) and known to be involved in the processes of cartilage destruction (particularly regarding the glycosaminoglycans). This review will discuss open questions about the contribution of HOCl in RA in order to improve the understanding of oxidative tissue damaging. First, the (chemical) composition of articular cartilage and SF and the mechanisms of cartilage degradation will be discussed. Afterwards, the products released by neutrophils during inflammation will be summarized and their effects towards the individual, most abundant cartilage compounds (collagen, proteoglycans) and selected cellular components (lipids, DNA) discussed. New developments about neutrophil extracellular traps (NETs) and the use of antioxidants as drugs will be outlined, too. Finally, we will try to estimate the effects induced by these different agents and their contributions in RA.
1. Introduction
Rheumatic diseases such as osteoarthritis (OA) or rheumatoid arthritis (RA) are abundant diseases, especially in industrialized countries with high life expectancy of the citizens [1]. RA represents an autoimmune disease where the immune system attacks the joints and/or its constituents like proteins and/or carbohydrates, leading to persistent inflammation and damage. On the other hand, OA is a degenerative joint disease caused by excessive mechanical wear on joints, resulting in the cartilage destruction. The onset of RA is relatively rapid, while OA develops slowly over the years and regularly causes symptoms at elevated age. RA also affects organs and systems beyond the joints and may cause systemic symptoms such as fatigue, fever, and loss of appetite, which are typically not seen in OA. Although both diseases are symptomatically different, we will discuss them here simultaneously since both have the same unique (chemical) characteristics: the damaging and modification of the articular cartilage layer of the joint and/or the inflammation of the synovial membrane (synovitis) (Figure 1) [2].
In the USA alone, more than 40 million citizens (about 12% of the overall US population) suffer from arthritis [3]. The socio-economic consequences (including but not limited to the inability to work and early retirement as personal consequences) and the annual all-cost ranged from USD 12,509 to USD 20,919 per RA patient in 2019 [4]. Unfortunately, the incidence of the disease is also increasing. The reasons are unknown to date [3], but akinesia (meaning absent movement, i.e., the inability to perform a clinically perceivable movement [5]) as well as extreme sports may also be responsible for the observed increase. Although medial laymen often assume that the term arthritis represents just a single disease, it actually comprises more than 100 different types of arthritic diseases, which collectively affect virtually all parts of the human body—not only the joints [6].
Therefore, it is regrettable that neither a convincing cure of the disease nor reliable methods of early disease diagnosis are available—although there is currently intense research being performed [7]. Here, we will discuss changes in (articular) cartilage as well as the joint (synovial) fluid since both are affected by OA and RA, respectively.
2. Composition of Articular Cartilage and Synovial Fluid
Since both cartilage and synovial fluid (SF) have a rather unique composition (Figure 1), the architecture of (hyaline, articular) cartilage and the composition of SF will be shortly discussed. Further details are available, for instance, in [8,9]. With 70–80% of the wet tissue weight, water is the most abundant constituent of the cartilage [10]. This is crucial for maintaining the function of cartilage, i.e., its shock-absorbing properties and the frictionless motion of the bones within the joint [11]. So-called chondrocytes in the hyaline articular cartilage are responsible for both the synthesis and secretion of major components of the extracellular matrix (ECM) [12,13]. Cartilage ECM contains a particular network of highly hydrated proteoglycans and collagen fibrils [13].
2.1. Collagen
Different collagen types are expressed in articular cartilage (60–70% of the dry weight), which are essential for the elasticity and shear strength of the tissue caused by the presence of water between the collagen fibrils [14]. Collagen has a remarkable amino acid composition with high amounts of the amino acids glycine (Gly), proline (Pro) and hydroxyproline (Hyp) [15]. In particular, Hyp occurs exclusively in collagen, where it is essential for the noteworthy (thermal) stability of collagen [16]. Gly, Pro and Hyp mediate the triple-helical structure of collagen, leading to the formation of collagen fibers [8]. This makes (native) collagen insoluble in virtually all solvents. Only the denaturation of collagen into gelatin makes it soluble in hot water, which is widely used in the food industry [17]. Under physiological conditions, the insolubility of collagen is essential to prevent swelling (water-uptake) of the cartilage. Collagen degradation by proteolytic enzymes is therefore (in addition to further aspects) closely linked to cartilage degradation [18]. The in vitro degradation of cartilage by collagenase results in a strongly increased water content [19] under otherwise identical conditions. This has two different reasons: On the one hand, the water binding of the polysaccharides of cartilage is overestimated since the collagen moiety is partially lost. On the other hand, the swelling limitation of the collagen network is reduced. Thus, proteolytic enzymes were believed to represent the main effectors of cartilage degradation in the past [20]. Intact, triple-helical collagen cannot be digested by common proteolytic enzymes (such as trypsin), while denatured collagen (called gelatin; without collagen fibers) is highly sensitive to the majority of all proteases [19].
2.2. Proteoglycans
The main proteoglycan (aggrecan) in the hyaline cartilage consists of a core protein connected with both chondroitin and keratan sulfate chains [21], as shown in Figure 1. Chondroitin sulfate (CS) and keratan sulfate (KS), as well as hyaluronic acid (HA, as the backbone of the aggrecan aggregate), are known as glycosaminoglycans (GAGs) and are susceptible to degradation under oxidative conditions. Thus, oxidatively modified GAG species will be the focus of this review and their reaction behavior will be discussed in more detail compared to other cartilage constituents.
Accordingly, calcification processes of cartilage may lead to pathological conditions due to the reduction in the water content by shielding the negative charges of the native cartilage [22].
HA is the simplest GAG, just a polysaccharide made of glucuronic acid and N-acetylglucosamine disaccharide units, which are 1→3 glycosidically linked and lead to high-molecular-weight HA by 1→4 glycosidic linkages. HA is the only GAG without any sulfate residues and is only a minor constituent in the proteoglycan aggregates [23]. Nevertheless, HA is present in the cartilage as well as (in significant concentrations) in the SF. Its sulfated analogues such as KS and CS are much more abundant in the cartilage tissue. The sulfation of GAGs does not occur randomly but seems to be species-dependent. For instance, bovine cartilage has a higher content of the 4-sulfate compared to human cartilage [24]. There are also many indications that the ratio between sulfated and non-sulfated GAGs is altered in the aged cartilage [25]. Although this topic is outside the scope of this paper, there are also indications that the “sulfation code” plays a major role in signal transduction [26]. This indicates that the sulfation of cartilage polysaccharides does not occur randomly.
Intact SF is a highly viscous fluid and acts as joint lubricant and shock absorber, as well as an important source of nutrition for the articular cartilage, which borders on the synovial joint [27]. SF may be considered an “ultrafiltrate” of blood [28], i.e., the concentrations of salts and small molecules are nearly identical in both body fluids. Providing detailed compositional information about physiological SF is difficult since it can hardly be obtained from healthy volunteers due to ethical reasons. Thus, control samples are often missing in related studies. Furthermore, SF is a very viscous body fluid. This high viscosity is caused by the significant concentration of high-molecular-weight HA (about 2–3 mg/mL). Many attempts were performed to use the mechanical (viscoelastic) properties as a diagnostic criterion of degenerative joint diseases in the past [29]. This was motivated by the finding that the viscosity of SF can be reduced in the presence of reactive oxygen species (ROS) [30] released by stimulated neutrophilic granulocytes. Additionally, SF from patients with inflammatory joint diseases are characterized by significant myeloperoxidase (MPO) activities in the U/mL range (one unit (U) is defined as the amount of MPO that will reduce 1.0 µmol H2O2 per minute at 25 °C, pH 6.0) [31]. However, it has to be emphasized that the availability of SF to monitor pathological changes has nowadays significantly decreased: puncturing was still a very common medical tool a few decades ago since the majority of rheumatic patients suffered from swollen knees. This was reduced due to the development of more efficient drugs and treatments—particularly by “biologicals” [32].
3. Mechanisms of Cartilage Degradation
It is commonly accepted that inflammatory joint diseases affect both cartilage composition and layer thickness by the release of enzymes and/or ROS generation. However, there is still no consensus on the details about the mechanisms of cartilage degeneration [33]. Only two models are established:
- Chondrocytes (in the cartilage) or synoviocytes (at the interior of the articular capsule) are negatively affected by products of typical inflammatory cells such as neutrophilic granulocytes (vide infra). This leads to a reduced overall ECM synthesis and the increased degradation of the ECM [34]. Additionally, the “quality” of the de novo generated ECM may be poor (fibrocartilage instead of hyaline cartilage).
- Inflammatory cells such as neutrophilic granulocytes, macrophages or T cells release harmful enzymes that can degrade the ECM. Matrixmetalloproteinases (collagenase, for instance) are often assumed to be particularly responsible for that degradation processes [35], especially in combination with ROS. This opinion is emphasized by the observation that the number of granulocytes is augmented in the blood from RA patients [36] and that polymorphonuclear granulocytes (abbreviated as both PMNs or neutrophils) can be found in large numbers in the pannus tissue, a replacement tissue as a consequence of ECM degradation, of cartilage [37,38].
Although macrophages [39] and T-cells [40] may also contribute to cartilage degradation, this review will focus on products derived from neutrophils (most potent inflammatory cells), which are characterized by much higher concentrations of MPO than other cells [41,42]. The main reason why the contribution of neutrophils was underestimated to date is—at least partially—the fact that neutrophils cannot be kept in cell culture or only with considerable limitations (for example, as “neutrophil-like” cells [43]). Thus, they must be freshly prepared from the blood from volunteers [44] that confers many difficulties, particularly the poor reproducibility of the investigations. Recently, the “neutrophil extracellular traps” (NETs) were introduced as important constituents in RA [45] which will be discussed in Section 9 of this review (vide infra).
Normally, there is an equilibrium between the generation of ROS/reactive nitrogen species (RNS) and their consumption by antioxidants [46]. However, excessive ROS generation leads to oxidative induced post-translational modifications of proteins and may give rise to neoepitopes that are recognized by the immune system as non-self-substance. The resulting formation of autoantibodies is detectable by specific antigens, which might improve both early diagnosis and monitoring of disease activity. Particularly promising diagnostic autoantibodies include, for instance, anti-carbamylated proteins (generated by the reaction with a degradation product of urea) and anti-oxidized type II collagen antibodies [47].
4. Reactive Oxygen/Nitrogen/Chlorine Species
Such reactive species are generated in or (after the release of the required enzymes) from many cells [48]. For instance, fibroblasts, chondrocytes, macrophages, and, particularly, neutrophils are known as sources of ROS [49]. Neutrophils accumulate in inflamed joints in huge amounts [50,51] with an average percentage in the whole SF as high as 85.7% [52]. The physiological contribution of neutrophils (compared to the total cell number) is, however, much smaller.
Although pathogenic microorganisms are involved in some kinds of arthritis, antibodies, cytokines and chemotactic agents are assumed to represent one of the prime reasons for the accumulation and activation of neutrophils in the inflamed joint [36]. The increased oxygen consumption by neutrophilic granulocytes upon stimulation is called respiratory burst [53] and the generated ROS, RNS and reactive chlorine species (RCS) are summarized in Figure 2. These reactive species contain either oxygen, nitrogen, chlorine [54]—or even two of these (electronegative) elements. In some cases, the nomination is thus ambiguous: for instance, NO2 is commonly considered an RNS. However, it also contains oxygen. Therefore, ROS would be an appropriate term, too. We will use ROS throughout this review because it includes most of the relevant species—particularly HOCl.
ROS were comprehensively discussed already in the past [8]. Thus, we will only discuss those species, which are particularly relevant in the context of MPO and start with atmospheric oxygen (O2), which constitutes about 21% of the air. In a first reaction (Equation (1)), the enzyme NADPH oxidase catalyzes the reduction of O2 into superoxide anion radicals (O2•−), at which point the required electrons are generated by the oxidation of NADPH by the enzyme NADPH oxidase [55]:
Afterwards, O2•− dismutates either spontaneously or particularly in the presence of the enzyme superoxide dismutase (SOD) into hydrogen peroxide (H2O2) and O2 (Equation (2)):
H2O2 is the substrate of the enzyme MPO, which is particularly abundant in neutrophils, monocytes [56] and macrophages [57]. H2O2 is the educt of more efficient oxidants [58]. MPO reduces H2O2 to water under the formation of the so-called compound I (Figure 2). This activated form of the enzyme is reduced to the native enzyme either by abstracting two electrons from (pseudo)halides or by two one-electron steps via the formation of compound II [59]. In the first case, (pseudo)halides such as SCN− are oxidized to (pseudo)hypohalous acids. The generation of hypochlorous acid (HOCl) and hypothiocyanate (OSCN−) is particularly important under physiological conditions [60]. The most important reaction can be summarized as follows:
MPO is stored in the azurophilic (primary) granules within the neutrophils. The release of MPO may occur either into phagosomes containing engulfed pathogens or into the extracellular space, depending on the respective stimulus [61]. The most common degranulation involves the release of primary granules mainly into the phagosome, which presumably helps to prevent excessive tissue damage. However, in particular inflammatory conditions or in response to specific pathogens, larger moieties of MPO may be released extracellularly. This involves the fusion of granule membranes with the phagosome or the plasma membrane, enabling the release of granule contents into the extracellular space or phagosome [62], which is surprisingly accompanied by a significant change in the pH: immediately after phagosome formation, the pH rises to alkaline values due to NADPH oxidase activation. This pH change may facilitate MPO release and enhance MPO activity [63].
More recently, MPO degranulation was reported to be mediated via the extrusion of NETs [64], which are also discussed in this review (vide infra). It should be noted that controlled MPO release at the site of infection is of paramount importance for optimum activities. Any uncontrolled degranulation exaggerates the inflammation and may lead to unwanted tissue damage even in the absence of inflammation.
Furthermore, the determination of the MPO activity is one of the best diagnostic tools of oxidative stress biomarkers in arthritic diseases. Important activators and inhibitors of MPO are discussed in [61]: since high amounts of MPO (in the blood) are a decisive factor for early death [65], the development of suitable MPO inhibitors is of paramount relevance regarding the cure of arthritis.
Other halogenides such as Br− are even more efficiently converted by MPO. However, hypobromous acid (HOBr) does regularly only play a minor role because it is by far less abundant than [66] at physiological conditions. Since HOCl is continuously generated by the system, but immediately consumed by its reaction with (abundant) biomolecules, the assessment of the HOCl concentration under in vivo conditions is difficult. Accordingly, the in vivo HOCl concentration is presumably very low. However, some authors provided (significantly varying) HOCl concentrations. For instance, 50–100 mM were reported under inflammatory conditions [67], while 0.34 mM was reported as the HOCl concentration in the extracellular space [68]. The determination of the HOCl concentration in physiological systems is nowadays a hot topic [69] and there are many reports describing new methods of HOCl quantitation in physiological systems.
MPO is an abundant enzyme in neutrophils and constitutes about 5% of the entire protein mass within neutrophils [70]. Therefore, both MPO and HOCl as its prime product have been assumed to be massively involved in the degradation of the polymeric components (particularly GAGs of the proteoglycans) of cartilage for many years [68]. This aspect was confirmed by the fact that characteristic degradation products of cartilage (acetate and oligosaccharides) could be detected in the inflamed joint fluids from patients suffering from RA [31]. The contribution of neutrophils and products derived thereof in cartilage degradation has been recently reviewed [36]. Physicians recommend slight physical activity to their patients suffering from rheumatic diseases. In fact, physical activity may trigger the generation of MPO and, thus, the extent of HOCl generation [71].
5. Release of Enzymes into the Synovial Fluid—A Short Survey
The presence of MPO in SF was—to the best of our knowledge—first described in 1979 [72]. Hadler and coworkers determined the concentrations of several neutrophil-derived lysosomal proteins by immunochemical and enzymatic assays in 28 inflammatory and 9 non-inflammatory SF samples. The quantities of lactoferrin, MPO and enzymatically determined lysozyme correlated with the number of neutrophils. In contrast, enzymatic activities measured with synthetic substrates developed for the assay of chymotryptic-like cationic protein (cathepsin G) and elastase, along with immunochemically determined lysozyme, were independent of the number of neutrophils. The elastase activity (which was determined with elastin as substrate) was close to zero. A negative correlation between the concentration of common proteases and the degree of radiographic destruction of the joint could be observed. Cathepsin G and elastase are stored in an active form in neutrophil azurophilic granules [73]. There are two possible explanations why these proteins could not be detected: On the one hand, both enzymes may be missing and/or they are potentially not released by the primary granules of the neutrophils. On the other hand, inhibitors such as Thrombospondin 1 may considerably reduce the activities of these enzymes [74]. It seems likely that the elastase-alpha 1 proteinase inhibitor complex (EIC) plays a major role: for instance, the EIC levels increased according to the stage of articular cartilage destruction and the activity of neutrophil elastase was elevated in destructive joints of RA patients [75]. With the progression of articular cartilage destruction, the EIC levels in plasma of RA patients increased as well.
In a nutshell, the role of proteases is overstated in many studies in comparison to ROS. This can be understood by the used methods: proteases give defined reaction products, which can be rather easily determined, for instance, by mass spectrometry. Compared to that, the determination of ROS is much more difficult since the obtained products represent often transient products (e.g., chloramines).
6. Function and Modifications Induced by MPO
MPO plays a major role in the killing of invaded microorganisms by generating a particular ROS: compound I of MPO oxidizes chloride anions to HOCl, a strong oxidizing and chlorinating species (Figure 2) [76]. HOCl is a very weak acid with a pKa value of 7.53 [77]. At physiological conditions (pH = 7.4) there is, thus, a nearly 1:1 molar ratio between HOCl and the hypochlorite ion (ClO−). Although contributions of ClO− cannot be ruled out at physiological conditions, there is nowadays an agreement that HOCl is the mainly relevant oxidizing species. Considering these aspects, one observation made in daily practice is somewhat strange: the product yield increases if the pH is lowered. Since HOCl should be reformed (according to Le Chatelier’s principle) from ClO−, a marked dependence on the pH could not be expected.
Dichlorine monoxide (Cl2O) may also be regarded as the anhydride of HOCl (Equation (4)) [78], which is increasingly assumed (in the same manner as chlorine gas [79]) to be involved in the deleterious actions of HOCl [78]. However, potential effects of Cl2O in the field of arthritis or cartilage degradation have not been investigated to date.
HOCl is an important source of other, more reactive ROS, such as hydroxyl radicals (, Equation (5)) or singlet oxygen (, Equation (6)):
Converting nitrogen dioxide (NO2) into nitryl chloride (NO2Cl) by HOCl [80] may lead to serious modifications of the sidechains of proteins [81]. However, a detailed discussion of these aspects is beyond the scope of this review.
HOCl reacts with virtually all biomolecules, i.e., with amino acids, nucleic acids, sugars and lipids (Figure 3) [82] but with strongly different velocities. Comparative data of the corresponding reaction kinetics are available in different publications [83]. Thiols (as in cysteine) and thioethers (as in methionine) are the preferred targets of HOCl (indicated by the thick arrow in Figure 3).
After all thiol and thioether groups are consumed, other functional groups, such as amino functions in sugars, are affected by HOCl [84], i.e., HOCl undergoes well-defined, gradual reactions. The second order rate constants of the reactions between HOCl and thiols account for about 107 M−1 s−1 [85] and with amino groups about 7 × 104 M−1 s−1 [86], while the second order rate constants with olefinic residues are much slower and comprise only about 9 M−1 s−1 [87]. More detailed data were also compiled by Panasenko and coworkers [88] and by Davies et al. [89], and a survey of the data is available in Figure 4.
We will now discuss the reactions between HOCl as the main product of the enzyme MPO and the most important constituents of cartilage and SF, i.e., collagen, GAGs, DNA and lipids.
7. Reactions between HOCl and Cartilage Components
7.1. Collagen
The reaction between ROS and collagen was by far less comprehensively studied compared to the enzymatic degradation of collagen (see Section 2). Nevertheless, it is well known that the in vitro reactivity between selected amino acids and O2•− or H2O2 (as the substrates of MPO) is poor [90]. Slight reactivity is exclusively observed at low pH values, at which particularly sulfur containing amino acids and (to a minor extent) aromatic amino acids are oxidized [90]. However, both are not very abundant in collagen.
Despite this poor reactivity, evidence has been provided that O2•− producing systems contribute to tissue and cartilage degradation. At these conditions, solubilized collagen (isolated from skin or cartilage) can be assessed by an increased concentration of 4-hydroxyproline in the supernatant of the otherwise insoluble material. Similar results were also obtained upon incubation of cartilage specimens [91]. In addition to collagen degradation, GAG degradation (vide infra) was also monitored by an increase in the uronic acid concentration. Selective scavengers of O2•− (SOD or catalase) decreased the extent of cartilage fragmentation. Despite these results, it can be assumed that O2•− plays only a minor role in cartilage destruction due to its poor reactivity. It is much more likely that O2•− is converted into a more deleterious species (particularly HO•) at pathological conditions. It is also reasonable to assume that iron ions are massively involved in these processes [92]. Nevertheless, there is a complex interplay between relevant enzymes such as SOD and ROS [93,94].
Although the reaction of HOCl and molecules like amino acids or carbohydrates is much faster compared to O2•− or H2O2, there is no consensus about the HOCl-mediated effects on the collagen moiety of cartilage to date. On the one hand, it was shown that the collagen moiety in the cartilage is affected by HOCl to a lesser extent than the polysaccharides of articular cartilage [95]. Similar results were obtained when other types of cartilage, e.g., bovine nasal cartilage with an enhanced GAG content but a reduced collagen content, were treated with HOCl [96]. This is a clear indication that elevated concentrations of HOCl are required to affect the collagen moiety of cartilage. On the other hand, it became evident that HOCl activates collagenase, which may subsequently lead to damages of collagen [97]. This is presumably mediated by the modification of collagenase inhibitors, not the enzyme itself. In contrast, taurine chloramine (one of the very few stable chloramines) had the opposite effect and led to an inhibition of the collagenase activity [98]. Stamp and coworkers [99] found elevated protein carbonyl and 3-chlorotyrosine concentrations along with an increased MPO activity in the SF of RA patients. Since only MPO is able to generate 3-chlorotyrosine, the detection of this molecule is a strong hint on the production of HOCl. Of course, 3-chlorotyrosine is not necessarily derived from collagen but might also stem from the link or the core protein of the proteoglycans.
Odobasic and coworkers [100] explored the role of endogenous MPO in experimental (collagen-induced arthritis) RA in normal and MPO knockout mice. They found that MPO contributes to the development of arthritis: MPO enhanced the proliferation and decreased the apoptosis of synovial fibroblasts in vitro.
Westman and coworkers found that the chlorination of collagen type II (the most abundant collagen of cartilage [101]) might represent a mechanism by which immunoreactivity is induced and by which chronic joint inflammation is supported. However, the detailed characterization of the chlorinated collagen was not performed which is a significant weakness of this study [102]. Interestingly, physiologically relevant concentrations of HOCl (between about 5 and 50 μM) lead to the degradation of collagen [103] and reduce the gel-forming tendency of collagen. Similar effects are induced by chloramines (e.g., from amino acids), if they are used instead of the reagent HOCl. It was also suggested that hypochlorite, N-chloramines, and chlorine are involved in the oxidation of the pyridinoline cross-linkages within the collagen type II in articular cartilage during acute inflammation [104]. Somewhat later, the collagen type II modification by hypochlorite was investigated in more detail [105]. The authors found that chlorination decreases the radius of collagen II aggregates from 30 to 6.8 nm. Since this alteration already occurs at low concentrations of HOCl, changes in the aggregate size were suggested as the optimum markers of HOCl-induced collagen oxidation. In another study it was elucidated that the in vitro oxidation of collagen promotes the formation of advanced protein oxidation products and is involved in the activation process of human neutrophils [106]. Although cartilage-related collagen was less comprehensively studied, there is increasing evidence [66] that thyroid peroxidase and peroxidasin are key enzymes for thyroid hormone synthesis. This is also important regarding the establishment of functional cross-links in collagen IV during basement membrane development. Although less abundant in cartilage than collagen type II, the collagen IV network plays a crucial role regarding the mechanical integrity of the basement membrane. A key event represents unequivocally the formation of intra- and inter-collagen fibril crosslinks. An inter-residue sulfilimine bond, which does otherwise rarely occur, was reported to be unique by collagen IV. These crosslinks are particularly formed between lysine/hydroxylysine or methionine residues and might occur inter- and intrafibrillarly.
Due to its significance as the major crosslink in the collagen IV network, the sulfilimine bond plays a critical role in tissue development and various human diseases [107]. In that way, HOCl and particularly HOBr are very important [108]. The basic reactions are illustrated in Figure 5:
At sites of inflammation, such as the SF of RA patients, increased RNS-mediated protein damage has been detected in the form of a biomarker, 3-nitrotyrosine, by immunohistochemistry, Western blotting, ELISAs and MS. Further details are available in [109].
7.2. GAGs
There is a consensus that GAGs are more efficiently degraded by HOCl in the diseased joint than the collagen moiety [110]. This is caused by the poor solubility of collagen and its triple-helical structure, which aggravates the attack by ROS.
It has been established that the reagent HOCl (as well as the entire MPO/H2O2/Cl− system) reduces the viscosity of solutions of high-molecular-weight hyaluronan. The term hyaluronan is a mixture of HA and hyaluronate which indicates that all species are included—independent of the charge state [111]. Baker and colleagues used size exclusion chromatography to evaluate the effects of either exogeneously added HOCl or the entire MPO/H2O2/Cl− system on aqueous hyaluronan solutions. It could be shown that already µM concentrations of HOCl (HA was in the mg/mL range which corresponds—related to the molecular weight of the polymer repeat unit—to a mM concentration) reduced the viscosity of HA. In contrast, elevated concentrations of HOCl were necessary to reduce the molecular weight of the HA polysaccharide [111], i.e., to induce scissions of the glycosidic linkages between the individual monosaccharide units. This was explained by structural modifications of the HA polysaccharide occurring (in contrast to the cleavage of chemical linkages) already in the presence of moderate HOCl concentrations and is accompanied by the reduction in the viscosity of HA. A few years later an interesting phenomenon was reported [30]: HOCl/MPO depolymerizes only purified umbilical cord HA (in a HO•-dependent way) but does not depolymerize the HA polysaccharide in SF. The authors concluded that HOCl/MPO has a scavenging action on SF HA by the consumption of H2O2. In this way, the formation of HO• radicals is suppressed. Therefore, aqueous HA solutions are not necessarily comparable with native SF since the aqueous solutions do not contain proteins [112].
The determination of the viscosity of (pathologically changed) SF was (and still is) an established method of the severity of inflammatory joint diseases after puncture of the affected joint [113,114]. Later studies emphasized the role of the proteins in biological materials—even if HA is still considered as a powerful mean to attenuate the oxidative stress in the inflamed joint: the injection of native HA into the joint is known to have several beneficial effects [114,115]. These studies were performed by monitoring potential effects of the inflammatory state by the measurement of some indicative effects such as the HA concentration and the protein composition on the SF viscosity [116].
The mechanism of HA degradation was studied by 1H nuclear magnetic resonance (NMR) spectroscopy in 1994. Both N-acetylglucosamine (GlcNAc) and chondroitinsulfate (CS) were separately treated with the reagent HOCl and the kinetics of the reaction monitored [117]. Since amino groups are particularly reactive, HOCl reacts first with the GlcNAc or the amino sugar within the GAG polysaccharide (Figure 6).
Two consecutive effects were observed: (a) the depolymerization of the GAG polysaccharide chain under scission of the glycosidic linkages; and (b) the cleavage of the N-acetyl groups via a transient, presumably chlorinated product. This product and the involved N-centered radicals could be later monitored by electron spin resonance and other methods [118]. Similar observations were made if isolated hyaluronan or HA from SF was subjected to γ-irradiation [119]: this leads to the scission of the water molecule under generation of HO• radicals, which are capable of depolymerizing the HA molecule. Similar data may also be obtained if the radicals are generated via Fenton chemistry, which may also occur under in vivo conditions [120].
The potential contribution of N- and O-centered free radicals during the HOCl-induced degradation of HA is illustrated in Figure 7:
The relative reactivities of selected functional groups within the carbohydrates can also be determined by measuring the HOCl consumption: the required time-dependent data can be easily obtained by UV spectroscopy since the hypochlorite anion is characterized by an intense UV absorption (ε290 = 350 M−1cm−1). While different amino sugars possess significant reactivities, glucose or glucuronic acid do not react at all with HOCl [122].
Although there were no attempts to detect carbon dioxide (CO2) as a final oxidation product of HA and other carbohydrates to date, intermediate oxidation products such as formate are exclusively detected if a massive excess of HOCl is used. Since formate is an established product of HO•-induced carbohydrate degradation [120], this implies the conversion of HOCl into more reactive ROS (Equation (7)) [123].
Akeel et al. [124] studied the in vitro reaction of HOCl with either HA or heparin (an agent against the consequences of arthritis [125]) using spectrophotometrically and enzymatically based methods. These authors found differences in dependence on the extent of sulfation of the involved carbohydrates. Although not yet completely clarified, the electron-withdrawing (-I) effect of the sulfate groups may play a significant role because it has an impact on the electron density at specific positions within the carbohydrate molecule.
It is very surprising that mass spectrometry (MS) has only been scarcely used to investigate the reaction between ROS and carbohydrates to date. Since oxidation is accompanied by changes in the molecular weight, MS represents a reliable method to monitor the ongoing changes. One of the available reports to date used electrospray ionization (ESI) MS to monitor the ROS-induced degradation of HA [126]. Unfortunately, this study gave only rather limited information but a clear indication that the investigation of oxidized carbohydrates by MS is even nowadays a challenging topic [127]. This is partially caused by the fact that sugars are rather refractive to the ionization process and give limited ion yields. This problem increases if the polarity of the carbohydrates increases, for instance by the introduction of sulfate groups [128].
In a more recent study, the chloramide of native, high-molecular-weight hyaluronan could be successfully synthesized. However, chlorinated isocyanuric acid had to be used as the chlorinating agent [129] because HOCl resulted in poor yields. In a very recent study, the degradation of HA and selected oligo- and monosaccharides by HOCl was studied by ESI MS in combination with thin-layer chromatography (TLC) [122]. It was demonstrated that the MS-based detection of N-chlorinated GAG amides is challenging while cleavages of the glycosidic linkages and the generation of chloramines in oligosaccharides are readily detectable. This study also provided evidence that the 1→4- and 1→3-glycosidic linkages exhibit different reactivities with HOCl [122].
In a previous study, pig articular cartilage was treated with different amounts of HOCl (at pH 7.4) and the composition of the supernatants assessed by 1H NMR [95]. Since collagen is very rich in glycine, the denaturation of collagen can be estimated by using the resonance at 3.55 ppm. However, this resonance was very weak and there were also no chlorinated products of glycine detectable [130]. In contrast, the intensities of GAG oligosaccharides (2.04 ppm) as well as acetate (1.90 ppm) were elevated subsequent to HOCl treatment of cartilage. This is a clear indication that the GAGs of cartilage are depolymerized by HOCl. Nevertheless, chlorinated peptides were also reported to be indicative of MPO activity [131]. These peptides are presumably not derived from the collagen but are stemming from other proteins, e.g., the core or the link protein of proteoglycans.
NMR is also a convenient method to discriminate SF from patients suffering either from OA or RA [132]. The particular advantage of NMR is that no prior knowledge about the sample composition is necessary.
Hawkins and Davies [133] made an interesting suggestion: MPO is a strongly cationic protein (positively charged, isoelectric point (pI) ≈ 10). In contrast, all GAGs within the cartilage are negatively charged, particularly the sulfated ones because sulfate represents a strong electrolyte. Due to the attraction of the differently charged molecules, it is likely that HOCl is generated in the vicinity of the GAGs. The short distance between MPO and/or the released HOCl may be one important reason why GAGs (similarly as negatively charged phospholipids [134]) represent one of the first targets of HOCl. This might explain the recent focus on NETs (vide infra).
7.3. DNA
Although it is well known that DNA reacts with HOCl [135,136], very little is known about this reaction in the context of RA and OA. Karaman and coworkers [137] reported DNA damage in RA lymphocytes in parallel with an increase in malondialdehyde (MDA) levels and decreased activities of SOD and glutathionperoxidase (GPx), which are both enzymes with antioxidative properties. These data emphasize again the increased oxidative stress in RA. A few years later, it was shown that polymorphisms of DNA damage repair genes play a role in RA pathogenesis. Accordingly, these DNA polymorphisms might be useful as RA disease markers [138]. Although a more detailed discussion of these aspects is beyond the scope of this review, it was very recently suggested that one hallmark of RA is impaired DNA repair observed in patient-derived peripheral blood mononuclear cells [139].
7.4. Cellular Lipids
Since the cartilage tissue contains only comparably small amounts of lipids [141], the main source of lipids comes from the cells. Even if the cell density is rather poor (cartilage is known as a bradytrophic tissue) all cells contain many lipids (particularly phospholipids) in their cellular membranes and organelles [142]. In order to warrant a sufficient extent of membrane fluidity, the majority of membrane lipids contain double bonds [143]. Second order rate constant of HOCl against double bonds is rather poor. Nevertheless, lipids are affected due to their comparably high concentration in membranes [142].
As already indicated, amino-phospholipids such as phosphatidylethanolamine (PE) or -serine (PS) exhibit the most pronounced reactivity with HOCl [144]. However, there are no reports on chlorinated PE within the cartilage to date, although there is a very recent study on lipid markers between infrapatellar fat pad biopsies of OA and cartilage defect patients using MALDI MS Imaging. The main products between unsaturated lipids of the phosphatidylcholine (PC) type and HOCl are chlorohydrins, i.e., addition products of HOCl onto the double bond(s) [145]. Although this reaction has been well established for more than 30 years [146], it has to be emphasized that there are also free fatty acids due to the natural turnover of phospholipids in all biological systems. These free fatty acids react with HOCl under the formation of chlorohydrins as the main products as well [147]. There are indications that these products are converted into dimeric and trimeric products [148] with particular properties. Non-chlorinated (or in general non-halogenated) fatty acid esters of hydroxy fatty acids (FAHFA) are assumed to possess significant physiological relevance but are commonly generated by different pathways under the involvement of different enzymes [149]. Although it is well known that lipid chlorohydrins decrease the stability of membranes of erythrocytes [150], there is nothing known about the toxic effects of lipid chlorohydrins or chloramines on cartilage cells to date. However, in 2002, it was shown that 1,3-dichloro-2-propanol (1,3-DCP), an abundant impurity in many compounds like hard resins or celluloid, represents a major health concern [151].
Nitrite is known to be present in concentrations of up to 4 mM in SF from patients suffering from RA [152]. Therefore, it is likely that nitrite reacts with HOCl under generation of nitrate (Equation (8)).
Thus, the concentration of HOCl (vide supra) is reduced. This was first shown using the reaction of isolated lipids with HOCl [153]. Afterwards, it was also confirmed by systems mimicking the real conditions within the inflamed joint [154]: it could be convincingly shown that the presence of nitrite (but not nitrate) decreased HOCl-dependent cellular toxicity even if very small amounts of nitrite (in the µM range) were used. In contrast, nitrite (even in higher concentrations) did not inhibit superoxide-, hydroxyl radical-, hydrogen peroxide-, or peroxynitrite-mediated cytotoxicity. This is a clear indication of the different reactivities of the individual species. Additionally, the oxidation of plasmalogens by HOCl may also play a major role. These lipids are not characterized by two ester linkages (as it is the case in common lipids) but possess one ester and one alkenyl-ether linkage [155]. These lipids are relatively scarce in the SF [156] but give very characteristic oxidation products upon the reaction with HOCl since the resulting 2-chlorfatty aldehyde [157] has a high reactivity with many other functional groups, particularly amino residues.
8. Antioxidants to Suppress HOCl-Induced Effects
HOCl is a molecular (non-radical agent) with defined gradual reactivity: first, sulfhydryl groups are oxidized, and, afterwards, amino groups are converted into mono- or dichloroamines depending on the stoichiometry. The double bonds of lipids have the lowest reactivity and chlorohydrines are, thus, generated with the lowest velocity [133]. However, the reaction between HOCl and common antioxidants such as trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid, an analogue of vitamin E) or ꞵ-carotene is faster compared to the majority of other biomolecules [89]. Compounds with reactive -SH or -S-S- groups such as lipoic acid [159,160] can be considered as effective antioxidants against HOCl although it is not yet completely clear whether the generated products are chemically inert and do not induce further reactions. Furthermore, different antioxidants were recently shown to be effective agents in the treatment of arthritis [161].
Taurine (2-aminoethanesulfonic acid) is often discussed [162] as the most important antioxidant against HOCl: the reaction between taurine and HOCl is slower compared to thiol groups but the products are characterized by significant stability [163]. This is important because there are also papers where the pharmacological aspects of the chloroamines are discussed. First, it was shown that the local administration of N-chlorotaurine represents an inhibitor of septic arthritis [164]. Additionally, N-chlorotaurine is discussed as a suitable topical (applied onto the skin) anti-infective. There is currently some research in order to develop novel mono- and dichloro- derivatives of dimethyltaurine, which are assumed to possess improved stability [165]. Although the pharmacological effects of well-known antirheumatic drugs such as methotrexate [166] and paracetamol [167] are surely different from the exclusive scavenging of HOCl, both are also known to react with HOCl. Therefore, one potential aspect might be the scavenging of HOCl.
The selective inhibition of MPO in the joint or the cartilage is unequivocally an effective way to suppress its effects and/or the generation of its most important product, HOCl. Compounds such as thioxanthines [168] or the KYC peptides (e.g., N-acetyl-lysyl-tyrosyl-cysteine amide) [169] seem to fulfill the corresponding needs. However, the long-term inhibitor use might be dangerous for the immune system. Many physicians are concerned about this problem and more research is needed in this field. The thioxanthines, for instance, have been shown to reduce, but not to prevent, the efficient killing of Staphylococcus aureus by neutrophils [170] and are, thus, potentially suitable for in vivo use. This aspect has been recently reviewed [171].
The fact that a knockdown of MPO is not lethal (the majority is clinically asymptomatic except if they are also diabetic [172]) may have two different reasons. Either residual MPO activity is sufficient for bactericidal activity or other enzymes can compensate for the inhibition of MPO: thioxanthines inhibit thyroid peroxidase and lactoperoxidase to a much lesser extent than MPO as assessed by in vitro assays [170]. In contrast, detailed studies with the KYC peptides are still lacking in the field of arthritis. Since chloride ions are substrates of MPO, reducing the intake of salt may have positive effects for arthritis patients: it was suggested that increased bone erosion under high-salt conditions can be attributed to an enhanced oxidative milieu maintained by infiltrating neutrophils [173].
Due to the incidence rate of RA and the limitations of reliable therapies, the search for new treatment strategies for RA became a transnational research focus. Specifically, ferroptosis is a novel type of programmed cell death characterized by iron-dependent lipid peroxidation. Ferroptosis is characterized by distinct differences from apoptosis, autophagy, and necrosis—and it may be triggered by ROS [174,175].
9. Neutrophil Extracellular Traps (NETs)
The fusion of both antimicrobial granules and the phagosome is an important event of neutrophils to kill invasive bacteria. A closely related new mechanism of neutrophils—called neutrophil extracellular traps (NETs)—was first described in 2004 [176]. The authors observed the release of different granule proteins and chromatin (simplified a mixture of DNA and basic proteins) by neutrophils upon in vivo activation by, e.g., lipopolysaccharides in order to attack pathological microorganism. Both compounds form extracellular fibers that are essential for the binding of bacteria and their subsequent killing.
Since this initial observation, the term and the discussion of NETs have been hot topics. NETs are defined as extracellular structures containing DNA, histones, and neutrophil-characteristic proteins such as MPO or elastase, which are secreted from the neutrophils in response to inflammatory stimuli. Additionally, neutrophils undergo chromatin remodeling with the subsequent release of the decondensed chromatin from the cells under generation of a network, illustrated in Figure 9.
Although many details of this mechanism are unknown to date [178], it is commonly accepted that NETs are produced in inflammatory areas and possess only a short half-life due to their fast enzymatic degradation [179]. Since a detailed discussion of all these events is clearly beyond the scope of this review, interested readers are advised to consult one of the more recent reviews [177]. Nevertheless, it is of interest to note that NADPH oxidase interactions with MPO are presumably involved which eventually lead to stimulation of neutrophil elastase. This is accompanied by the degradation of histones, chromatin decondensation and NET release.
This “suicidal NETosis” represents a new type of cell death in addition to apoptosis and necrosis [180] and with some peculiarities [181]. NETosis is nowadays assumed to represent a defense mechanism that is activated in response to the presence of inflammatory stimuli. In a nutshell, such processes are currently of huge interest regarding the development of antirheumatic drugs and the treatment of the disease in general [182]. Since our main interest is on the chemical aspects of cartilage degeneration, we will not delve into further details. However, a very crude survey of these processes is illustrated in Figure 10.
The role of NET detection in monitoring the severity of disease as well as the effect of different anti-rheumatic therapies is an important research topic. It seems that NETs play a major role in the pathogenesis of the disease and are not just a secondary finding associated with the inflammatory process. Further mechanistic processes (including immunological details) are available in [51,184]. It was also shown that neutrophils from RA SF drive inflammation through the production of chemokines, ROS, and NETs [51]. Inhibiting the formation of excessive NETosis, reducing the release of pro-inflammatory biological mediators, and converting the death form of neutrophils into a safer form of cell death represents presumably a key event. Finally, it is important to note that NETs are not only relevant in joint diseases but are also involved in important diseases such as atherosclerosis, sepsis and COVID-19 [185]. The available research literature in this field (between 1985 and 2023) has been recently reviewed [37].
10. Conclusions
Although clinically very different, RA and OA represent the most common musculoskeletal diseases, at which the synovia or the bone/cartilage is damaged at OA or RA conditions, respectively. One characteristic of OA is the degeneration of the articular cartilage, which is initiated by malfunctions of cartilage cells after joint injury. The degradation is additionally accelerated by inflammation, i.e., the generation of ROS and the release of tissue-degrading enzymes. The degeneration triggered by these biomechanical and biochemical mechanisms is irreversible. Although a lot of work was performed in the past, the complex mechanisms of cartilage degradation are insufficiently understood to date. One particular problem regarding the interpretation of the obtained data is the limitation of many studies on a particular compound class and/or a single degeneration-causing agent.
In particular, cartilage degeneration mechanisms on the cellular level have only been scarcely discussed to date, although there are indications that early post-traumatic biomechanical and inflammatory effects on cartilage cells impact the composition of cartilage [186]. Stimulated macrophages or neutrophils have only been scarcely used to mimic the “real” events of cartilage degradation to date. Since the events of cartilage degradation are very complex, there is an increasing number of in vitro models. For instance, it was suggested that the mitigation of post-injury inflammation leads presumably to the recovery of cartilage composition.
MPO is assumed to be massively involved in inflammatory joint diseases [99] for the following reasons:
- The number of neutrophils is massively enhanced in the SF from patients suffering from RA [187].
- Since MPO is a very abundant protein within the neutrophils (it constitutes about 5% of all proteins), the contribution of this enzyme and its product, HOCl, to cartilage degradation during RA is obvious.
Thus, MPO seems to be an effective mediator in the development of rheumatic diseases, presumably accompanied by necrosis of the cartilage cells. Necrosis is also suggested to result in the release of damage-associated molecular patterns (DAMPs) and pro-inflammatory cytokines [189], leading to ECM degeneration caused by proteolytic enzymes [190]. Apoptosis, controlled cell death, has also been associated with excessive production of ROS [191]. Excessive ROS production has been suggested to promote ECM degeneration via decreased matrix biosynthesis [192]. However, further studies to improve the treatment of RA and OA are needed.
Author Contributions
J.L. and J.S. provided the original manuscript draft preparation. The supervision, project administration and funding acquisition were the tasks of J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the German Research Council (SCHI 476/19-1, 26-1 (531487925) and SFB 1052/Z3), and by the Open Access Publishing Fund of Leipzig University.
Acknowledgments
We would also like to thank all colleagues who helped during the writing process.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mela, G.S.; Cimmino, M.A. An overview of rheumatological research in the European Union. Ann. Rheum. Dis. 1998, 57, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Pincus, T.; Castrejon, I.; Yazici, Y.; Gibson, K.A.; Bergman, M.J.; Block, J.A. Osteoarthritis is as severe as rheumatoid arthritis: Evidence over 40 years according to the same measure in each disease. Clin. Exp. Rheumatol. 2019, 37 (Suppl. 120), 7–17. [Google Scholar] [PubMed]
- Lawrence, R.C.; Helmick, C.G.; Arnett, F.C.; Deyo, R.A.; Felson, D.T.; Giannini, E.H.; Heyse, S.P.; Hirsch, R.; Hochberg, M.C.; Hunder, G.G.; et al. Estimates of the prevalence of arthritis and selected musculoskeletal disorders in the United States. Arthritis Rheum. 1998, 41, 778–799. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.; Chan, L.; Flynn, S. A Systematic Review of the Incidence, Prevalence, Costs, and Activity and Work Limitations of Amputation, Osteoarthritis, Rheumatoid Arthritis, Back Pain, Multiple Sclerosis, Spinal Cord Injury, Stroke, and Traumatic Brain Injury in the United States: A 2019 Update. Arch. Phys. Med. Rehabil. 2021, 102, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Elkouzi, A.; Bit-Ivan, E.N.; Elble, R.J. Pure akinesia with gait freezing: A clinicopathologic study. J. Clin. Mov. Disord. 2017, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Flugge, L.A.; Miller-Deist, L.A.; Petillo, P.A. Towards a molecular understanding of arthritis. Chem. Biol. 1999, 6, R157–R166. [Google Scholar] [CrossRef]
- Thoms, B.L.; Bonnell, L.N.; Tompkins, B.; Nevares, A.; Lau, C. Predictors of inflammatory arthritis among new rheumatology referrals: A cross-sectional study. Rheumatol. Adv. Pract. 2023, 7, rkad067. [Google Scholar] [CrossRef]
- Schiller, J.; Fuchs, B.; Arnhold, J.; Arnold, K. Contribution of reactive oxygen species to cartilage degradation in rheumatic diseases: Molecular pathways, diagnosis and potential therapeutic strategies. Curr. Med. Chem. 2003, 10, 2123–2145. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Fuchs, B.; Arnold, K. The Molecular Organization of Polymers of Cartilage in Health and Disease. Curr. Org. Chem. 2006, 10, 1771–1789. [Google Scholar] [CrossRef]
- Rich, M.J.; Burnash, S.; Krishnan, R.R.; Chubinskaya, S.; Loeser, R.F.; Polacheck, W.J.; Diekman, B.O. Use of a novel magnetically actuated compression system to study the temporal dynamics of axial and lateral strain in human osteochondral plugs. J. Biomech. 2024, 162, 111887. [Google Scholar] [CrossRef] [PubMed]
- Bursac, P.; Arnoczky, S.; York, A. Dynamic compressive behavior of human meniscus correlates with its extra-cellular matrix composition. Biorheology 2009, 46, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, R.A. The cell density of human articular and costal cartilage. J. Anat. 1967, 101, 753–763. [Google Scholar]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The extracellular matrix: Tools and insights for the “omics” era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, Y.; Grodzinsky, A.J. Cartilage diseases. Matrix Biol. 2018, 71–72, 51–69. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, G. Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 2018, 50, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Krane, S.M. The importance of proline residues in the structure, stability and susceptibility to proteolytic degradation of collagens. Amino Acids 2008, 35, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.I.; Li, Y.; Pan, J.; Liu, F.; Dai, H.; Fu, Y.; Huang, T.; Farooq, S.; Zhang, H. Collagen and gelatin: Structure, properties, and applications in food industry. Int. J. Biol. Macromol. 2024, 254, 128037. [Google Scholar] [CrossRef]
- Nimni, M.E. Collagen: Structure, function, and metabolism in normal and fibrotic tissues. Semin. Arthritis Rheum. 1983, 13, 1–86. [Google Scholar] [CrossRef] [PubMed]
- Knauss, R.; Schiller, J.; Fleischer, G.; Kärger, J.; Arnold, K. Self-diffusion of water in cartilage and cartilage components as studied by pulsed field gradient NMR. Magn. Reson. Med. 1999, 41, 285–292. [Google Scholar] [CrossRef]
- Rousseau, J.-C.; Chapurlat, R.; Garnero, P. Soluble biological markers in osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2021, 13, 1759720X211040300. [Google Scholar] [CrossRef] [PubMed]
- Roughley, P.J. The structure and function of cartilage proteoglycans. Eur. Cells Mater. 2006, 12, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Hyc, A.; Osiecka-Iwan, A.; Jóźwiak, J.; Moskalewski, S. The morphology and selected biological properties of articular cartilage. Ortop. Traumatol. Rehabil. 2001, 3, 151–162. [Google Scholar] [PubMed]
- Kim, Y.S.; Guilak, F. Engineering Hyaluronic Acid for the Development of New Treatment Strategies for Osteoarthritis. Int. J. Mol. Sci. 2022, 23, 8662. [Google Scholar] [CrossRef] [PubMed]
- Torchia, D.A.; Hasson, M.A.; Hascall, V.C. Investigation of molecular motion of proteoglycans in cartilage by 13C magnetic resonance. J. Biol. Chem. 1977, 252, 3617–3625. [Google Scholar] [CrossRef] [PubMed]
- Belcher, C.; Yaqub, R.; Fawthrop, F.; Bayliss, M.; Doherty, M. Synovial fluid chondroitin and keratan sulphate epitopes, glycosaminoglycans, and hyaluronan in arthritic and normal knees. Ann. Rheum. Dis. 1997, 56, 299–307. [Google Scholar] [CrossRef]
- Gama, C.I.; Hsieh-Wilson, L.C. Chemical approaches to deciphering the glycosaminoglycan code. Curr. Opin. Chem. Biol. 2005, 9, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Guidolin, D. Potential mechanism of action of intra-articular hyaluronan therapy in osteoarthritis: Are the effects molecular weight dependent? Semin. Arthritis Rheum. 2002, 32, 10–37. [Google Scholar] [CrossRef] [PubMed]
- Seidman, A.J.; Limaiem, F. Synovial Fluid Analysis; StatPearls [Internet]: Treasure Island, FL, USA, 2023. [Google Scholar]
- Ben-Trad, L.; Matei, C.I.; Sava, M.M.; Filali, S.; Duclos, M.-E.; Berthier, Y.; Guichardant, M.; Bernoud-Hubac, N.; Maniti, O.; Landoulsi, A.; et al. Synovial Extracellular Vesicles: Structure and Role in Synovial Fluid Tribological Performances. Int. J. Mol. Sci. 2022, 23, 11998. [Google Scholar] [CrossRef] [PubMed]
- Saari, H.; Konttinen, Y.T.; Friman, C.; Sorsa, T. Differential effects of reactive oxygen species on native synovial fluid and purified human umbilical cord hyaluronate. Inflammation 1993, 17, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Arnhold, J.; Sonntag, K.; Arnold, K. NMR studies on human, pathologically changed synovial fluids: Role of hypochlorous acid. Magn. Reson. Med. 1996, 35, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Tieng, A.; Franchin, G. Knee Arthrocentesis in Adults. J. Vis. Exp. 2022, 180, e63135. [Google Scholar] [CrossRef]
- Ma, C.; Wang, J.; Hong, F.; Yang, S. Mitochondrial Dysfunction in Rheumatoid Arthritis. Biomolecules 2022, 12, 1216. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Sun, H.; Bunpetch, V.; Koh, Y.; Wen, Y.; Wu, D.; Ouyang, H. The regulation of cartilage extracellular matrix homeostasis in joint cartilage degeneration and regeneration. Biomaterials 2021, 268, 120555. [Google Scholar] [CrossRef] [PubMed]
- Pulik, Ł.; Łęgosz, P.; Motyl, G. Matrix metalloproteinases in rheumatoid arthritis and osteoarthritis: A state of the art review. Rheumatology 2023, 61, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Fresneda Alarcon, M.; McLaren, Z.; Wright, H.L. Neutrophils in the Pathogenesis of Rheumatoid Arthritis and Systemic Lupus Erythematosus: Same Foe Different M.O. Front. Immunol. 2021, 12, 649693. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Ou, Q.; Zhou, X.; Nie, K.; Liu, J.; Zhang, S. Global research trends and focus on the link between rheumatoid arthritis and neutrophil extracellular traps: A bibliometric analysis from 1985 to 2023. Front. Immunol. 2023, 14, 1205445. [Google Scholar] [CrossRef] [PubMed]
- Mohr, W.; Westerhellweg, H.; Wessinghage, D. Polymorphonuclear granulocytes in rheumatic tissue destruction. III. an electron microscopic study of PMNs at the pannus-cartilage junction in rheumatoid arthritis. Ann. Rheum. Dis. 1981, 40, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, D.; Bai, X. Macrophages regulate the progression of osteoarthritis. Osteoarthr. Cartil. 2020, 28, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Huang, W.; Zhang, W.; Mei, J.; Zhu, C. A Tale of Two Immune Cells in Rheumatoid Arthritis: The Crosstalk between Macrophages and T Cells in the Synovium. Front. Immunol. 2021, 12, 655477. [Google Scholar] [CrossRef]
- Chaney, S.; Vergara, R.; Qiryaqoz, Z.; Suggs, K.; Akkouch, A. The Involvement of Neutrophils in the Pathophysiology and Treatment of Osteoarthritis. Biomedicines 2022, 10, 1604. [Google Scholar] [CrossRef]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Rincón, E.; Rocha-Gregg, B.L.; Collins, S.R. A map of gene expression in neutrophil-like cell lines. BMC Genomics 2018, 19, 573. [Google Scholar] [CrossRef] [PubMed]
- Watson, F.; Robinson, J.J.; Edwards, S.W. Neutrophil function in whole blood and after purification: Changes in receptor expression, oxidase activity and responsiveness to cytokines. Biosci. Rep. 1992, 12, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Jariwala, M.P.; Laxer, R.M. NETosis in Rheumatic Diseases. Curr. Rheumatol. Rep. 2021, 23, 9. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, A.V.; Javadov, S.; Sommer, N. Cellular ROS and Antioxidants: Physiological and Pathological Role. Antioxidants 2024, 13, 602. [Google Scholar] [CrossRef] [PubMed]
- Smallwood, M.J.; Nissim, A.; Knight, A.R.; Whiteman, M.; Haigh, R.; Winyard, P.G. Oxidative stress in autoimmune rheumatic diseases. Free Radic. Biol. Med. 2018, 125, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Jing, W.; Liu, C.; Su, C.; Liu, L.; Chen, P.; Li, X.; Zhang, X.; Yuan, B.; Wang, H.; Du, X. Role of reactive oxygen species and mitochondrial damage in rheumatoid arthritis and targeted drugs. Front. Immunol. 2023, 14, 1107670. [Google Scholar] [CrossRef]
- Loh, J.T.; Lam, K.-P. Neutrophils in the Pathogenesis of Rheumatic Diseases. Rheumatol. Immunol. Res. 2022, 3, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Arve-Butler, S.; Schmidt, T.; Mossberg, A.; Berthold, E.; Gullstrand, B.; Bengtsson, A.A.; Kahn, F.; Kahn, R. Synovial fluid neutrophils in oligoarticular juvenile idiopathic arthritis have an altered phenotype and impaired effector functions. Arthritis Res. Ther. 2021, 23, 109. [Google Scholar] [CrossRef] [PubMed]
- Wright, H.L.; Lyon, M.; Chapman, E.A.; Moots, R.J.; Edwards, S.W. Rheumatoid Arthritis Synovial Fluid Neutrophils Drive Inflammation Through Production of Chemokines, Reactive Oxygen Species, and Neutrophil Extracellular Traps. Front. Immunol. 2020, 11, 584116. [Google Scholar] [CrossRef] [PubMed]
- MacWilliams, P.S.; Friedrichs, K.R. Laboratory evaluation and interpretation of synovial fluid. Vet. Clin. N. Am. Small Anim. Pract. 2003, 33, 153–178. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.-C.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Lanza, V.; Greco, V.; Bocchieri, E.; Sciuto, S.; Inturri, R.; Messina, L.; Vaccaro, S.; Bellia, F.; Rizzarelli, E. Synergistic Effect of L-Carnosine and Hyaluronic Acid in Their Covalent Conjugates on the Antioxidant Abilities and the Mutual Defense against Enzymatic Degradation. Antioxidants 2022, 11, 664. [Google Scholar] [CrossRef] [PubMed]
- Babior, B.M.; Kipnes, R.S.; Curnutte, J.T. Biological defense mechanisms. The production by leukocytes of superoxide, a potential bactericidal agent. J. Clin. Investig. 1973, 52, 741–744. [Google Scholar] [CrossRef]
- Mohan, I.K.; Baba, K.S.S.S.; Iyyapu, R.; Thirumalasetty, S.; Satish, O.S. Advances in congestive heart failure biomarkers. Adv. Clin. Chem. 2023, 112, 205–248. [Google Scholar] [CrossRef] [PubMed]
- Rayner, B.S.; Zhang, Y.; Brown, B.E.; Reyes, L.; Cogger, V.C.; Hawkins, C.L. Role of hypochlorous acid (HOCl) and other inflammatory mediators in the induction of macrophage extracellular trap formation. Free Radic. Biol. Med. 2018, 129, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Kettle, A.J.; Winterbourn, C.C. Myeloperoxidase: A key regulator of neutrophil oxidant production. Redox Rep. 1997, 3, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, J.; Flemmig, J. Human myeloperoxidase in innate and acquired immunity. Arch. Biochem. Biophys. 2010, 500, 92–106. [Google Scholar] [CrossRef]
- Van Dalen, C.J.; Whitehouse, M.W.; Winterbourn, C.C.; Kettle, A.J. Thiocyanate and chloride as competing substrates for myeloperoxidase. Biochem. J. 1997, 327, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an Active Disease Biomarker: Recent Biochemical and Pathological Perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Bedouhène, S.; Dang, P.M.-C.; Hurtado-Nedelec, M.; El-Benna, J. Neutrophil Degranulation of Azurophil and Specific Granules. Methods Mol. Biol. 2020, 2087, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, J. The Dual Role of Myeloperoxidase in Immune Response. Int. J. Mol. Sci. 2020, 21, 8057. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Krumbholz, M.; Schönermarck, U.; Back, W.; Gross, W.L.; Werb, Z.; Gröne, H.-J.; Brinkmann, V.; Jenne, D.E. Netting neutrophils in autoimmune small-vessel vasculitis. Nat. Med. 2009, 15, 623–625. [Google Scholar] [CrossRef] [PubMed]
- Baldus, S.; Heeschen, C.; Meinertz, T.; Zeiher, A.M.; Eiserich, J.P.; Münzel, T.; Simoons, M.L.; Hamm, C.W. Myeloperoxidase serum levels predict risk in patients with acute coronary syndromes. Circulation 2003, 108, 1440–1445. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, J.; Malle, E. Halogenation Activity of Mammalian Heme Peroxidases. Antioxidants 2022, 11, 890. [Google Scholar] [CrossRef] [PubMed]
- Korotaeva, A.; Samoilova, E.; Pavlunina, T.; Panasenko, O.M. Halogenated phospholipids regulate secretory phospholipase A2 group IIA activity. Chem. Phys. Lipids 2013, 167–168, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Katrantzis, M.; Baker, M.S.; Handley, C.J.; Lowther, D.A. The oxidant hypochlorite (OCl−), a product of the myeloperoxidase system, degrades articular cartilage proteoglycan aggregate. Free Radic. Biol. Med. 1991, 10, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Yin, C.; Huo, F. Small-Molecule Fluorescent Probes for Detecting Several Abnormally Expressed Substances in Tumors. Micromachines 2022, 13, 1328. [Google Scholar] [CrossRef] [PubMed]
- Haegens, A.; Vernooy, J.H.J.; Heeringa, P.; Mossman, B.T.; Wouters, E.F.M. Myeloperoxidase modulates lung epithelial responses to pro-inflammatory agents. Eur. Respir. J. 2008, 31, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Morozov, V.I.; Pryatkin, S.A.; Kalinski, M.I.; Rogozkin, V.A. Effect of exercise to exhaustion on myeloperoxidase and lysozyme release from blood neutrophils. Eur. J. Appl. Physiol. 2003, 89, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Hadler, N.M.; Spitznagel, J.K.; Quinet, R.J. Lysosomal Enzymes in Inflammatory Synovial Effusions. J. Immunol. 1979, 123, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, B.; Horwitz, M.S.; Jenne, D.E.; Gauthier, F. Neutrophil elastase, proteinase 3, and cathepsin G as therapeutic targets in human diseases. Pharmacol. Rev. 2010, 62, 726–759. [Google Scholar] [CrossRef] [PubMed]
- Gotis-Graham, I.; Hogg, P.J.; McNeil, H.P. Significant correlation between thrombospondin 1 and serine proteinase expression in rheumatoid synovium. Arthritis Rheum. 1997, 40, 1780–1787. [Google Scholar] [CrossRef] [PubMed]
- Momohara, S.; Kashiwazaki, S.; Inoue, K.; Saito, S.; Nakagawa, T. Elastase from polymorphonuclear leukocyte in articular cartilage and synovial fluids of patients with rheumatoid arthritis. Clin. Rheumatol. 1997, 16, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Parker, H.; Albrett, A.M.; Kettle, A.J.; Winterbourn, C.C. Myeloperoxidase associated with neutrophil extracellular traps is active and mediates bacterial killing in the presence of hydrogen peroxide. J. Leukoc. Biol. 2012, 91, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.C. The Acid Ionization Constant of HOCl from 5 to 35°. J. Phys. Chem. 1966, 70, 3798–3805. [Google Scholar] [CrossRef]
- Wan, Z.; Zhu, C.; Francisco, J.S. Molecular Insights into the Spontaneous Generation of Cl2O in the Reaction of ClONO2 and HOCl at the Air-Water Interface. J. Am. Chem. Soc. 2023, 145, 17478–17484. [Google Scholar] [CrossRef] [PubMed]
- Sivey, J.D.; McCullough, C.E.; Roberts, A.L. Chlorine monoxide (Cl2O) and molecular chlorine (Cl2) as active chlorinating agents in reaction of dimethenamid with aqueous free chlorine. Environ. Sci. Technol. 2010, 44, 3357–3362. [Google Scholar] [CrossRef] [PubMed]
- Eiserich, J.P.; Hristova, M.; Cross, C.E.; Jones, A.D.; Freeman, B.A.; Halliwell, B.; van der Vliet, A. Formation of nitric oxide-derived inflammatory oxidants by myeloperoxidase in neutrophils. Nature 1998, 391, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Chuang, C.Y.; Hawkins, C.L.; Hägglund, P.; Davies, M.J. Identification and quantification of protein nitration sites in human coronary artery smooth muscle cells in the absence and presence of peroxynitrous acid/peroxynitrite. Redox Biol. 2023, 64, 102799. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. Comparative reactivities of various biological compounds with myeloperoxidase-hydrogen peroxide-chloride, and similarity of the oxidant to hypochlorite. Biochim. Biophys. Acta 1985, 840, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Pattison, D.I.; Davies, M.J. Absolute rate constants for the reaction of hypochlorous acid with protein side chains and peptide bonds. Chem. Res. Toxicol. 2001, 14, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, J.; Hammerschmidt, S.; Arnold, K. Role of functional groups of human plasma and luminol in scavenging of NaOCl and neutrophil-derived hypochlorous acid. Biochim. Biophys. Acta 1991, 1097, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Prütz, W.A. Hypochlorous acid interactions with thiols, nucleotides, DNA, and other biological substrates. Arch. Biochem. Biophys. 1996, 332, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Armesto, X.L.; Canle, M.L.; Santaballa, J.A. α-amino acids chlorination in aqueous media. Tetrahedron 1993, 49, 275–284. [Google Scholar] [CrossRef]
- Pattison, D.I.; Hawkins, C.L.; Davies, M.J. Hypochlorous acid-mediated oxidation of lipid components and antioxidants present in low-density lipoproteins: Absolute rate constants, product analysis, and computational modeling. Chem. Res. Toxicol. 2003, 16, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Panasenko, O.M.; Torkhovskaya, T.I.; Gorudko, I.V.; Sokolov, A.V. The Role of Halogenative Stress in Atherogenic Modification of Low-Density Lipoproteins. Biochemistry 2020, 85, S34–S55. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef] [PubMed]
- Bielski, B.H.; Shiue, G.G. Reaction Rates of Superoxide Radicals with the Essential Amino Acids; Excerpta Medica: Amsterdam, The Netherlands, 1978; pp. 43–56. [Google Scholar] [CrossRef]
- Burkhardt, H.; Schwingel, M.; Menninger, H.; Macartney, H.W.; Tschesche, H. Oxygen radicals as effectors of cartilage destruction. Direct degradative effect on matrix components and indirect action via activation of latent collagenase from polymorphonuclear leukocytes. Arthritis Rheum. 1986, 29, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Abe, C.; Miyazawa, T.; Miyazawa, T. Current Use of Fenton Reaction in Drugs and Food. Molecules 2022, 27, 5451. [Google Scholar] [CrossRef]
- Fujii, J.; Homma, T.; Osaki, T. Superoxide Radicals in the Execution of Cell Death. Antioxidants 2022, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Regan, E.; Flannelly, J.; Bowler, R.; Tran, K.; Nicks, M.; Carbone, B.D.; Glueck, D.; Heijnen, H.; Mason, R.; Crapo, J. Extracellular superoxide dismutase and oxidant damage in osteoarthritis. Arthritis Rheum. 2005, 52, 3479–3491. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Arnhold, J.; Arnold, K. NMR studies of the action of hypochlorous acid on native pig articular cartilage. Eur. J. Biochem. 1995, 233, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Arnhold, J.; Zachäus, A.; Arnold, K. Reaction of hypochlorous acid with bovine nasal cartilage comparison to pig articular cartilage. Z. Naturforsch. C J. Biosci. 1997, 52, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Hadjigogos, K. The role of free radicals in the pathogenesis of rheumatoid arthritis. Panminerva Med. 2003, 45, 7–13. [Google Scholar]
- Davies, J.M.; Horwitz, D.A.; Davies, K.J. Inhibition of collagenase activity by N-chlorotaurine, a product of activated neutrophils. Arthritis Rheum. 1994, 37, 424–427. [Google Scholar] [CrossRef]
- Stamp, L.K.; Khalilova, I.; Tarr, J.M.; Senthilmohan, R.; Turner, R.; Haigh, R.C.; Winyard, P.G.; Kettle, A.J. Myeloperoxidase and oxidative stress in rheumatoid arthritis. Rheumatology 2012, 51, 1796–1803. [Google Scholar] [CrossRef] [PubMed]
- Odobasic, D.; Yang, Y.; Muljadi, R.C.M.; O’Sullivan, K.M.; Kao, W.; Smith, M.; Morand, E.F.; Holdsworth, S.R. Endogenous myeloperoxidase is a mediator of joint inflammation and damage in experimental arthritis. Arthritis Rheumatol. 2014, 66, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Garvican, E.R.; Vaughan-Thomas, A.; Innes, J.F.; Clegg, P.D. Biomarkers of cartilage turnover. Part 1: Markers of collagen degradation and synthesis. Vet. J. 2010, 185, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Westman, E.; Lundberg, K.; Erlandsson Harris, H. Arthritogenicity of collagen type II is increased by chlorination. Clin. Exp. Immunol. 2006, 145, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.M.; Horwitz, D.A.; Davies, K.J. Potential roles of hypochlorous acid and N-chloroamines in collagen breakdown by phagocytic cells in synovitis. Free Radic. Biol. Med. 1993, 15, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Daumer, K.M.; Khan, A.U.; Steinbeck, M.J. Chlorination of pyridinium compounds. Possible role of hypochlorite, N-chloramines, and chlorine in the oxidation of pyridinoline cross-links of articular cartilage collagen type II during acute inflammation. J. Biol. Chem. 2000, 275, 34681–34692. [Google Scholar] [CrossRef] [PubMed]
- Olszowski, S.; Mak, P.; Olszowska, E.; Marcinkiewicz, J. Collagen type II modification by hypochlorite. Acta Biochim. Pol. 2003, 50, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Bochi, G.V.; Torbitz, V.D.; de Campos, L.P.; Sangoi, M.B.; Fernandes, N.F.; Gomes, P.; Moretto, M.B.; Barbisan, F.; Da Cruz, I.B.M.; Moresco, R.N. In Vitro Oxidation of Collagen Promotes the Formation of Advanced Oxidation Protein Products and the Activation of Human Neutrophils. Inflammation 2016, 39, 916–927. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Gauld, J.W. Sulfilimine bond formation in collagen IV. Chem. Commun. 2024, 60, 646–657. [Google Scholar] [CrossRef] [PubMed]
- Ronsein, G.E.; Winterbourn, C.C.; Di Mascio, P.; Kettle, A.J. Cross-linking methionine and amine residues with reactive halogen species. Free Radic. Biol. Med. 2014, 70, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Winyard, P.G.; Ryan, B.; Eggleton, P.; Nissim, A.; Taylor, E.; Lo Faro, M.L.; Burkholz, T.; Szabó-Taylor, K.E.; Fox, B.; Viner, N.; et al. Measurement and meaning of markers of reactive species of oxygen, nitrogen and sulfur in healthy human subjects and patients with inflammatory joint disease. Biochem. Soc. Trans. 2011, 39, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, B.; Schiller, J. Glycosaminoglycan degradation by selected reactive oxygen species. Antioxid. Redox Signal. 2014, 21, 1044–1062. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.S.; Green, S.P.; Lowther, D.A. Changes in the viscosity of hyaluronic acid after exposure to a myeloperoxidase-derived oxidant. Arthritis Rheum. 1989, 32, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Martin-Alarcon, L.; Schmidt, T.A. Rheological effects of macromolecular interactions in synovial fluid. Biorheology 2016, 53, 49–67. [Google Scholar] [CrossRef]
- Cowman, M.K.; Matsuoka, S. Experimental approaches to hyaluronan structure. Carbohydr. Res. 2005, 340, 791–809. [Google Scholar] [CrossRef] [PubMed]
- Sprott, H.; Fleck, C. Hyaluronic Acid in Rheumatology. Pharmaceutics 2023, 15, 2247. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.H.; Xue, J.F.; Zheng, Z.Y.; Shuhaidi, M.; Thu, H.E.; Hussain, Z. Hyaluronic acid, an efficient biomacromolecule for treatment of inflammatory skin and joint diseases: A review of recent developments and critical appraisal of preclinical and clinical investigations. Int. J. Biol. Macromol. 2018, 116, 572–584. [Google Scholar] [CrossRef]
- Conrozier, T.; Mathieu, P.; Vignon, E.; Piperno, M.; Rinaudo, M. Differences in the osteoarthritic synovial fluid composition and rheology between patients with or without flare: A pilot study. Clin. Exp. Rheumatol. 2012, 30, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Arnhold, J.; Gründer, W.; Arnold, K. The action of hypochlorous acid on polymeric components of cartilage. Biol. Chem. Hoppe Seyler 1994, 375, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Degradation of hyaluronic acid, poly- and monosaccharides, and model compounds by hypochlorite: Evidence for radical intermediates and fragmentation. Free Radic. Biol. Med. 1998, 24, 1396–1410. [Google Scholar] [CrossRef] [PubMed]
- Grootveld, M.; Henderson, E.B.; Farrell, A.; Blake, D.R.; Parkes, H.G.; Haycock, P. Oxidative damage to hyaluronate and glucose in synovial fluid during exercise of the inflamed rheumatoid joint. Detection of abnormal low-molecular-mass metabolites by proton-n.m.r. spectroscopy. Biochem. J. 1991, 273 Pt 2, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.; Arnhold, J.; Schwinn, J.; Sprinz, H.; Brede, O.; Arnold, K. Reactivity of cartilage and selected carbohydrates with hydroxyl radicals: An NMR study to detect degradation products. Free Radic. Res. 1998, 28, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Rees, M.D.; Hawkins, C.L.; Davies, M.J. Hypochlorite and superoxide radicals can act synergistically to induce fragmentation of hyaluronan and chondroitin sulphates. Biochem. J. 2004, 381, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.; Prabutzki, P.; Nimptsch, A.; Schiller, J. Mass spectrometric investigations of the action of hypochlorous acid on monomeric and oligomeric components of glycosaminoglycans. Biochem. Biophys. Rep. 2023, 34, 101448. [Google Scholar] [CrossRef]
- Kishimoto, N.; Kitamura, T.; Kato, M.; Otsu, H. Reusability of iron sludge as an iron source for the electrochemical Fenton-type process using Fe2+/HOCl system. Water Res. 2013, 47, 1919–1927. [Google Scholar] [CrossRef] [PubMed]
- Akeel, A.; Sibanda, S.; Martin, S.W.; Paterson, A.W.J.; Parsons, B.J. Chlorination and oxidation of heparin and hyaluronan by hypochlorous acid and hypochlorite anions: Effect of sulfate groups on reaction pathways and kinetics. Free Radic. Biol. Med. 2013, 56, 72–88. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Ginsburg, I. A Novel Hypothetical Approach to Explain the Mechanisms of Pathogenicity of Rheumatic Arthritis. Mediterr. J. Rheumatol. 2021, 32, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Corsaro, M.M.; Pietraforte, D.; Di Lorenzo, A.S.; Minetti, M.; Marino, G. Reaction of peroxynitrite with hyaluronan and related saccharides. Free Radic. Res. 2004, 38, 343–353. [Google Scholar] [CrossRef]
- Lemmnitzer, K.; Köhling, S.; Freyse, J.; Rademann, J.; Schiller, J. Characterization of defined sulfated heparin-like oligosaccharides by electrospray ionization ion trap mass spectrometry. J. Mass Spectrom. 2021, 56, e4692. [Google Scholar] [CrossRef] [PubMed]
- Grabarics, M.; Lettow, M.; Kirschbaum, C.; Greis, K.; Manz, C.; Pagel, K. Mass Spectrometry-Based Techniques to Elucidate the Sugar Code. Chem. Rev. 2022, 122, 7840–7908. [Google Scholar] [CrossRef]
- Buffa, R.; Hermannová, M.; Sojka, M.; Svozil, V.; Šulc, P.; Halamková, P.; Pospíšilová, M.; Krejčí, H.; Velebný, V. Hyaluronic acid chloramide-Synthesis, chemical structure, stability and analysis of antimicrobials. Carbohydr. Polym. 2020, 250, 116928. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.; Szabó, M.; Fábián, I. The chlorination of glycine and α-alanine at excess HOCl: Kinetics and mechanism. J. Hazard. Mater. 2023, 447, 130794. [Google Scholar] [CrossRef] [PubMed]
- Steinbeck, M.J.; Nesti, L.J.; Sharkey, P.F.; Parvizi, J. Myeloperoxidase and chlorinated peptides in osteoarthritis: Potential biomarkers of the disease. J. Orthop. Res. 2007, 25, 1128–1135. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R.; Chokesuwattanaskul, S.; Phelan, M.M.; Welting, T.J.M.; Lian, L.-Y.; Peffers, M.J.; Wright, H.L. 1H NMR Metabolomics Identifies Underlying Inflammatory Pathology in Osteoarthritis and Rheumatoid Arthritis Synovial Joints. J. Proteome Res. 2018, 17, 3780–3790. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.L.; Davies, M.J. Role of myeloperoxidase and oxidant formation in the extracellular environment in inflammation-induced tissue damage. Free Radic. Biol. Med. 2021, 172, 633–651. [Google Scholar] [CrossRef]
- Flemmig, J.; Spalteholz, H.; Schubert, K.; Meier, S.; Arnhold, J. Modification of phosphatidylserine by hypochlorous acid. Chem. Phys. Lipids 2009, 161, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; La Pérez de Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Hypochlorous Acid Chemistry in Mammalian Cells-Influence on Infection and Role in Various Pathologies. Int. J. Mol. Sci. 2022, 23, 10735. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Wagner, J.R. DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb. Perspect. Biol. 2013, 5, a012559. [Google Scholar] [CrossRef] [PubMed]
- Karaman, A.; Binici, D.N.; Melikoğlu, M.A. Comet assay and analysis of micronucleus formation in patients with rheumatoid arthritis. Mutat. Res. 2011, 721, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Galita, G.; Sarnik, J.; Brzezinska, O.; Budlewski, T.; Dragan, G.; Poplawska, M.; Majsterek, I.; Poplawski, T.; Makowska, J.S. Polymorphisms in DNA Repair Genes and Association with Rheumatoid Arthritis in a Pilot Study on a Central European Population. Int. J. Mol. Sci. 2023, 24, 3804. [Google Scholar] [CrossRef] [PubMed]
- Galita, G.; Sarnik, J.; Brzezinska, O.; Budlewski, T.; Poplawska, M.; Sakowski, S.; Dudek, G.; Majsterek, I.; Makowska, J.; Poplawski, T. The Association between Inefficient Repair of DNA Double-Strand Breaks and Common Polymorphisms of the HRR and NHEJ Repair Genes in Patients with Rheumatoid Arthritis. Int. J. Mol. Sci. 2024, 25, 2619. [Google Scholar] [CrossRef] [PubMed]
- De Sandes Kimura, O.; Mozella, A.; Cobra, H.; Maciel Saraiva, A.C.; Carvalho de Almendra Freitas, E.H.; Cury Fernandes, M.B.; Matheus Guimarães, J.A.; Defino, H.; Leal, A.C. Neutrophil Extracellular Trap-related Biomarkers Are Increased in the Synovial Fluid of Patients with Periprosthetic Joint Infections. Clin. Orthop. Relat. Res. 2024, 482, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, R.A. Lipid content of human costal and articular cartilage. Ann. Rheum. Dis. 1967, 26, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Schröter, J.; Schiller, J. Chlorinated Phospholipids and Fatty Acids: (Patho)physiological Relevance, Potential Toxicity, and Analysis of Lipid Chlorohydrins. Oxid. Med. Cell. Longev. 2016, 2016, 8386362. [Google Scholar] [CrossRef] [PubMed]
- Harayama, T.; Antonny, B. Beyond Fluidity: The Role of Lipid Unsaturation in Membrane Function. Cold Spring Harb. Perspect. Biol. 2023, 15, a041409. [Google Scholar] [CrossRef] [PubMed]
- Richter, G.; Schober, C.; Süss, R.; Fuchs, B.; Birkemeyer, C.; Schiller, J. Comparison of the positive and negative ion electrospray ionization and matrix-assisted laser desorption ionization-time-of-flight mass spectra of the reaction products of phosphatidylethanolamines and hypochlorous acid. Anal. Biochem. 2008, 376, 157–159. [Google Scholar] [CrossRef] [PubMed]
- Arnhold, J.; Osipov, A.N.; Spalteholz, H.; Panasenko, O.M.; Schiller, J. Effects of hypochlorous acid on unsaturated phosphatidylcholines. Free Radic. Biol. Med. 2001, 31, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; van den Berg, J.J.; Roitman, E.; Kuypers, F.A. Chlorohydrin formation from unsaturated fatty acids reacted with hypochlorous acid. Arch. Biochem. Biophys. 1992, 296, 547–555. [Google Scholar] [CrossRef] [PubMed]
- De-Madaria, E.; Molero, X.; Bonjoch, L.; Casas, J.; Cárdenas-Jaén, K.; Montenegro, A.; Closa, D. Oleic acid chlorohydrin, a new early biomarker for the prediction of acute pancreatitis severity in humans. Ann. Intensive Care 2018, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Schröter, J.; Griesinger, H.; Reuß, E.; Schulz, M.; Riemer, T.; Süß, R.; Schiller, J.; Fuchs, B. Unexpected products of the hypochlorous acid-induced oxidation of oleic acid: A study using high performance thin-layer chromatography-electrospray ionization mass spectrometry. J. Chromatogr. A 2016, 1439, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Leopold, J.; Prabutzki, P.; Engel, K.M.; Schiller, J. From Oxidized Fatty Acids to Dimeric Species: In Vivo Relevance, Generation and Methods of Analysis. Molecules 2023, 28, 7850. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Vissers, M.C.; Domigan, N.M.; Winterbourn, C.C. Modification of red cell membrane lipids by hypochlorous acid and haemolysis by preformed lipid chlorohydrins. Redox Rep. 1997, 3, 263–271. [Google Scholar] [CrossRef] [PubMed]
- L’Huillier, N.; Pratten, M.K.; Clothier, R.H. The relative embryotoxicity of 1,3-dichloro-2-propanol on primary chick embryonic cells. Toxicol. Vitr. 2002, 16, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.J.; Blake, D.R.; Palmer, R.M.; Moncada, S. Increased concentrations of nitrite in synovial fluid and serum samples suggest increased nitric oxide synthesis in rheumatic diseases. Ann. Rheum. Dis. 1992, 51, 1219–1222. [Google Scholar] [CrossRef] [PubMed]
- Fitzl, M.; Süss, R.; Arnold, K.; Schiller, J. Nitrite reduces the effects of HOCl on unsaturated phosphatidylcholines—A MALDI-TOF MS study. Chem. Phys. Lipids 2006, 140, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Rose, P.; Siau, J.L.; Halliwell, B. Nitrite-mediated protection against hypochlorous acid-induced chondrocyte toxicity: A novel cytoprotective role of nitric oxide in the inflamed joint? Arthritis Rheum. 2003, 48, 3140–3150. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.S.; Mawatari, S.; Fujino, T. Biological Functions of Plasmalogens. Adv. Exp. Med. Biol. 2020, 1299, 171–193. [Google Scholar] [CrossRef] [PubMed]
- Kosinska, M.K.; Liebisch, G.; Lochnit, G.; Wilhelm, J.; Klein, H.; Kaesser, U.; Lasczkowski, G.; Rickert, M.; Schmitz, G.; Steinmeyer, J. A lipidomic study of phospholipid classes and species in human synovial fluid. Arthritis Rheum. 2013, 65, 2323–2333. [Google Scholar] [CrossRef] [PubMed]
- Shakya, S.; Herr, R.A.; Carlson, H.L.; Zoeller, R.A.; Albert, C.J.; Ford, D.A. Endothelial Cell Protein Targeting by Myeloperoxidase-Derived 2-Chlorofatty Aldehyde. Antioxidants 2022, 11, 940. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, H.B.; Melo, T.; Paiva, A.; Domingues, M.d.R. Insights in the Role of Lipids, Oxidative Stress and Inflammation in Rheumatoid Arthritis Unveiled by New Trends in Lipidomic Investigations. Antioxidants 2021, 10, 45. [Google Scholar] [CrossRef] [PubMed]
- Packer, L.; Kraemer, K.; Rimbach, G. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition 2001, 17, 888–895. [Google Scholar] [CrossRef]
- Packer, L.; Witt, E.H.; Tritschler, H.J. alpha-Lipoic acid as a biological antioxidant. Free Radic. Biol. Med. 1995, 19, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, H.H.; Hozayen, W.G.; Khaliefa, A.K.; El-Kenawy, A.E.; Ali, T.M.; Ahmed, O.M. Diosmin and Trolox Have Anti-Arthritic, Anti-Inflammatory and Antioxidant Potencies in Complete Freund’s Adjuvant-Induced Arthritic Male Wistar Rats: Roles of NF-κB, iNOS, Nrf2 and MMPs. Antioxidants 2022, 11, 1721. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Ximenes, V.F.; Ximenes, T.P.; Morgon, N.H.; de Souza, A.R. Taurine Chloramine and Hydrogen Peroxide as a Potential Source of Singlet Oxygen for Topical Application. Photochem. Photobiol. 2021, 97, 963–970. [Google Scholar] [CrossRef]
- Verdrengh, M.; Tarkowski, A. Inhibition of septic arthritis by local administration of taurine chloramine, a product of activated neutrophils. J. Rheumatol. 2005, 32, 1513–1517. [Google Scholar] [PubMed]
- Gottardi, W.; Debabov, D.; Nagl, M. N-chloramines, a promising class of well-tolerated topical anti-infectives. Antimicrob. Agents Chemother. 2013, 57, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Takeuchi, R. Reactions of Methotrexate with Hypobromous Acid and Hypochlorous Acid. Chem. Pharm. Bull. 2019, 67, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Koelsch, M.; Mallak, R.; Graham, G.G.; Kajer, T.; Milligan, M.K.; Nguyen, L.Q.; Newsham, D.W.; Keh, J.S.; Kettle, A.J.; Scott, K.F.; et al. Acetaminophen (paracetamol) inhibits myeloperoxidase-catalyzed oxidant production and biological damage at therapeutically achievable concentrations. Biochem. Pharmacol. 2010, 79, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Chaikijurajai, T.; Tang, W.H.W. Myeloperoxidase: A potential therapeutic target for coronary artery disease. Expert Opin. Ther. Targets 2020, 24, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, K.A.; Jing, X.; Teng, M.; Wells, C.; Jia, S.; Afolayan, A.J.; Jarzembowski, J.; Day, B.W.; Naylor, S.; Hessner, M.J.; et al. Role of endoplasmic reticulum stress in impaired neonatal lung growth and bronchopulmonary dysplasia. PLoS ONE 2022, 17, e0269564. [Google Scholar] [CrossRef] [PubMed]
- Tidén, A.-K.; Sjögren, T.; Svensson, M.; Bernlind, A.; Senthilmohan, R.; Auchère, F.; Norman, H.; Markgren, P.-O.; Gustavsson, S.; Schmidt, S.; et al. 2-thioxanthines are mechanism-based inactivators of myeloperoxidase that block oxidative stress during inflammation. J. Biol. Chem. 2011, 286, 37578–37589. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, H.; Chen, X.; Guo, C. The Roles of Neutrophil-Derived Myeloperoxidase (MPO) in Diseases: The New Progress. Antioxidants 2024, 13, 132. [Google Scholar] [CrossRef]
- Pahwa, R.; Modi, P.; Jialal, I. Myeloperoxidase Deficiency; StatPearls [Internet]: Treasure Island, FL, USA, 2024. [Google Scholar]
- Zlatar, L.; Mahajan, A.; Muñoz-Becerra, M.; Weidner, D.; Bila, G.; Bilyy, R.; Titze, J.; Hoffmann, M.H.; Schett, G.; Herrmann, M.; et al. Suppression of neutrophils by sodium exacerbates oxidative stress and arthritis. Front. Immunol. 2023, 14, 1174537. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Tang, C.; Wang, M.; Zhao, H.; Zhu, Y. Ferroptosis as an emerging target in rheumatoid arthritis. Front. Immunol. 2023, 14, 1260839. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhang, B.; Xi, C.; Qin, Y.; Lin, X.; Wang, B.; Kong, P.; Yan, J. Ferroptosis Plays a Role in Human Chondrocyte of Osteoarthritis Induced by IL-1β In Vitro. Cartilage 2023, 14, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Frade-Sosa, B.; Sanmartí, R. Neutrophils, neutrophil extracellular traps, and rheumatoid arthritis: An updated review for clinicians. Reumatol. Clin. (Engl. Ed.) 2023, 19, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Jung, N.; Bueb, J.-L.; Tolle, F.; Bréchard, S. Regulation of neutrophil pro-inflammatory functions sheds new light on the pathogenesis of rheumatoid arthritis. Biochem. Pharmacol. 2019, 165, 170–180. [Google Scholar] [CrossRef]
- Apel, F.; Zychlinsky, A.; Kenny, E.F. The role of neutrophil extracellular traps in rheumatic diseases. Nat. Rev. Rheumatol. 2018, 14, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Du, C.; Zhang, Y.; Zhu, L. Composition and Function of Neutrophil Extracellular Traps. Biomolecules 2024, 14, 416. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Ren, H.; Jiang, J.; Shao, C.; Shi, Y.; Li, P. Dying to Defend: Neutrophil Death Pathways and their Implications in Immunity. Adv. Sci. 2024, 11, e2306457. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Jiang, P.; Guo, S.; Schrodi, S.J.; He, D. Apoptosis, Autophagy, NETosis, Necroptosis, and Pyroptosis Mediated Programmed Cell Death as Targets for Innovative Therapy in Rheumatoid Arthritis. Front. Immunol. 2021, 12, 809806. [Google Scholar] [CrossRef] [PubMed]
- Reshetnyak, T.; Nurbaeva, K.; Ptashnik, I.; Kudriaeva, A.; Belogurov, A.; Lila, A.; Nasonov, E. Markers of NETosis in Patients with Systemic Lupus Erythematosus and Antiphospholipid Syndrome. Int. J. Mol. Sci. 2023, 24, 9210. [Google Scholar] [CrossRef] [PubMed]
- Singhal, A.; Kumar, S. Neutrophil and remnant clearance in immunity and inflammation. Immunology 2022, 165, 22–43. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, L.A.E.; Barlous, K.; Hawkins, C.L. Antioxidant Strategies to Modulate NETosis and the Release of Neutrophil Extracellular Traps during Chronic Inflammation. Antioxidants 2023, 12, 478. [Google Scholar] [CrossRef] [PubMed]
- Kosonen, J.P.; Eskelinen, A.S.A.; Orozco, G.A.; Nieminen, P.; Anderson, D.D.; Grodzinsky, A.J.; Korhonen, R.K.; Tanska, P. Injury-related cell death and proteoglycan loss in articular cartilage: Numerical model combining necrosis, reactive oxygen species, and inflammatory cytokines. PLoS Comput. Biol. 2023, 19, e1010337. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Jian, Z.; Guo, J.; Ning, X. Increased levels of serum myeloperoxidase in patients with active rheumatoid arthritis. Life Sci. 2014, 117, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Talib, J.; Maghzal, G.J.; Cheng, D.; Stocker, R. Detailed protocol to assess in vivo and ex vivo myeloperoxidase activity in mouse models of vascular inflammation and disease using hydroethidine. Free Radic. Biol. Med. 2016, 97, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, J.H.; Rai, V.; Dilisio, M.F.; Agrawal, D.K. Damage-associated molecular patterns in the pathogenesis of osteoarthritis: Potentially novel therapeutic targets. Mol. Cell. Biochem. 2017, 434, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cao, J.; Caterson, B.; Hughes, C.E. Proteoglycan metabolism, cell death and Kashin-Beck disease. Glycoconj. J. 2012, 29, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Lepetsos, P.; Papavassiliou, A.G. ROS/oxidative stress signaling in osteoarthritis. Biochim. Biophys. Acta 2016, 1862, 576–591. [Google Scholar] [CrossRef] [PubMed]
- Lepetsos, P.; Papavassiliou, K.A.; Papavassiliou, A.G. Redox and NF-κB signaling in osteoarthritis. Free Radic. Biol. Med. 2019, 132, 90–100. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Survey of the most abundant glycosaminoglycans (GAGs) of healthy hyaline articular cartilage. As schematically shown, chrondroitin sulfate (CS, green colored) chains are more abundant than keratan sulfate (KS, purple colored) chains. Hyaluronic acid (HA, red colored), also named hyaluronan, functions as the backbone in the so-called “bottle-brush” structure. Structural changes in GAGs may lead to the degradation of the cartilage and thus to function loss of the cartilage. Characteristic patterns of healthy, rheumatoid arthritis (RA) as well as osteoarthritis (OA) cartilage are illustrated using stock photos from the illustration platform Colorbox (reproduced with permission using the campus license from Leipzig University).
Figure 1.
Survey of the most abundant glycosaminoglycans (GAGs) of healthy hyaline articular cartilage. As schematically shown, chrondroitin sulfate (CS, green colored) chains are more abundant than keratan sulfate (KS, purple colored) chains. Hyaluronic acid (HA, red colored), also named hyaluronan, functions as the backbone in the so-called “bottle-brush” structure. Structural changes in GAGs may lead to the degradation of the cartilage and thus to function loss of the cartilage. Characteristic patterns of healthy, rheumatoid arthritis (RA) as well as osteoarthritis (OA) cartilage are illustrated using stock photos from the illustration platform Colorbox (reproduced with permission using the campus license from Leipzig University).
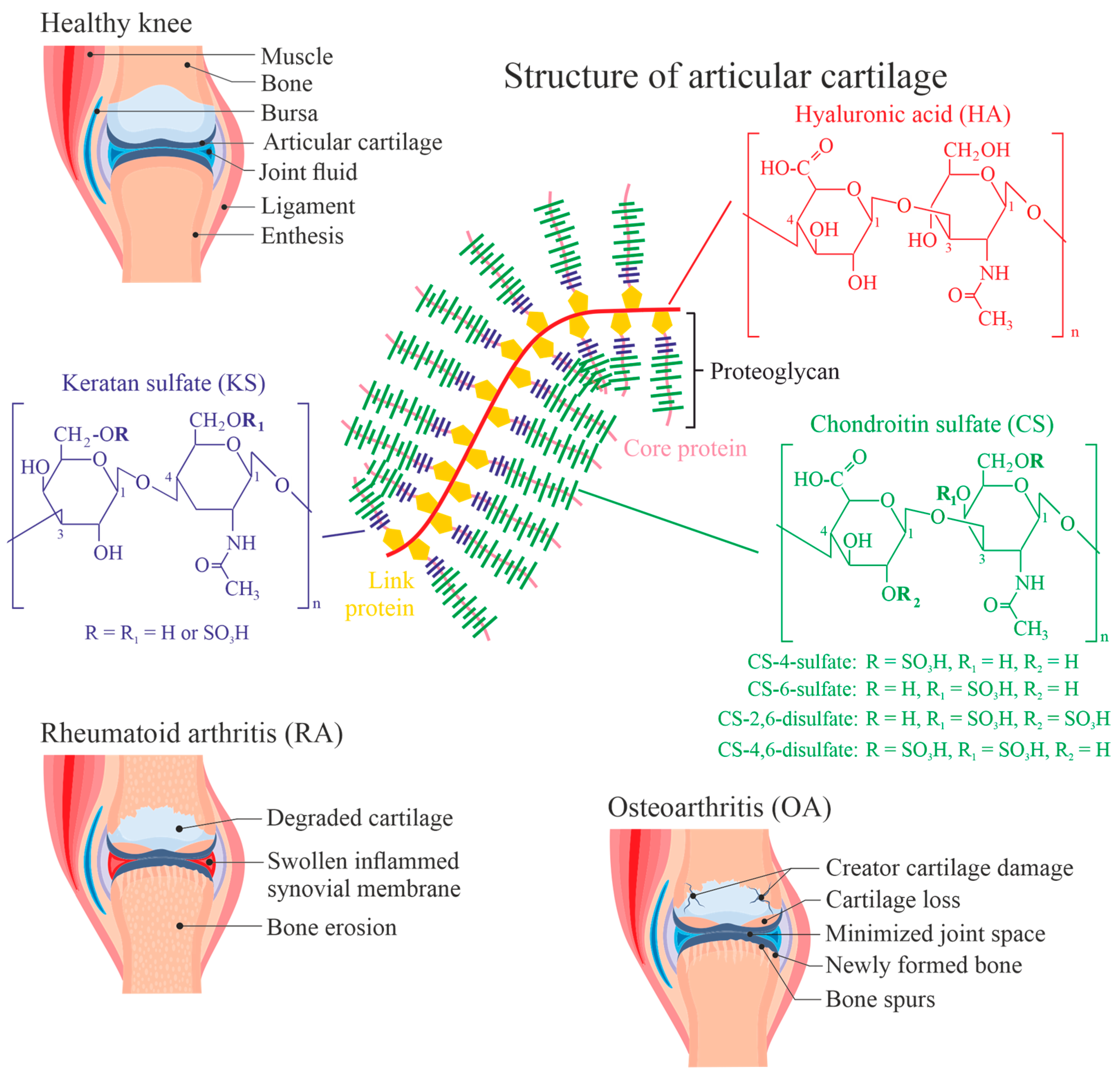
Figure 2.
Pathways leading to and from different relevant reactive species (marked in blue). The HOCl generation is catalyzed by the enzyme myeloperoxidase (MPO) via the chlorination cycle (shown in detail, blue colored box). MPO does not exclusively metabolize Cl−, but Br− and SCN− as well.
Figure 2.
Pathways leading to and from different relevant reactive species (marked in blue). The HOCl generation is catalyzed by the enzyme myeloperoxidase (MPO) via the chlorination cycle (shown in detail, blue colored box). MPO does not exclusively metabolize Cl−, but Br− and SCN− as well.
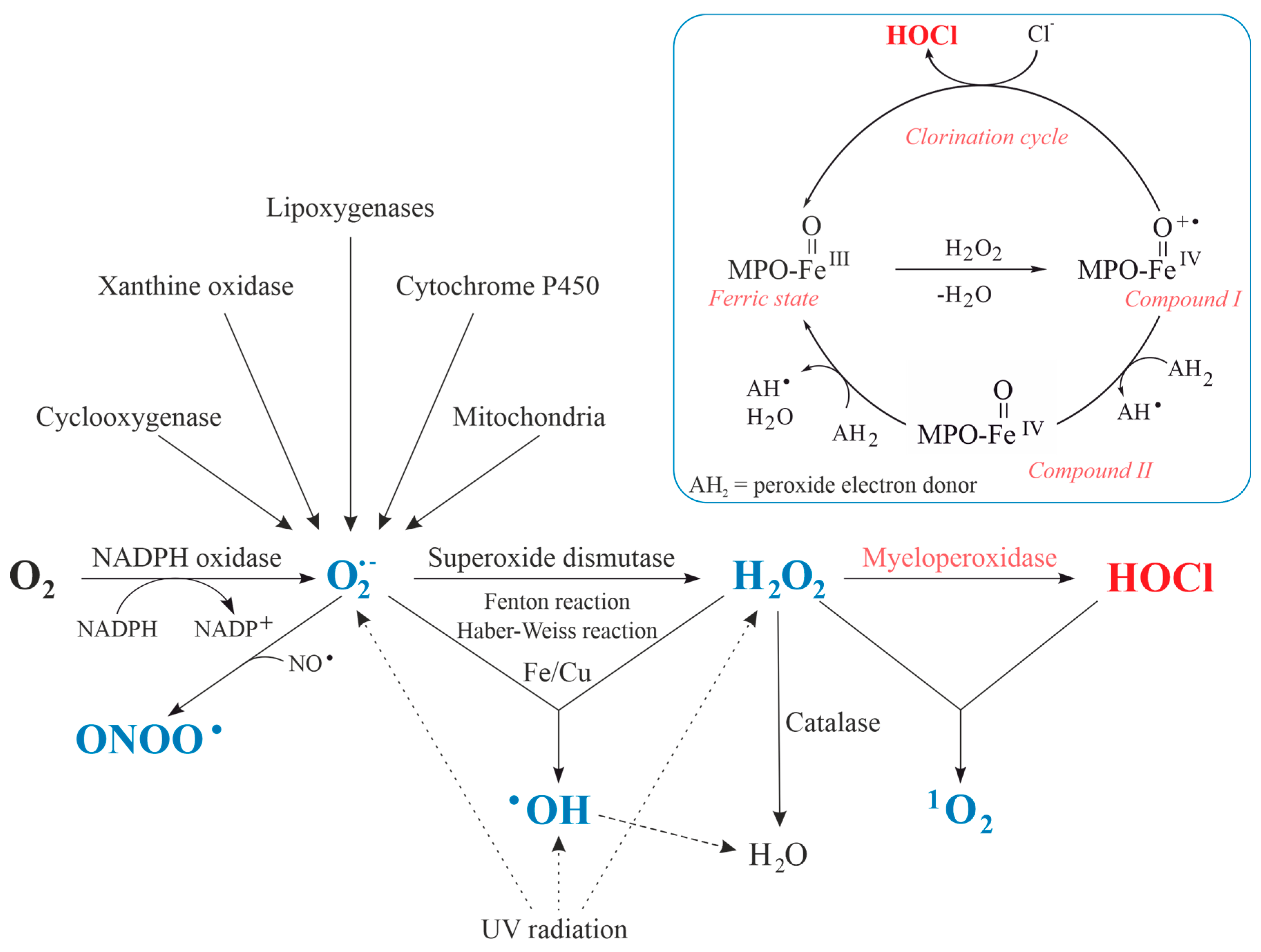
Figure 3.
Overview about reaction products of different functional groups in biomolecules by HOCl. The thickness of the arrows is indicative of the corresponding reaction rates.
Figure 3.
Overview about reaction products of different functional groups in biomolecules by HOCl. The thickness of the arrows is indicative of the corresponding reaction rates.

Figure 4.
Plot summarizing the second-order rate constants (on a log scale) for the reactions of HOCl with model compounds of protein, lipid, and carbohydrate components, nucleobases, and antioxidants. Reprinted from [89] with permission from Mary Ann Liebert, Inc. (New Rochelle, NY, USA) The indicated k values may differ significantly in dependence on the method how they were determined. Therefore, relative reactivities are most relevant.
Figure 4.
Plot summarizing the second-order rate constants (on a log scale) for the reactions of HOCl with model compounds of protein, lipid, and carbohydrate components, nucleobases, and antioxidants. Reprinted from [89] with permission from Mary Ann Liebert, Inc. (New Rochelle, NY, USA) The indicated k values may differ significantly in dependence on the method how they were determined. Therefore, relative reactivities are most relevant.

Figure 5.
Proposed mechanism of sulfilimine bond formation in collagen IV referring to [107]. The oxidation is based on the reaction with hypohalous acid (HOX; X = Cl, Br) and the sulfur or nitrogen in methionine (purple) or lysine/hydroxylysine (red) residues, respectively. Both mechanisms lead to the generation of the sulfilmine bond.
Figure 5.
Proposed mechanism of sulfilimine bond formation in collagen IV referring to [107]. The oxidation is based on the reaction with hypohalous acid (HOX; X = Cl, Br) and the sulfur or nitrogen in methionine (purple) or lysine/hydroxylysine (red) residues, respectively. Both mechanisms lead to the generation of the sulfilmine bond.
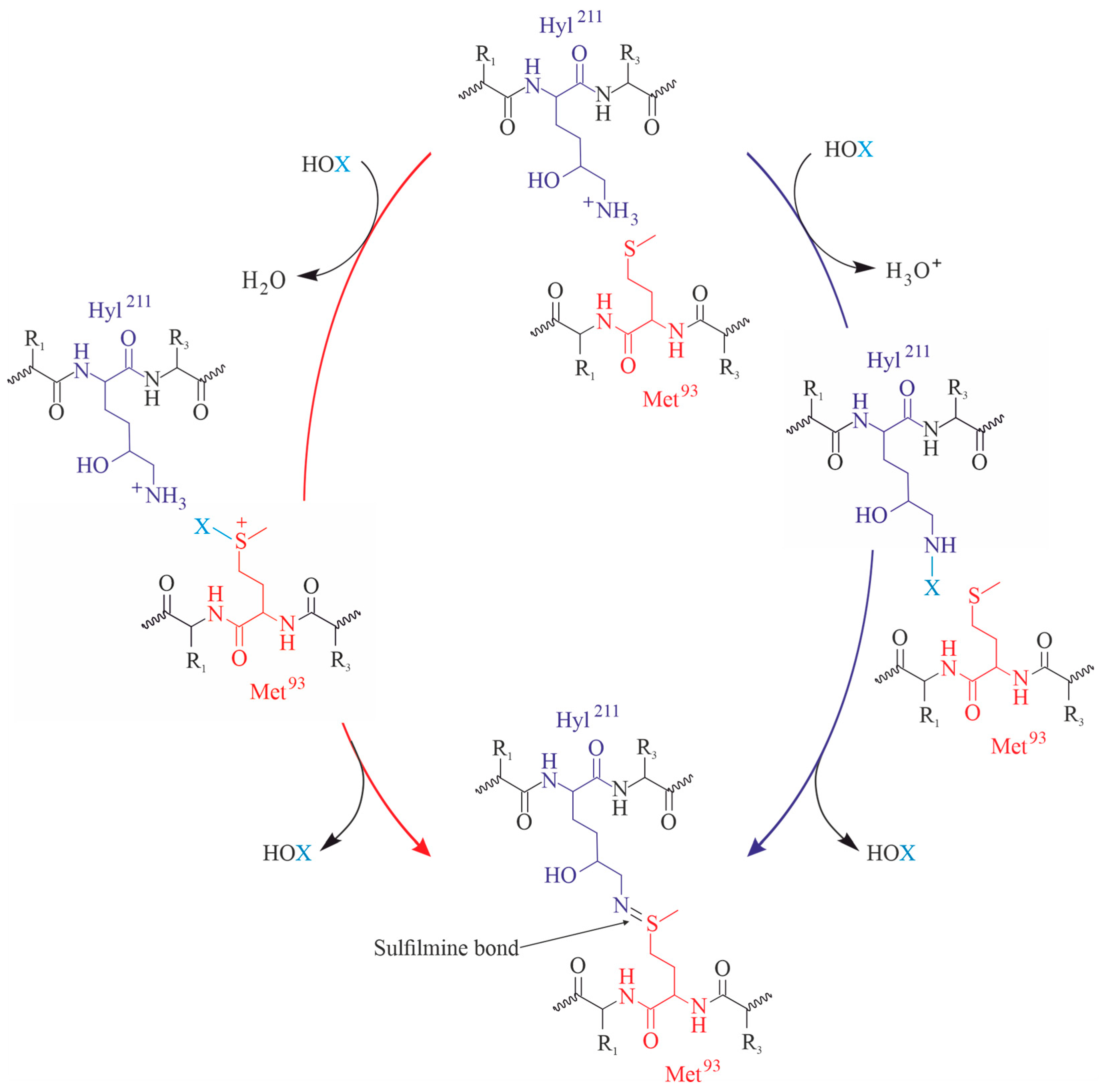
Figure 6.
Pathway of the reaction between the reagent HOCl and N-acetylglucosamine as one important monosaccharide within the hyaluronan chain.
Figure 6.
Pathway of the reaction between the reagent HOCl and N-acetylglucosamine as one important monosaccharide within the hyaluronan chain.

Figure 7.
HOCl-induced fragmentation of the HA moiety as one representative example of GAGs. Reproduced from the work of Rees et al. [121] with permission and with slight modifications.
Figure 7.
HOCl-induced fragmentation of the HA moiety as one representative example of GAGs. Reproduced from the work of Rees et al. [121] with permission and with slight modifications.
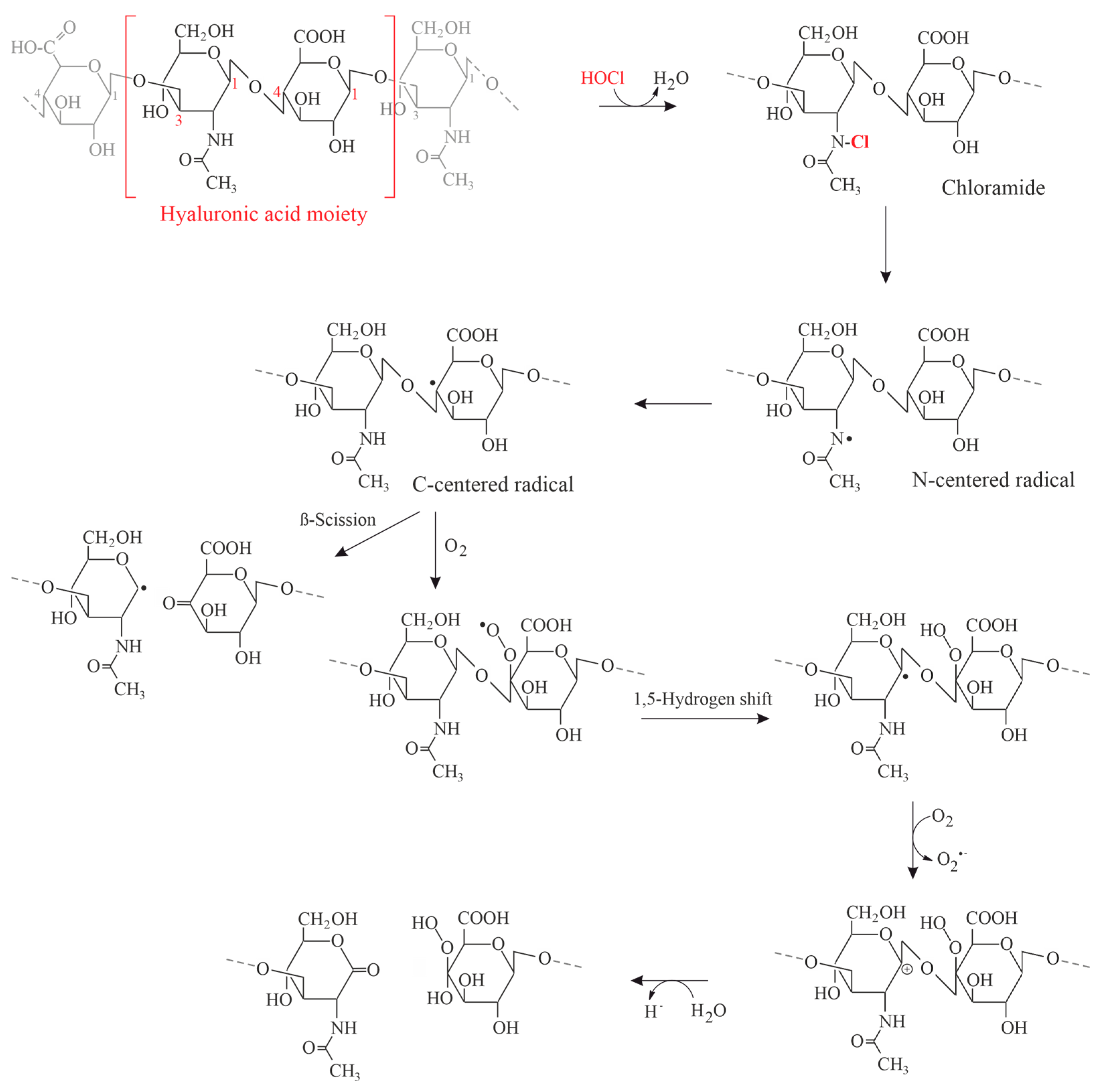
Figure 8.
Main lipidomic alterations described in several studies of rheumatoid arthritis. Reprinted from [158] under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Figure 8.
Main lipidomic alterations described in several studies of rheumatoid arthritis. Reprinted from [158] under the terms and conditions of the Creative Commons Attribution (CC BY) license.
Figure 9.
Schematic representation of Neutrophil Extracellular Traps (NETs) as network to kill invasive bacteria in accordance with [177]. Abbreviations: MPO, myeloperoxidase; NE, neutrophil elastase.
Figure 9.
Schematic representation of Neutrophil Extracellular Traps (NETs) as network to kill invasive bacteria in accordance with [177]. Abbreviations: MPO, myeloperoxidase; NE, neutrophil elastase.
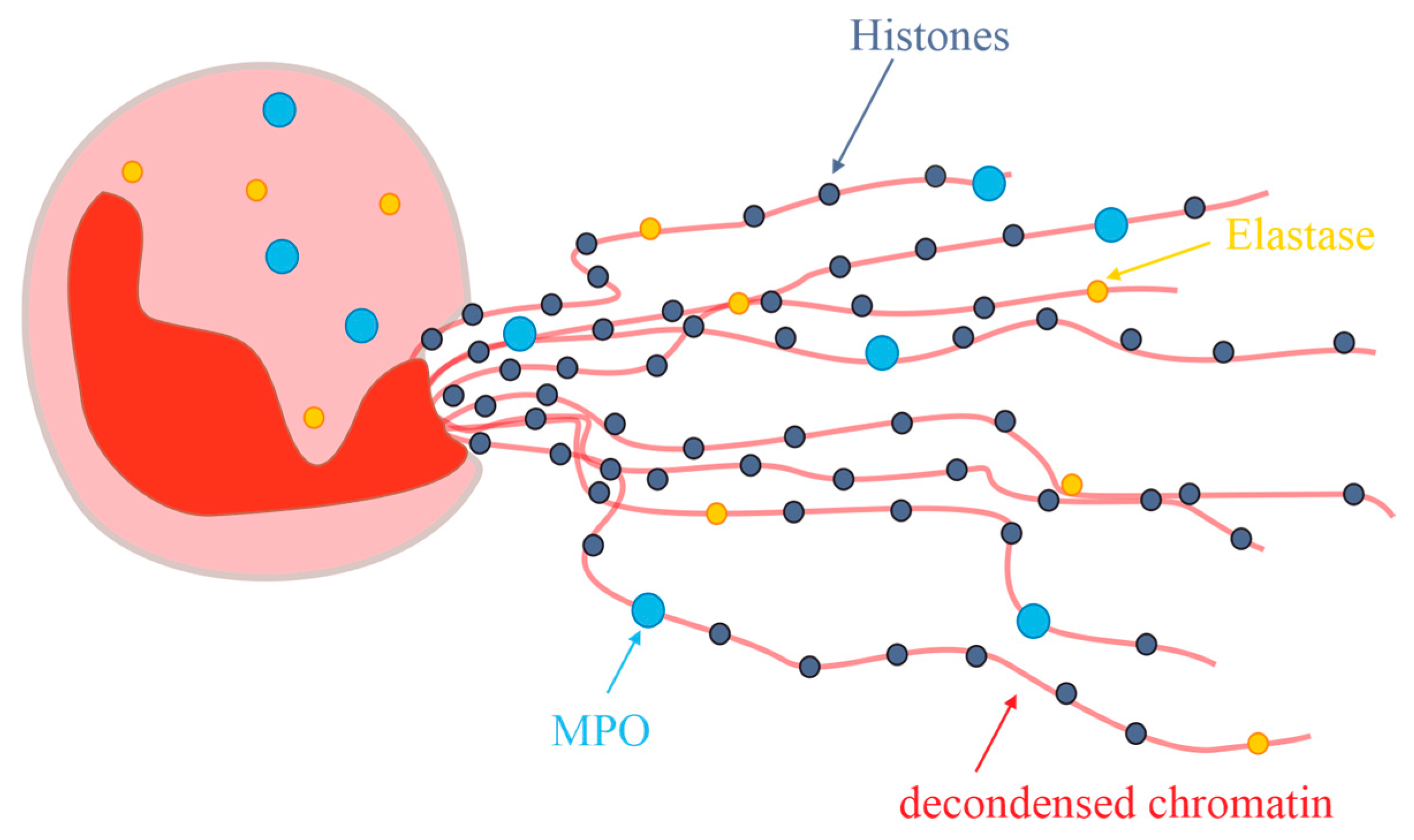
Figure 10.
Neutrophil multifaceted death pathways in inflammatory conditions. Neutrophils are generated in the bone marrow through granulopoiesis and subsequently enter the circulatory system. Depending on the specific microenvironment, neutrophils undergo various mechanisms of cell death. These mechanisms encompass both non-lytic apoptosis and lytic death modalities, including necroptosis, pyroptosis, ferroptosis, and NETosis. These lytic processes are accompanied by the release of cytotoxic cellular proteases, cell-free DNA, and chromatin into the microenvironment. Each death pathway operates through distinct molecular mechanisms and regulatory networks, ultimately resulting in either immunosuppressive or pro-inflammatory outcomes. Defects in the clearance of apoptotic neutrophils and the accumulation of cellular remnants contribute to the onset of inflammatory diseases and autoimmune disorders (Reprinted from [181] under the terms and conditions of the Creative Commons Attribution (CC BY) license). Abbreviations: DAMP, Damage-Associated Molecular Patterns; IL1-ꞵ, Interleukin 1-ꞵ; NET, Neutrophil Extracellular Traps; PE-OOH, Phosphatidylethanolamine Hydroperoxide (The detection of anti-phosphatidylethanolamine autoantibodies will not be discussed here) [183].
Figure 10.
Neutrophil multifaceted death pathways in inflammatory conditions. Neutrophils are generated in the bone marrow through granulopoiesis and subsequently enter the circulatory system. Depending on the specific microenvironment, neutrophils undergo various mechanisms of cell death. These mechanisms encompass both non-lytic apoptosis and lytic death modalities, including necroptosis, pyroptosis, ferroptosis, and NETosis. These lytic processes are accompanied by the release of cytotoxic cellular proteases, cell-free DNA, and chromatin into the microenvironment. Each death pathway operates through distinct molecular mechanisms and regulatory networks, ultimately resulting in either immunosuppressive or pro-inflammatory outcomes. Defects in the clearance of apoptotic neutrophils and the accumulation of cellular remnants contribute to the onset of inflammatory diseases and autoimmune disorders (Reprinted from [181] under the terms and conditions of the Creative Commons Attribution (CC BY) license). Abbreviations: DAMP, Damage-Associated Molecular Patterns; IL1-ꞵ, Interleukin 1-ꞵ; NET, Neutrophil Extracellular Traps; PE-OOH, Phosphatidylethanolamine Hydroperoxide (The detection of anti-phosphatidylethanolamine autoantibodies will not be discussed here) [183].
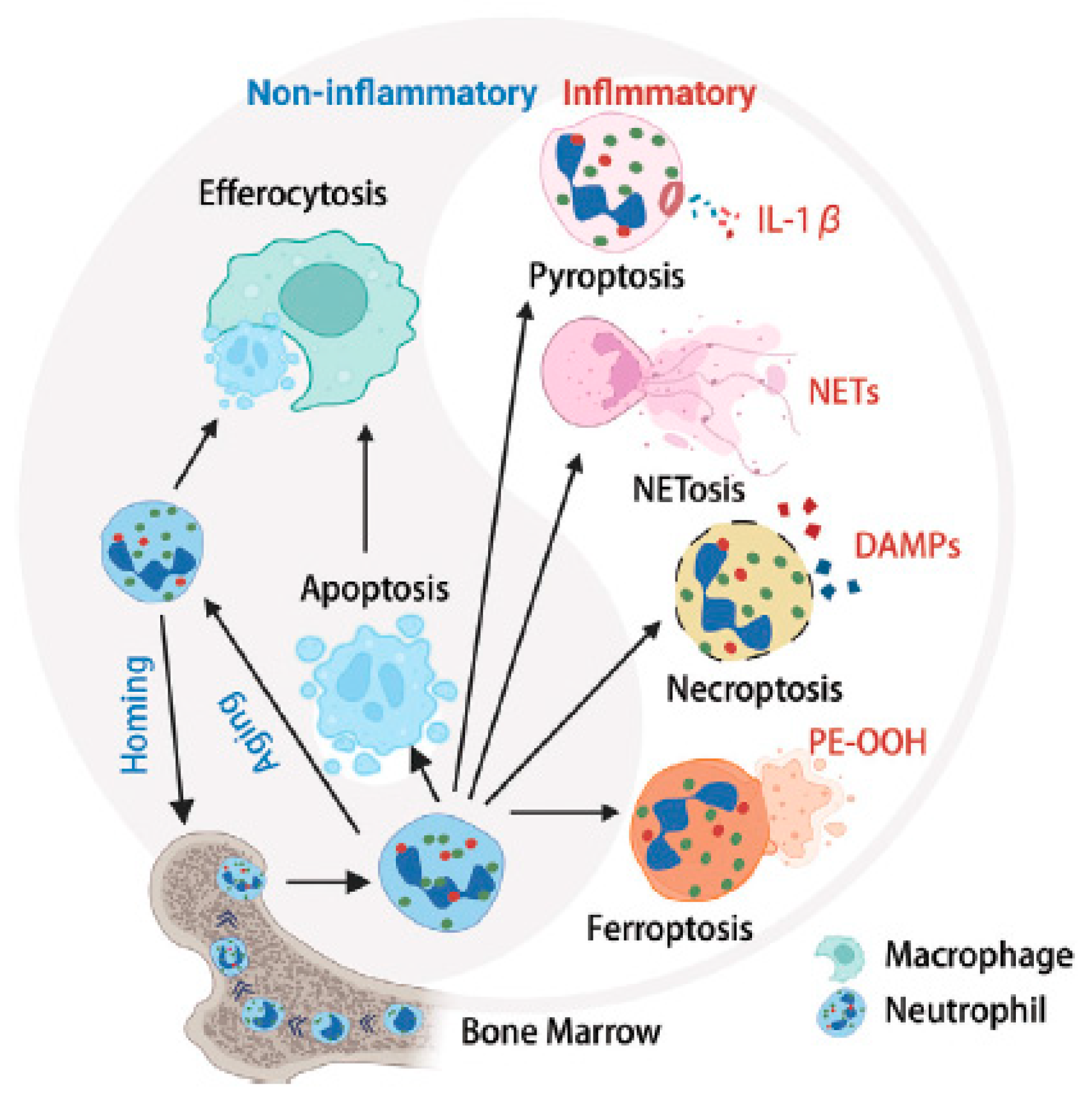
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Leopold, J.; Schiller, J. (Chemical) Roles of HOCl in Rheumatic Diseases. Antioxidants 2024, 13, 921. https://doi.org/10.3390/antiox13080921
AMA Style
Leopold J, Schiller J. (Chemical) Roles of HOCl in Rheumatic Diseases. Antioxidants. 2024; 13(8):921. https://doi.org/10.3390/antiox13080921
Chicago/Turabian StyleLeopold, Jenny, and Jürgen Schiller. 2024. "(Chemical) Roles of HOCl in Rheumatic Diseases" Antioxidants 13, no. 8: 921. https://doi.org/10.3390/antiox13080921
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.