
Immunization with Pooled Antigens for Clostridium perfringens Conferred Partial Protection against Experimental Necrotic Enteritis in Broiler Chickens

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Construction of Recombinant Chimeric NetB-CPA (NA) and FBA-Zm (FZ), and Cna Expression Vector
2.2. Expression and Purification of Recombinant C. perfringens Proteins
2.3. Bacterial Strain
2.4. Broiler Chick Husbandry and Experimental Design
2.5. In Vivo Evaluation of Vaccine Efficacy of Recombinant C. perfringens Proteins
2.6. Determination of Antibody Titers in Vaccinated Birds by ELISA
2.7. Statistical Analysis
3. Results
3.1. Mortality Post-Challenge in Vaccinated Groups
3.2. Jejunum Lesion Scores
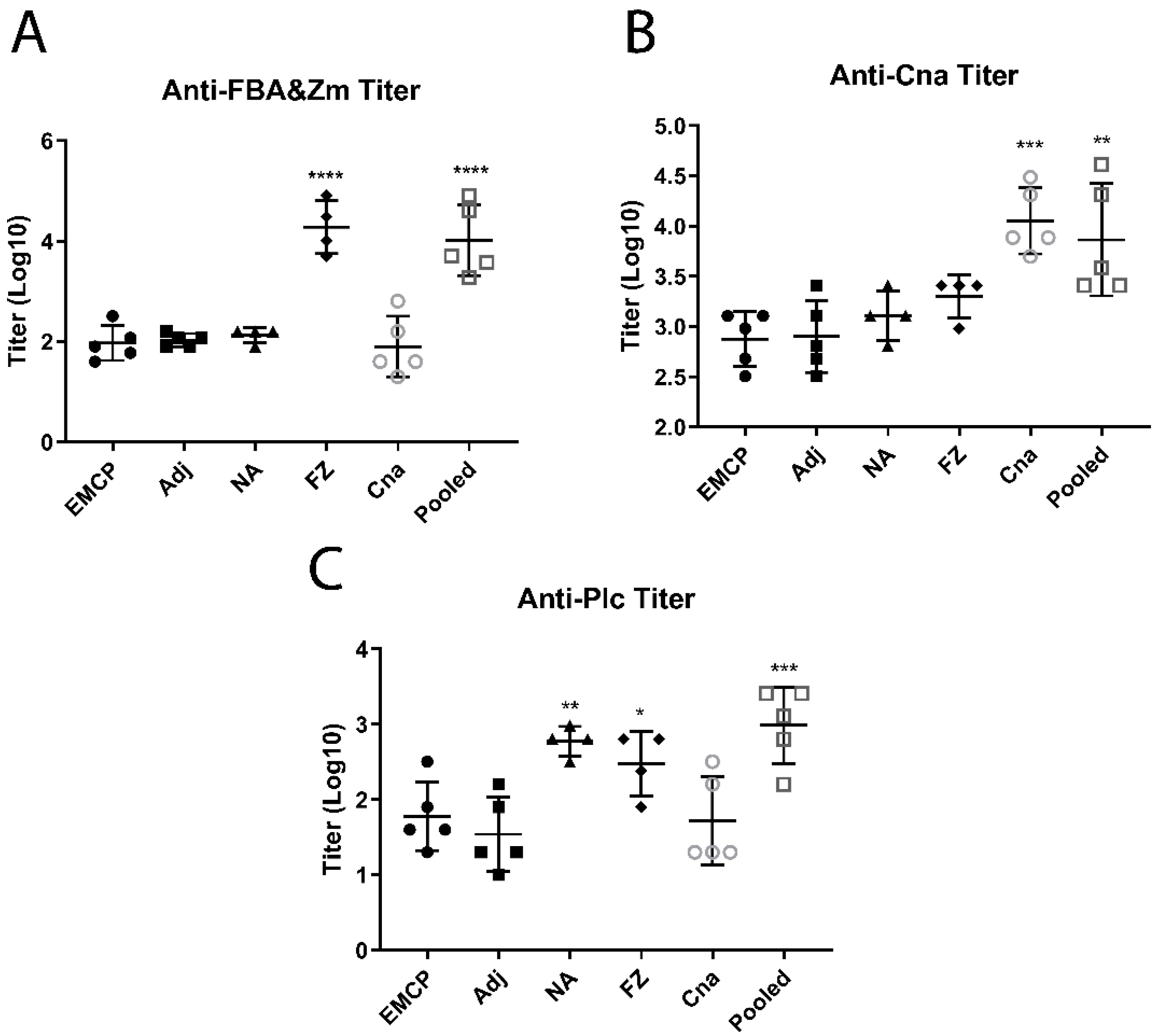
3.3. Determination of Antibody Titers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NE | Necrotic enteritis |
| CPA | Alpha-toxin |
| Plc | Phospholipase C (PLC) enzyme of alpha-toxin |
| NetB | Necrotic enteritis B-like toxin |
| FBA | Fructose-1,6-bisphosphate aldolase (FBA) |
| Zm | Zinc metalloprotease (Zm) |
| Cna | Collagen adhesion protein |
| AGP | Antibiotic growth promoters |
| EM | Eimeria maxima |
| CFU | Colony-forming units |
References
- Wade, B.; Keyburn, A. The true cost of necrotic enteritis. World Poult. 2015, 31, 16–17. [Google Scholar]
- Uzal, F.A.; Navarro, M.A.; Li, J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Gohari, I.M.; Navarro, M.A.; Li, J.; Shrestha, A.; Uzal, F.; McClane, B.A. Pathogenicity and virulence of Clostridium perfringens. Virulence 2021, 12, 723–753. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.F.; Smyth, J.A.; Shojadoost, B.; Vince, A. Experimental reproduction of necrotic enteritis in chickens: A review. Avian Pathol. 2016, 45, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: A critical review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef] [Green Version]
- Cooper, K.K.; Songer, J.G. Necrotic enteritis in chickens: A paradigm of enteric infection by Clostridium perfringens type A. Anaerobe 2009, 15, 55–60. [Google Scholar] [CrossRef]
- Thompson, D.R.; Parreira, V.R.; Kulkarni, R.R.; Prescott, J.F. Live attenuated vaccine-based control of necrotic enteritis of broiler chickens. Vet. Microbiol. 2006, 113, 25–34. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, P.; Wang, B.; Ma, B.; Wang, J. A combined Clostridium perfringens/Trueperella pyogenes inactivated vaccine induces complete immunoprotection in a mouse model. Biologicals 2017, 47, 1–10. [Google Scholar] [CrossRef]
- Wilde, S.; Jiang, Y.; Tafoya, A.M.; Horsman, J.; Yousif, M.; Vazquez, L.A.; Roland, K.L. Salmonella-vectored vaccine delivering three Clostridium perfringens antigens protects poultry against necrotic enteritis. PLoS ONE 2019, 14, e0197721. [Google Scholar] [CrossRef] [Green Version]
- Mishra, N.; Smyth, J.A. Oral vaccination of broiler chickens against necrotic enteritis using a non-virulent NetB positive strain of Clostridium perfringens type A. Vaccine 2017, 35 Pt B, 6858–6865. [Google Scholar] [CrossRef]
- Awad, M.M.; Bryant, A.E.; Stevens, D.L.; Rood, J.I. Virulence studies on chromosomal alpha-toxin and theta-toxin mutants constructed by allelic exchange provide genetic evidence for the essential role of alpha-toxin in Clostridium perfringens-mediated gas gangrene. Mol. Microbiol. 1995, 15, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Coursodon, C.F.; Trinh, H.T.; Mallozzi, M.; Vedantam, G.; Glock, R.; Songer, J. Clostridium perfringens alpha toxin is produced in the intestines of broiler chicks inoculated with an alpha toxin mutant. Anaerobe 2010, 16, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Lanckriet, A.; Gholamiandehkordi, A.R.; Pasmans, F.; Martel, A.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Origin of Clostridium perfringens isolates determines the ability to induce necrotic enteritis in broilers. Comp. Immunol. Microbiol. Infect. Dis. 2009, 32, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Shams, F.; Oldfield, N.J.; Wooldridge, K.G.; Turner, D.P. Fructose-1,6-bisphosphate aldolase (FBA)-a conserved glycolytic enzyme with virulence functions in bacteria: ‘ill met by moonlight’. Biochem. Soc. Trans. 2014, 42, 1792–1795. [Google Scholar] [CrossRef]
- Wade, B.; Keyburn, A.L.; Haring, V.; Ford, M.; Rood, J.I.; Moore, R.J. Two putative zinc metalloproteases contribute to the virulence of Clostridium perfringens strains that cause avian necrotic enteritis. J. Vet. Diagn. Investig. 2020, 32, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Lepp, D.; Zhou, Y.; Ojha, S.; Gohari, I.M.; Carere, J.; Yang, C.; Prescott, J.F.; Gong, J. Clostridium perfringens Produces an Adhesive Pilus Required for the Pathogenesis of Necrotic Enteritis in Poultry. J. Bacteriol. 2021, 203, e00578-20. [Google Scholar] [CrossRef]
- Jiang, Y.; Kulkarni, R.; Parreira, V.R.; Prescott, J.F. Immunization of broiler chickens against clostridium perfringens–Induced necrotic enteritis using purified recombinant immunogenic proteins. Avian Dis. 2009, 53, 409–415. [Google Scholar] [CrossRef]
- Jang, S.I.; Lillehoj, H.S.; Lee, S.-H.; Lee, K.-W.; Lillehoj, E.P.; Hong, Y.H.; An, D.-J.; Jeong, W.; Chun, J.-E.; Bertrand, F.; et al. Vaccination with Clostridium perfringens recombinant proteins in combination with Montanide™ ISA 71 VG adjuvant increases protection against experimental necrotic enteritis in commercial broiler chickens. Vaccine 2012, 30, 5401–5406. [Google Scholar] [CrossRef]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Immunization of broiler chickens against Clostridium perfringens-induced necrotic enteritis. Clin. Vaccine Immunol. 2007, 14, 1070–1077. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Clostridium perfringens antigens recognized by broiler chickens immune to necrotic enteritis. Clin. Vaccine Immunol. 2006, 13, 1358–1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wade, B.; Keyburn, A.L.; Haring, V.; Ford, M.; Rood, J.I.; Moore, R.J. The adherent abilities of Clostridium perfringens strains are critical for the pathogenesis of avian necrotic enteritis. Vet. Microbiol. 2016, 197, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Wade, B.; Keyburn, A.; Seemann, T.; Rood, J.; Moore, R. Binding of Clostridium perfringens to collagen correlates with the ability to cause necrotic enteritis in chickens. Vet. Microbiol. 2015, 180, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Mo, H.; Willingham, C.; Wang, S.; Park, J.-Y.; Kong, W.; Roland, K.L.; Curtiss, R. Protection Against Necrotic Enteritis in Broiler Chickens by Regulated Delayed Lysis Salmonella Vaccines. Avian Dis. 2015, 59, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.R.; Parreira, V.R.; Sharif, S.; Prescott, J.F. Oral immunization of broiler chickens against necrotic enteritis with an attenuated Salmonella vaccine vector expressing Clostridium perfringens antigens. Vaccine 2008, 26, 4194–4203. [Google Scholar] [CrossRef]
- Lepp, D.; Ojha, S.; Gohari, I.M.; Chakravarty, B.; Prescott, J.; Gong, J. Immunization with subunits of a novel pilus produced by virulent Clostridium perfringens strains confers partial protection against necrotic enteritis in chickens. Vet. Microbiol. 2019, 230, 7–13. [Google Scholar] [CrossRef]
- Gu, C.; Lillehoj, H.S.; Sun, Z.; Lee, Y.; Zhao, H.; Xianyu, Z.; Yan, X.; Wang, Y.; Lin, S.; Li, C.; et al. Characterization of Virulent netB(+)/tpeL(+) Clostridium perfringens Strains from Necrotic Enteritis-Affected Broiler Chicken Farms. Avian Dis. 2019, 63, 461–467. [Google Scholar] [CrossRef]
- Liu, L.; Yan, X.; Lillehoj, H.; Sun, Z.; Zhao, H.; Xianyu, Z.; Lee, Y.; Melville, S.; Gu, C.; Wang, Y.; et al. Comparison of the Pathogenicity of Five Clostridium perfringens Isolates Using an Eimeria maxima Coinfection Necrotic Enteritis Disease Model in Commercial Broiler Chickens. Avian Dis. 2020, 64, 386–392. [Google Scholar] [CrossRef]
- Li, C.; Lillehoj, H.S.; Gadde, U.D.; Ritter, D.; Oh, S. Characterization of Clostridium perfringens Strains Isolated from Healthy and Necrotic Enteritis-Afflicted Broiler Chickens. Avian Dis. 2017, 61, 178–185. [Google Scholar] [CrossRef]
- Lee, K.-W.; Lillehoj, H.S.; Jeong, W.; Jeoung, H.Y.; An, D.J. Avian necrotic enteritis: Experimental models, host immunity, pathogenesis, risk factors, and vaccine development. Poult. Sci. 2011, 90, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.S.; Jang, S.I.; Panebra, A.; Lillehoj, E.P.; Dupuis, L.; Ben Arous, J.; Lee, S.K.; Oh, S.T. In ovo vaccination using Eimeria profilin and Clostridium perfringens NetB proteins in Montanide IMS adjuvant increases protective immunity against experimentally-induced necrotic enteritis. Asian-Australas. J. Anim. Sci. 2017, 30, 1478–1485. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-W.; Lillehoj, H.-S.; Jang, S.-I.; Lee, S.-H.; Bautista, D.A.; Ritter, G.D.; Lillehoj, E.P.; Siragusa, G.R. Comparison of live Eimeria vaccination with in-feed salinomycin on growth and immune status in broiler chickens. Res. Vet. Sci. 2013, 95, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.F. The prevention of experimentally induced necrotic enteritis in chickens by avoparcin. Avian Dis. 1979, 23, 1072. [Google Scholar] [CrossRef]
- Alizadeh, M.; Shojadoost, B.; Boodhoo, N.; Astill, J.; Taha-Abdelaziz, K.; Hodgins, D.C.; Kulkarni, R.R.; Sharif, S. Necrotic enteritis in chickens: A review of pathogenesis, immune responses and prevention, focusing on probiotics and vaccination. Anim. Health Res. Rev. 2021, 22, 147–162. [Google Scholar] [CrossRef]
- Sarmah, H.; Hazarika, R.; Tamuly, S.; Deka, P.; Manoharan, S.; Sharma, R.K. Evaluation of different antigenic preparations against necrotic enteritis in broiler birds using a novel Clostridium perfringens type G strain. Anaerobe 2021, 70, 102377. [Google Scholar] [CrossRef]
- Mot, D.; Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Progress and problems in vaccination against necrotic enteritis in broiler chickens. Avian Pathol. 2014, 43, 290–300. [Google Scholar] [CrossRef] [Green Version]
- Goshadrou, F.; Langroudi, R.P.; Riazi, A.; Rostami, A.; Bathaie, S.Z.; Amani, J.; Ahmadian, G. Design and expression of a chimeric vaccine candidate for avian necrotic enteritis. Protein Eng. Des. Sel. 2017, 30, 39–45. [Google Scholar]
- Katalani, C.; Ahmadian, G.; Nematzadeh, G.; Amani, J.; Ehsani, P.; Razmyar, J.; Kiani, G. Immunization with oral and parenteral subunit chimeric vaccine candidate confers protection against Necrotic Enteritis in chickens. Vaccine 2020, 38, 7284–7291. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Group No. | Group Name | Bird Number | Vaccination (D4, D15) | Challenge | Sacrifice (D28) | |
|---|---|---|---|---|---|---|
| EM(D22) | CP (D26) | |||||
| 1 | Naive | 13 | PBS | PBS | BYC | S |
| 2 | Challenge Control (EMCP) | 13 | PBS | EM | CP | S |
| 3 | Adjuvant control (Adj) | 13 | Adj + PBS | EM | CP | S |
| 4 | Cna | 13 | Adj + Cna | EM | CP | S |
| 5 | FZ | 13 | Adj + FZ | EM | CP | S |
| 6 | NA | 13 | Adj + NA | EM | CP | S |
| 7 | Pooled (NA + FZ + Cna) | 13 | Adj + NA + FZ + Cna | EM | CP | S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, B.; Sun, Z.; Lu, M.; Lillehoj, H.; Lee, Y.; Liu, L.; Yan, X.; Yang, D.A.; Li, C. Immunization with Pooled Antigens for Clostridium perfringens Conferred Partial Protection against Experimental Necrotic Enteritis in Broiler Chickens. Vaccines 2022, 10, 979. https://doi.org/10.3390/vaccines10060979
Yuan B, Sun Z, Lu M, Lillehoj H, Lee Y, Liu L, Yan X, Yang DA, Li C. Immunization with Pooled Antigens for Clostridium perfringens Conferred Partial Protection against Experimental Necrotic Enteritis in Broiler Chickens. Vaccines. 2022; 10(6):979. https://doi.org/10.3390/vaccines10060979
Chicago/Turabian StyleYuan, Baohong, Zhifeng Sun, Mingmin Lu, Hyun Lillehoj, Youngsub Lee, Liheng Liu, Xianghe Yan, Danchen Aaron Yang, and Charles Li. 2022. "Immunization with Pooled Antigens for Clostridium perfringens Conferred Partial Protection against Experimental Necrotic Enteritis in Broiler Chickens" Vaccines 10, no. 6: 979. https://doi.org/10.3390/vaccines10060979
APA StyleYuan, B., Sun, Z., Lu, M., Lillehoj, H., Lee, Y., Liu, L., Yan, X., Yang, D. A., & Li, C. (2022). Immunization with Pooled Antigens for Clostridium perfringens Conferred Partial Protection against Experimental Necrotic Enteritis in Broiler Chickens. Vaccines, 10(6), 979. https://doi.org/10.3390/vaccines10060979