
Evaluation of Three Candidate Live-Attenuated Salmonella enterica Serovar Typhimurium Vaccines to Prevent Non-Typhoidal Salmonella Infection in an Infant Mouse Model
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Construction of CVD 1921 ΔhtrA
2.3. Generation of Bioluminescent S. Typhimurium Strains
2.4. In Vivo Bioluminescence Imaging of S. Typhimurium Vaccines in Infant and Adult Mice
2.5. Mouse Immunization and Protection against Challenge
2.6. Measurement of Serum and Fecal Antibodies
2.7. Statistical Analyses
3. Results
3.1. Immunogenicity and Vaccine Efficacy of S. Typhimurium Vaccines in Adult BALB/c Mice
3.2. Immunogenicity and Vaccine Efficacy of S. Typhimurium Vaccines in Infant BALB/c Mice Following Intranasal Administration
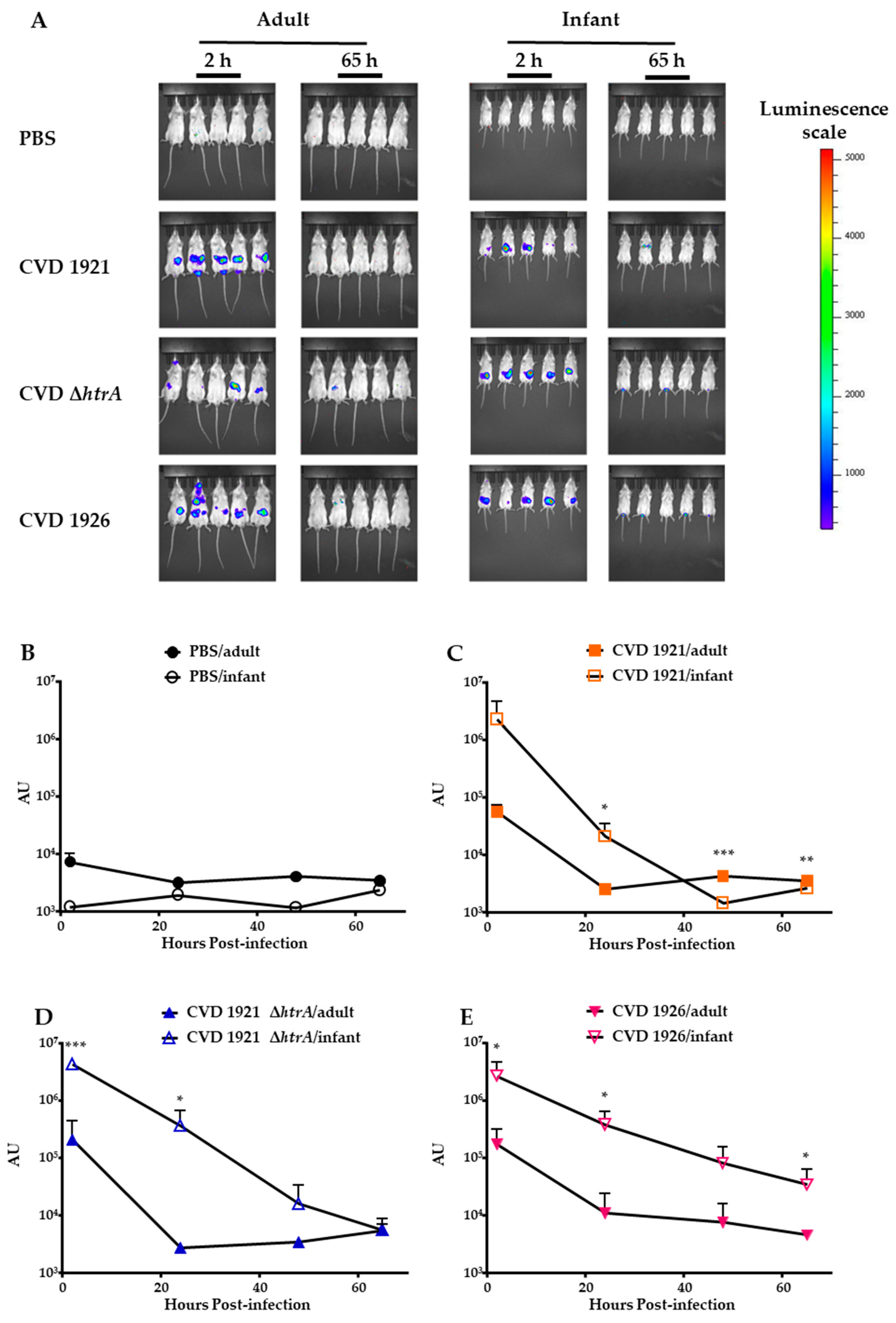
3.3. In Vivo Clearance of S. Typhimurium Vaccines in Infant and Adult BALB/c Mice Following Peroral Administration
3.4. Effect of Age and Immunization Schedule on Immunogenicity and Protection Following Peroral Administration
4. Discussion
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- GBD 2019 Diseases and Injuries Collaborators. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar] [CrossRef]
- Majowicz, S.E.; Musto, J.; Scallan, E.; Angulo, F.J.; Kirk, M.; O’Brien, S.J.; Jones, T.F.; Fazil, A.; Hoekstra, R.M. The global burden of non-typhoidal Salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef]
- Collins, J.P.; Shah, H.J.; Weller, D.L.; Ray, L.C.; Smith, K.; McGuire, S.; Trevejo, R.T.; Jervis, R.H.; Vugia, D.J.; Rissman, T.; et al. Preliminary incidence and trends of infections caused by pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2016–2021. MMWR Morb. Mortal. Wkly. Rep. 2022, 71, 1260–1264. [Google Scholar] [CrossRef] [PubMed]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne illness acquired in the United States--major pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2015–2018. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 369–373. [Google Scholar] [CrossRef] [PubMed]
- The Division of Foodborne Waterborne and Environmental Diseases (DFWED). National Enteric Disease Surveillance: Salmonella Annual Report, 2016; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018; p. 87.
- Mastroeni, P.; Chabalgoity, J.A.; Dunstan, S.J.; Maskell, D.J.; Dougan, G. Salmonella: Immune responses and vaccines. Vet. J. 2001, 161, 132–164. [Google Scholar] [CrossRef]
- Tennant, S.M.; Muhsen, K.; Pasetti, M.F. Gut immunology and oral vaccination. In Molecular Vaccines: From Prophylaxis to Therapy—Volume 1; Giese, M., Ed.; Springer: Vienna, Austria, 2013; pp. 59–84. [Google Scholar]
- Sztein, M.B. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica serovar Typhi strains used as live oral vaccines in humans. Clin. Infect. Dis. 2007, 45 (Suppl. S1), S15–S19. [Google Scholar] [CrossRef]
- Laouar, A. Maternal leukocytes and infant immune programming during breastfeeding. Trends Immunol. 2020, 41, 225–239. [Google Scholar] [CrossRef]
- Saso, A.; Kampmann, B. Vaccine responses in newborns. Semin. Immunopathol. 2017, 39, 627–642. [Google Scholar] [CrossRef]
- GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect. Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef]
- Tate, J.E.; Mutuc, J.D.; Panozzo, C.A.; Payne, D.C.; Cortese, M.M.; Cortes, J.E.; Yen, C.; Esposito, D.H.; Lopman, B.A.; Patel, M.M.; et al. Sustained decline in rotavirus detections in the United States following the introduction of rotavirus vaccine in 2006. Pediatr. Infect. Dis. J. 2011, 30, S30–S34. [Google Scholar] [CrossRef] [PubMed]
- Mijatovic-Rustempasic, S.; Jaimes, J.; Perkins, C.; Ward, M.L.; Esona, M.D.; Gautam, R.; Lewis, J.; Sturgeon, M.; Panjwani, J.; Bloom, G.A.; et al. Rotavirus strain trends in United States, 2009–2016: Results from the National Rotavirus Strain Surveillance System (NRSSS). Viruses 2022, 14, 1775. [Google Scholar] [CrossRef] [PubMed]
- Gastanaduy, P.A.; Curns, A.T.; Parashar, U.D.; Lopman, B.A. Gastroenteritis hospitalizations in older children and adults in the United States before and after implementation of infant rotavirus vaccination. JAMA 2013, 310, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, A.S.; Garon, J.; Seib, K.; Orenstein, W.A. Polio vaccination: Past, present and future. Future Microbiol. 2015, 10, 791–808. [Google Scholar] [CrossRef]
- Levine, M.M.; Ferreccio, C.; Cryz, S.; Ortiz, E. Comparison of enteric-coated capsules and liquid formulation of Ty21a typhoid vaccine in randomised controlled field trial. Lancet 1990, 336, 891–894. [Google Scholar] [CrossRef]
- Amicizia, D.; Arata, L.; Zangrillo, F.; Panatto, D.; Gasparini, R. Overview of the impact of typhoid and paratyphoid fever. Utility of Ty21a vaccine (Vivotif(R)). J. Prev. Med. Hyg. 2017, 58, E1–E8. [Google Scholar]
- Shi, H.; Wang, S.; Roland, K.L.; Gunn, B.M.; Curtiss, R., 3rd. Immunogenicity of a live recombinant Salmonella enterica serovar Typhimurium vaccine expressing pspA in neonates and infant mice born from naive and immunized mothers. Clin. Vaccine Immunol. 2010, 17, 363–371. [Google Scholar] [CrossRef]
- Galen, J.E.; Curtiss, R., 3rd. The delicate balance in genetically engineering live vaccines. Vaccine 2014, 32, 4376–4385. [Google Scholar] [CrossRef]
- Achtman, M.; Wain, J.; Weill, F.X.; Nair, S.; Zhou, Z.; Sangal, V.; Krauland, M.G.; Hale, J.L.; Harbottle, H.; Uesbeck, A.; et al. Multilocus sequence typing as a replacement for serotyping in Salmonella enterica. PLoS Pathog. 2012, 8, e1002776. [Google Scholar] [CrossRef]
- Tennant, S.M.; Wang, J.Y.; Galen, J.E.; Simon, R.; Pasetti, M.F.; Gat, O.; Levine, M.M. Engineering and preclinical evaluation of attenuated nontyphoidal Salmonella strains serving as live oral vaccines and as reagent strains. Infect. Immun. 2011, 79, 4175–4185. [Google Scholar] [CrossRef]
- Higginson, E.E.; Ramachandran, G.; Panda, A.; Shipley, S.T.; Kriel, E.H.; DeTolla, L.J.; Lipsky, M.; Perkins, D.J.; Salerno-Goncalves, R.; Sztein, M.B.; et al. Improved tolerability of a Salmonella enterica serovar Typhimurium live-attenuated vaccine strain achieved by balancing inflammatory potential with immunogenicity. Infect. Immun. 2018, 86, 1110–1128. [Google Scholar] [CrossRef]
- Ault, A.; Tennant, S.M.; Gorres, J.P.; Eckhaus, M.; Sandler, N.G.; Roque, A.; Livio, S.; Bao, S.; Foulds, K.E.; Kao, S.F.; et al. Safety and tolerability of a live oral Salmonella Typhimurium vaccine candidate in SIV-infected nonhuman primates. Vaccine 2013, 31, 5879–5888. [Google Scholar] [CrossRef] [PubMed]
- Higginson, E.E.; Panda, A.; Toapanta, F.R.; Terzi, M.C.; Jones, J.A.; Sen, S.; Permala-Booth, J.; Pasetti, M.F.; Sztein, M.B.; DeTolla, L.; et al. Immunogenicity and efficacy of live-attenuated Salmonella Typhimurium vaccine candidate CVD 1926 in a rhesus macaque model of gastroenteritis. Infect. Immun. 2021, 89, e0008721. [Google Scholar] [CrossRef]
- Allen, J.C.; Toapanta, F.R.; Baliban, S.M.; Sztein, M.B.; Tennant, S.M. Reduced immunogenicity of a live Salmonella enterica serovar Typhimurium vaccine in aged mice. Front. Immunol. 2023, 14, 1190339. [Google Scholar] [CrossRef] [PubMed]
- Chatfield, S.N.; Strahan, K.; Pickard, D.; Charles, I.G.; Hormaeche, C.E.; Dougan, G. Evaluation of Salmonella Typhimurium strains harbouring defined mutations in htrA and aroA in the murine salmonellosis model. Microb. Pathog. 1992, 12, 145–151. [Google Scholar] [CrossRef]
- Baumler, A.J.; Kusters, J.G.; Stojiljkovic, I.; Heffron, F. Salmonella Typhimurium loci involved in survival within macrophages. Infect. Immun. 1994, 62, 1623–1630. [Google Scholar] [CrossRef]
- Tennant, S.M.; Diallo, S.; Levy, H.; Livio, S.; Sow, S.O.; Tapia, M.; Fields, P.I.; Mikoleit, M.; Tamboura, B.; Kotloff, K.L.; et al. Identification by PCR of non-typhoidal Salmonella enterica serovars associated with invasive infections among febrile patients in Mali. PLoS Negl. Trop. Dis. 2010, 4, e621. [Google Scholar] [CrossRef] [PubMed]
- Tapia, M.D.; Tennant, S.M.; Bornstein, K.; Onwuchekwa, U.; Tamboura, B.; Maiga, A.; Sylla, M.B.; Sissoko, S.; Kourouma, N.; Toure, A.; et al. Invasive nontyphoidal Salmonella infections among children in Mali, 2002–2014: Microbiological and epidemiologic features guide vaccine development. Clin. Infect. Dis. 2015, 61 (Suppl. S4), S332–S338. [Google Scholar] [CrossRef]
- Ramachandran, G.; Panda, A.; Higginson, E.E.; Ateh, E.; Lipsky, M.M.; Sen, S.; Matson, C.A.; Permala-Booth, J.; DeTolla, L.J.; Tennant, S.M. Virulence of invasive Salmonella Typhimurium ST313 in animal models of infection. PLoS Negl. Trop. Dis. 2017, 11, e0005697. [Google Scholar] [CrossRef]
- Hegerle, N.; Bose, J.; Ramachandran, G.; Galen, J.E.; Levine, M.M.; Simon, R.; Tennant, S.M. Overexpression of O-polysaccharide chain length regulators in Gram-negative bacteria using the Wzx-/Wzy-dependent pathway enhances production of defined modal length O-polysaccharide polymers for use as haptens in glycoconjugate vaccines. J. Appl. Microbiol. 2018, 125, 575–585. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef]
- Morin, C.E.; Kaper, J.B. Use of stabilized luciferase-expressing plasmids to examine in vivo-induced promoters in the Vibrio cholerae vaccine strain CVD 103-HgR. FEMS Immunol. Med. Microbiol. 2009, 57, 69–79. [Google Scholar] [CrossRef]
- Baliban, S.M.; Yang, M.; Ramachandran, G.; Curtis, B.; Shridhar, S.; Laufer, R.S.; Wang, J.Y.; Van Druff, J.; Higginson, E.E.; Hegerle, N.; et al. Development of a glycoconjugate vaccine to prevent invasive Salmonella Typhimurium infections in sub-Saharan Africa. PLoS Negl. Trop. Dis. 2017, 11, e0005493. [Google Scholar] [CrossRef]
- Sears, K.T.; Tennant, S.M.; Reymann, M.K.; Simon, R.; Konstantopoulos, N.; Blackwelder, W.C.; Barry, E.M.; Pasetti, M.F. Bioactive immune components of anti-diarrheagenic enterotoxigenic Escherichia coli hyperimmune bovine colostrum products. Clin. Vaccine Immunol. 2017, 24, e00186-16. [Google Scholar] [CrossRef]
- Meyer, P.N.; Wilmes-Riesenberg, M.R.; Stathopoulos, C.; Curtiss, R., 3rd. Virulence of a Salmonella typhimurium OmpD mutant. Infect. Immun. 1998, 66, 387–390. [Google Scholar] [CrossRef]
- Galan, J.E.; Curtiss, R., 3rd. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells. Proc. Natl. Acad. Sci. USA 1989, 86, 6383–6387. [Google Scholar] [CrossRef]
- Wheeler, T.T.; Hodgkinson, A.J.; Prosser, C.G.; Davis, S.R. Immune components of colostrum and milk--a historical perspective. J. Mammary Gland. Biol. Neoplasia 2007, 12, 237–247. [Google Scholar] [CrossRef]
- Contag, C.H.; Contag, P.R.; Mullins, J.I.; Spilman, S.D.; Stevenson, D.K.; Benaron, D.A. Photonic detection of bacterial pathogens in living hosts. Mol. Microbiol. 1995, 18, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Wellawa, D.H.; Lam, P.S.; White, A.P.; Allan, B.; Koster, W. Characterization of colonization kinetics and virulence potential of Salmonella Enteritidis in chickens by photonic detection. Front. Vet. Sci. 2022, 9, 948448. [Google Scholar] [CrossRef] [PubMed]
- Heine, S.J.; Franco-Mahecha, O.L.; Chen, X.; Choudhari, S.; Blackwelder, W.C.; van Roosmalen, M.L.; Leenhouts, K.; Picking, W.L.; Pasetti, M.F. Shigella IpaB and IpaD displayed on L. lactis bacterium-like particles induce protective immunity in adult and infant mice. Immunol. Cell Biol. 2015, 93, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Heine, S.J.; Franco-Mahecha, O.L.; Sears, K.T.; Drachenberg, C.B.; van Roosmalen, M.L.; Leenhouts, K.; Picking, W.L.; Pasetti, M.F. A combined YopB and LcrV subunit vaccine elicits protective immunity against Yersinia infection in adult and infant mice. J. Immunol. 2019, 202, 2005–2016. [Google Scholar] [CrossRef] [PubMed]
- Boyd, M.A.; Tennant, S.M.; Saague, V.A.; Simon, R.; Muhsen, K.; Ramachandran, G.; Cross, A.S.; Galen, J.E.; Pasetti, M.F.; Levine, M.M. Serum bactericidal assays to evaluate typhoidal and nontyphoidal Salmonella vaccines. Clin. Vaccine Immunol. 2014, 21, 712–721. [Google Scholar] [CrossRef]
- Ramachandran, G.; Boyd, M.A.; MacSwords, J.; Higginson, E.E.; Simon, R.; Galen, J.E.; Pasetti, M.F.; Levine, M.M.; Tennant, S.M. Opsonophagocytic assay to evaluate immunogenicity of nontyphoidal Salmonella vaccines. Clin. Vaccine Immunol. 2016, 23, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Fresnay, S.; McArthur, M.A.; Magder, L.S.; Darton, T.C.; Jones, C.; Waddington, C.S.; Blohmke, C.J.; Angus, B.; Levine, M.M.; Pollard, A.J.; et al. Importance of Salmonella Typhi-responsive CD8+ T cell immunity in a human typhoid fever challenge model. Front. Immunol. 2017, 8, 208. [Google Scholar] [CrossRef] [PubMed]
- Benoun, J.M.; Peres, N.G.; Wang, N.; Pham, O.H.; Rudisill, V.L.; Fogassy, Z.N.; Whitney, P.G.; Fernandez-Ruiz, D.; Gebhardt, T.; Pham, Q.M.; et al. Optimal protection against Salmonella infection requires noncirculating memory. Proc. Natl. Acad. Sci. USA 2018, 115, 10416–10421. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Characteristics * | Reference |
|---|---|---|
| S. Typhimurium strain I77 | Clinical isolate, sequence type 19 | [29,30,31] |
| CVD 1921 | I77 ∆guaBA ∆clpP | [22] |
| CVD 1921 ∆htrA | I77 ΔguaBA ΔclpP ΔhtrA | This work |
| CVD 1926 | I77 ΔguaBA ΔclpP ΔpipA ΔhtrA | [23] |
| CVD 1925 (pSEC10-wzzB) | I77 ∆guaBA ∆clpP ∆fliD ∆fljB (pSEC10-wzzB) | [22,32] |
| pKD13 | KanR template plasmid | [33] |
| pKD46 | AmpR, expresses red recombinase | [33] |
| pCP20 | AmpR CmR, FLP synthesis | [33] |
| pCM17 | ori101 luxCDABE aph hok-sok par parA rrnB | [34] |
| Vaccine Strain | Immunization Schedule | |||||
|---|---|---|---|---|---|---|
| 1-Month Interval | 1-Week Interval | |||||
| Mortality | VE | p-Value * | Mortality | VE | p-Value * | |
| CVD 1921 | 6/28 | 79% | <0.0001 | 0/15 | 93% | <0.0001 |
| CVD 1921 ∆htrA | 14/30 | 53% | <0.0001 | 5/15 | 64% | 0.017 |
| CVD 1926 | 14/29 | 52% | <0.0001 | 5/15 | 64% | 0.017 |
| PBS | 30/30 | - | - | 14/15 | - | - |
| Vaccine Group | Mortality | VE | p-Value * |
|---|---|---|---|
| CVD 1921 | 3/15 | 80% | <0.0001 |
| CVD 1921 ∆htrA | 6/16 | 63% | 0.0002 |
| CVD 1926 | 11/16 | 31% | 0.0434 |
| PBS | 15/15 | - | - |
| Vaccine Group | Age and Immunization Schedule | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 14-Day-Old | 28-Day-Old | ||||||||
| 1-Week Interval | 2-Week Interval | 2-Week Interval | |||||||
| Mortality | VE | p-Value * | Mortality | VE | p-Value * | Mortality | VE | p-Value * | |
| CVD 1921 | 8/15 | 34% | 0.1351 | 3/17 | 79% | 0.0004 | 2/16 | 83% | 0.010 |
| CVD 1921 ∆htrA | 12/15 | 1.5% | 1.0000 | 10/17 | 29% | 0.2587 | 6/14 | 43% | 0.1349 |
| CVD 1926 | 9/15 | 26% | 0.2524 | 10/16 | 24% | 0.2587 | 5/16 | 58% | 0.0320 |
| PBS | 13/16 | - | 14/17 | - | 12/16 | - | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sears, K.T.; Nasrin, S.; Baliban, S.M.; Council, D.N.; Pasetti, M.F.; Tennant, S.M. Evaluation of Three Candidate Live-Attenuated Salmonella enterica Serovar Typhimurium Vaccines to Prevent Non-Typhoidal Salmonella Infection in an Infant Mouse Model. Vaccines 2023, 11, 1562. https://doi.org/10.3390/vaccines11101562
Sears KT, Nasrin S, Baliban SM, Council DN, Pasetti MF, Tennant SM. Evaluation of Three Candidate Live-Attenuated Salmonella enterica Serovar Typhimurium Vaccines to Prevent Non-Typhoidal Salmonella Infection in an Infant Mouse Model. Vaccines. 2023; 11(10):1562. https://doi.org/10.3390/vaccines11101562
Chicago/Turabian StyleSears, Khandra T., Shamima Nasrin, Scott M. Baliban, Danielle N. Council, Marcela F. Pasetti, and Sharon M. Tennant. 2023. "Evaluation of Three Candidate Live-Attenuated Salmonella enterica Serovar Typhimurium Vaccines to Prevent Non-Typhoidal Salmonella Infection in an Infant Mouse Model" Vaccines 11, no. 10: 1562. https://doi.org/10.3390/vaccines11101562
APA StyleSears, K. T., Nasrin, S., Baliban, S. M., Council, D. N., Pasetti, M. F., & Tennant, S. M. (2023). Evaluation of Three Candidate Live-Attenuated Salmonella enterica Serovar Typhimurium Vaccines to Prevent Non-Typhoidal Salmonella Infection in an Infant Mouse Model. Vaccines, 11(10), 1562. https://doi.org/10.3390/vaccines11101562