

Lack of Specific Immune Response after Five Doses of mRNA SARS-CoV-2 Vaccine in a Patient with CD4+ T-Cell Lymphopenia but Preserved Responses to CMV
 , ,
, ,
Abstract
1. Introduction
2. Case Report and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, P.; Anand, A.; Rana, S.; Kumar, A.; Goel, P.; Kumar, S.; Gouda, K.C.; Singh, H. Impact of COVID-19 vaccination: A global perspective. Front. Public Health 2023, 11, 1272961. [Google Scholar] [CrossRef] [PubMed]
- Watson, O.J.; Barnsley, G.; Toor, J.; Hogan, A.B.; Winskill, P.; Ghani, A.C. Global impact of the first year of COVID-19 vaccination: A mathematical modelling study. Lancet Infect. Dis. 2022, 22, 1293–1302. [Google Scholar] [CrossRef] [PubMed]
- Meyts, I.; Bucciol, G.; Quinti, I.; Neven, B.; Fischer, A.; Seoane, E.; Lopez-Granados, E.; Gianelli, C.; Robles-Marhuenda, A.; Jeandel, P.-Y.; et al. Coronavirus disease 2019 in patients with inborn errors of immunity: An international study. J. Allergy Clin. Immunol. 2021, 147, 520–531. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.E.; Mathew, S.; Peluso, M.J.; Cunningham-Rundles, C. Clinical outcomes and features of COVID-19 in patients with primary immunodeficiencies in New York City. J. Allergy Clin. Immunol. Pract. 2021, 9, 490–493.e2. [Google Scholar] [CrossRef] [PubMed]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef] [PubMed]
- Ministerio de Sanidad—Areas—Public Health—Prevention and Promotion—Vacunaciones—Estrategia de Vacunación COVID-19: Actualizaciones. Available online: https://www.sanidad.gob.es/en/areas/promocionPrevencion/vacunaciones/covid19/Actualizaciones_EstrategiaVacunacionCOVID-19.htm (accessed on 19 January 2024).
- Hagin, D.; Freund, T.; Navon, M.; Halperin, T.; Adir, D.; Marom, R.; Levi, I.; Benor, S.; Alcalay, Y.; Freund, N.T. Immunogenicity of Pfizer-BioNTech COVID-19 vaccine in patients with inborn errors of immunity. J. Allergy Clin. Immunol. 2021, 148, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Salinas, A.F.; Mortari, E.P.; Terreri, S.; Quintarelli, C.; Pulvirenti, F.; Di Cecca, S.; Guercio, M.; Milito, C.; Bonanni, L.; Auria, S.; et al. SARS-CoV-2 Vaccine Induced Atypical Immune Responses in Antibody Defects: Everybody Does their Best. J. Clin. Immunol. 2021, 41, 1709–1722. [Google Scholar] [CrossRef] [PubMed]
- Amodio, D.; Ruggiero, A.; Sgrulletti, M.; Pighi, C.; Cotugno, N.; Medri, C.; Morrocchi, E.; Colagrossi, L.; Russo, C.; Zaffina, S.; et al. Humoral and Cellular Response Following Vaccination With the BNT162b2 mRNA COVID-19 Vaccine in Patients Affected by Primary Immunodeficiencies. Front. Immunol. 2021, 12, 727850. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.M.; Guevara-Hoyer, K.; García, C.J.; Bravo, L.G.; Jiménez-Huete, A.; de la Peña, A.R.; Valeros, B.M.; Velázquez, C.C.; López, E.C.; Cabello, N.; et al. Specific Cellular and Humoral Immune Responses to the Neoantigen RBD of SARS-CoV-2 in Patients with Primary and Secondary Immunodeficiency and Healthy Donors. Biomedicines 2023, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Gómez, S.; Pérez de Diego, R.; García-Martínez, E.; Fernández-Cruz, E.; Gil-Herrera, J. Adulto con criterios clínicos de imunodeficiencia combinada no severa y acúmulo de variantes en genes implicados en la producción de anticuerpos y enfermedad inflamatoria intestinal. In Inmunología Clínica. Casos Clínicos en Inmunodeficiencias VI; Cámara, C., Fernández, L., Gil-Herrera, J., Eds.; Sociedad Española de Inmunología: Madrid, Spain, 2022; pp. 199–203. ISBN 978-84-09-43738-2. [Google Scholar]
- Gil-Manso, S.; Alonso, R.; Catalán, P.; Sánchez-Arcilla, I.; Marzola, M.; Correa-Rocha, R.; Pion, M.; Muñoz, P. IgG anti-RBD levels during 8-month follow-up post-vaccination with BNT162b2 and mRNA-1273 vaccines in healthcare workers: A one-center study. Front. Cell. Infect. Microbiol. 2022, 12, 1035155. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.D.; Wang, E.; Garrigan, E.; Goodwin, B.; Sutherland, A.; Tarke, A.; Chang, J.; Gálvez, R.I.; Mateus, J.; Ramirez, S.I.; et al. Development of a T cell-based immunodiagnostic system to effectively distinguish SARS-CoV-2 infection and COVID-19 vaccination status. Cell Host Microbe 2022, 30, 388–399.e3. [Google Scholar] [CrossRef] [PubMed]
- Christophorou, E.; Nilsson, A.C.; Petersen, I.; Lindvig, S.O.; Davidsen, J.R.; Abazi, R.; Poulsen, M.K.; Pedersen, R.M.; Justesen, U.S.; Johansen, N.E.; et al. Humoral antibody response following mRNA vaccines against SARS-CoV-2 in solid organ transplant recipients; a status after a fifth and bivalent vaccine dose. Front. Immunol. 2023, 14, 1270814. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Yang, X.; Lin, B.Y.; Li, Y.Z.; Huang, G.; Xu, Y. Immunological Findings in a Group of Individuals Who Were Poor or Non-Responders to Standard Two-Dose SARS-CoV-2 Vaccines. Vaccines 2023, 11, 461. [Google Scholar] [CrossRef]
- Granadier, D.; Iovino, L.; Kinsella, S.; Dudakov, J.A. Dynamics of thymus function and T cell receptor repertoire breadth in health and disease. Semin. Immunopathol. 2021, 43, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Gulla, S.; Reddy, M.C.; Reddy, V.C.; Chitta, S.; Bhanoori, M.; Lomada, D. Role of thymus in health and disease. Int. Rev. Immunol. 2023, 42, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Pothast, C.R.; Dijkland, R.C.; Thaler, M.; Hagedoorn, R.S.; Kester, M.G.D.; Wouters, A.K.; Hiemstra, P.S.; van Hemert, M.J.; Gras, S.; Falkenburg, J.H.F.; et al. SARS-CoV-2-specific CD4+ and CD8+ T cell responses can originate from cross-reactive CMV-specific T cells. eLife 2022, 11, e82050. [Google Scholar] [CrossRef] [PubMed]
- Tofighi Zavareh, F.; Mirshafiey, A.; Yazdani, R.; Keshtkar, A.A.; Abolhassani, H.; Mahdaviani, S.A.; Habibi, S.; Sohani, M.; Rezaei, N.; Aghamohammadi, A. Immunophenotypic and functional analysis of lymphocyte subsets in common variable immunodeficiency patients without monogenic defects. Scand. J. Immunol. 2022, 96, e13164. [Google Scholar] [CrossRef] [PubMed]
- Marashi, S.M.; Raeiszadeh, M.; Enright, V.; Tahami, F.; Workman, S.; Chee, R.; Webster, A.D.; Milne, R.S.B.; Emery, V.C. Influence of cytomegalovirus infection on immune cell phenotypes in patients with common variable immunodeficiency. J. Allergy Clin. Immunol. 2012, 129, 1349–1356.e3. [Google Scholar] [CrossRef] [PubMed]
- Attardi, E.; Di Cesare, S.; Amodio, D.; Giancotta, C.; Cotugno, N.; Cifaldi, C.; Chiriaco, M.; Palma, P.; Finocchi, A.; Di Matteo, G.; et al. Phenotypical T Cell Differentiation Analysis: A Diagnostic and Predictive Tool in the Study of Primary Immunodeficiencies. Front. Immunol. 2019, 10, 2735. [Google Scholar] [CrossRef] [PubMed]
- Sylwester, A.W.; Mitchell, B.L.; Edgar, J.B.; Taormina, C.; Pelte, C.; Ruchti, F.; Sleath, P.R.; Grabstein, K.H.; Hosken, N.A.; Kern, F.; et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 2005, 202, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Pardieck, I.N.; Beyrend, G.; Redeker, A.; Arens, R. Cytomegalovirus infection and progressive differentiation of effector-memory T cells. F1000Research 2018, 7, F1000 Faculty Rev-1554. [Google Scholar] [CrossRef]
- Zhang, W.; Morris, A.B.; Peek, E.V.; Karadkhele, G.; Robertson, J.M.; Kissick, H.T.; Larsen, C.P. CMV Status Drives Distinct Trajectories of CD4+ T Cell Differentiation. Front. Immunol. 2021, 12, 620386. [Google Scholar] [CrossRef] [PubMed]
- Cicin-Sain, L. Cytomegalovirus memory inflation and immune protection. Med. Microbiol. Immunol. 2019, 208, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Chidrawar, S.; Khan, N.; Wei, W.; McLarnon, A.; Smith, N.; Nayak, L.; Moss, P. Cytomegalovirus-seropositivity has a profound influence on the magnitude of major lymphoid subsets within healthy individuals. Clin. Exp. Immunol. 2009, 155, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Pourgheysari, B.; Khan, N.; Best, D.; Bruton, R.; Nayak, L.; Moss, P.A.H. The cytomegalovirus-specific CD4+ T-cell response expands with age and markedly alters the CD4+ T-cell repertoire. J. Virol. 2007, 81, 7759–7765. [Google Scholar] [CrossRef] [PubMed]
- Almyroudis, N.G.; Jakubowski, A.; Jaffe, D.; Sepkowitz, K.; Pamer, E.; O’Reilly, R.J.; Papanicolaou, G.A. Predictors for persistent cytomegalovirus reactivation after T-cell-depleted allogeneic hematopoietic stem cell transplantation. Transpl. Infect. Dis. 2007, 9, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Gabanti, E.; Lilleri, D.; Ripamonti, F.; Bruno, F.; Zelini, P.; Furione, M.; Colombo, A.A.; Alessandrino, E.P.; Gerna, G. Reconstitution of Human Cytomegalovirus-Specific CD4+ T Cells is Critical for Control of Virus Reactivation in Hematopoietic Stem Cell Transplant Recipients but Does Not Prevent Organ Infection. Biol. Blood Marrow Transpl. J. Am. Soc. Blood Marrow Transplant. 2015, 21, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, N.; Isozaki, T.; Shirakawa, K.; Ikegaya, N.; Yamamoto, T.; Hishida, A. Cytomegalovirus colitis following immunosuppressive therapy for lupus peritonitis and lupus nephritis. Intern. Med. Tokyo Jpn. 2003, 42, 362–366. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nakase, H.; Herfarth, H. Cytomegalovirus Colitis, Cytomegalovirus Hepatitis and Systemic Cytomegalovirus Infection: Common Features and Differences. Inflamm. Intest. Dis. 2016, 1, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Chanouzas, D.; Sagmeister, M.; Faustini, S.; Nightingale, P.; Richter, A.; Ferro, C.J.; Morgan, M.D.; Moss, P.; Harper, L. Subclinical Reactivation of Cytomegalovirus Drives CD4+CD28null T-Cell Expansion and Impaired Immune Response to Pneumococcal Vaccination in Antineutrophil Cytoplasmic Antibody-Associated Vasculitis. J. Infect. Dis. 2019, 219, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Cantenys-Molina, S.; Fernández-Cruz, E.; Francos, P.; Lopez Bernaldo de Quirós, J.C.; Muñoz, P.; Gil-Herrera, J. Lymphocyte subsets early predict mortality in a large series of hospitalized COVID-19 patients in Spain. Clin. Exp. Immunol. 2021, 203, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Maple, P.A.C. COVID-19, SARS-CoV-2 Vaccination, and Human Herpesviruses Infections. Vaccines 2023, 11, 232. [Google Scholar] [CrossRef] [PubMed]



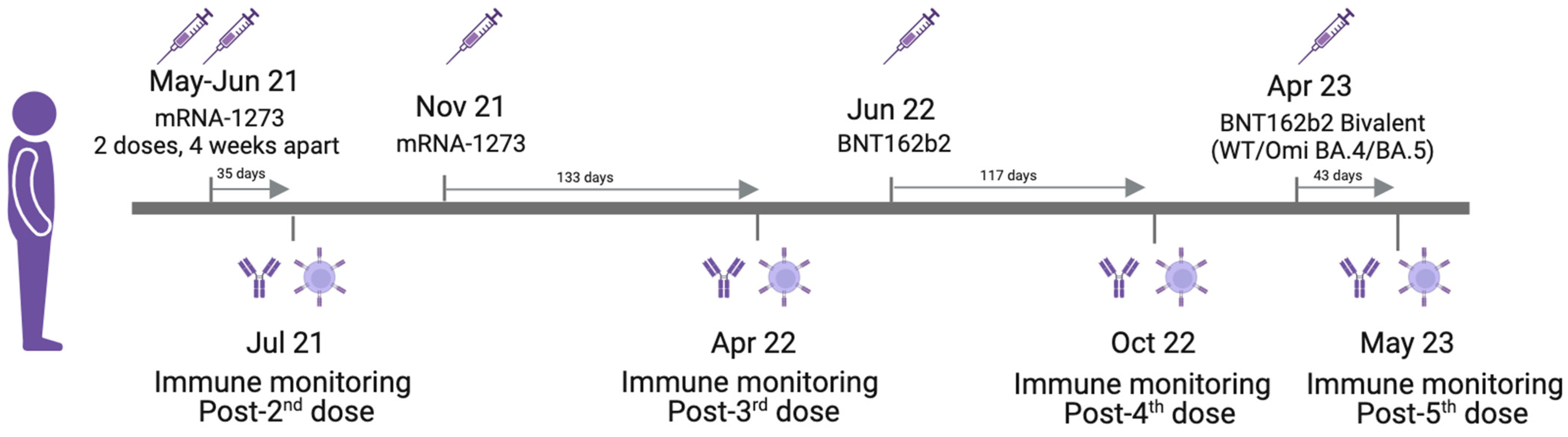{kind=link}
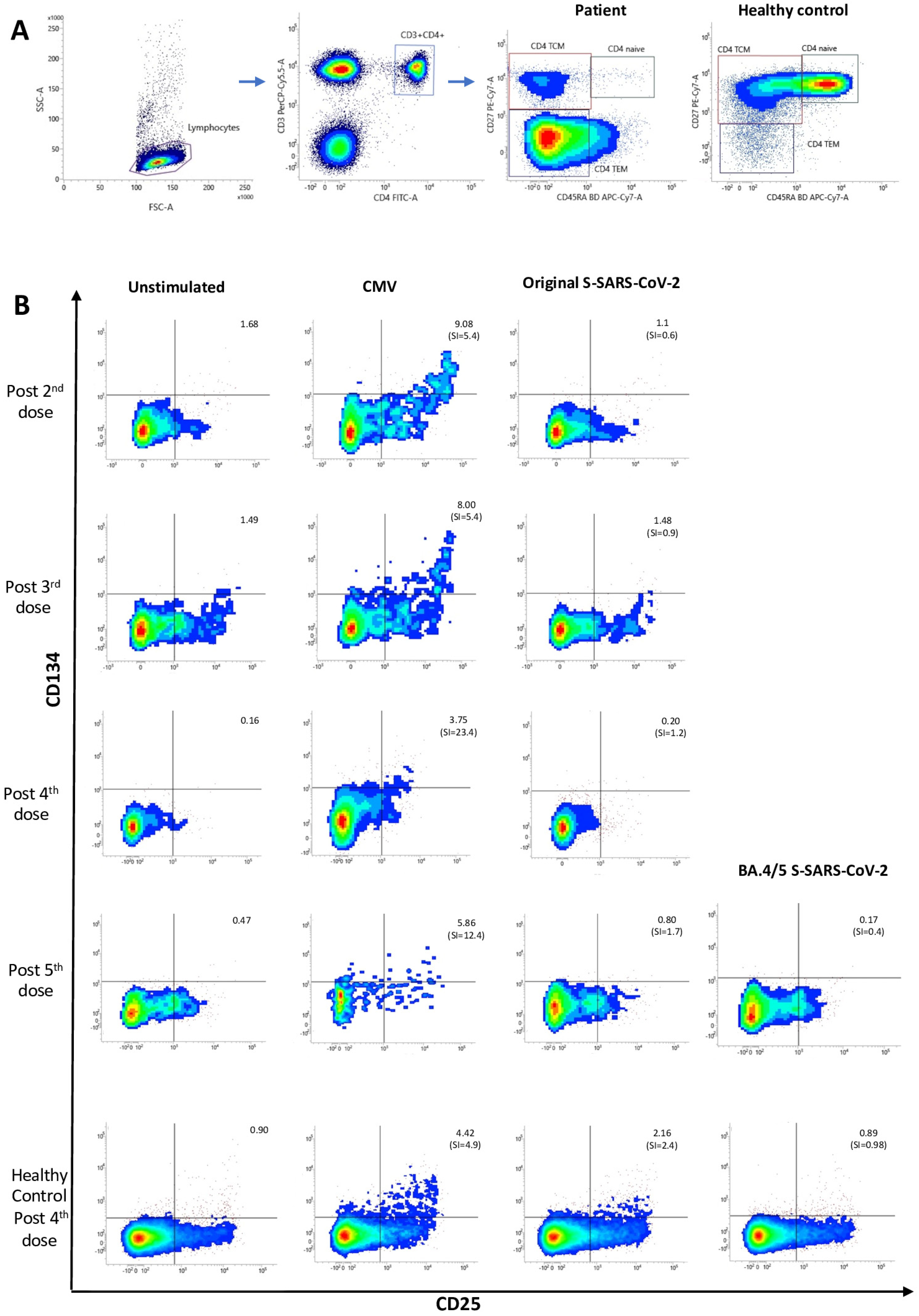{kind=link}
| Laboratory Parameter | Result | Reference Range | ||
|---|---|---|---|---|
| Immunoglobulins | ||||
| IgG (mg/dL) | 1460 | 670–1650 | ||
| IgA (mg/dL) | 104 | 98–543 | ||
| IgM (mg/dL) | 139 | 49–302 | ||
| IgG subclasses | ||||
| IgG1 (g/L) | 7.48 | 2.78–8.22 | ||
| IgG2 (g/L) | 6.28 | 1.47–6.29 | ||
| IgG3 (g/L) | 0.62 | 0.15–1.3 | ||
| IgG4 (g/L) | 0.41 | 0.032–1.315 | ||
| Antigen-specific responses | ||||
| Anti-pneumococcal polysaccharide IgG antibodies (mg/dL) | Pre-vaccination 1.1 Post-vaccination > 27 | 1–19.1 | ||
| Anti-cytomegalovirus (CMV) IgG antibodies (AU/mL) | Positive | ≥6.0 | ||
| Autoimmunity | ||||
| Anti-nuclear autoantibodies | Positive 1/640 | <1/80 | ||
| Anti-dsDNA autoantibodies (IU/mL) | Positive 39 | 0–15 | ||
| Anti-histone autoantibodies (relative intensity (%)) | Positive 30.9 | ≥15.9 | ||
| Anti-cardiolipin IgG autoantibodies (GPL U/mL) | Positive 13.7 | 0–10 | ||
| Lymphocyte subpopulations | ||||
| % T cells CD3+ (cells/μL) | 69 (396) | 55–82 (700–2100) | ||
| % helper T cells CD4+ (cells/μL) | 25 (148) | 28–57 (300–1400) | ||
| % cytotoxic T cells CD8+ (cells/μL) | 37 (218) | 10–39 (200–1200) | ||
| CD4/CD8 | 0.68 | 1.00–3.60 | ||
| % TCR γ/δ+ T cells | 18 | 0.8–11.00 | ||
| % CD4+CD45RA+CD31+ cells (recent thymic emigrants) | 3 | 14.4–38.3 | ||
| % CD3+CD45RA+CD45RO− (naïve T cells) | 12 | 17.6–39.6 | ||
| % B cells CD19+ (cells/μL) | 13 (75) | 6–19 (100–500) | ||
| % CD19+IgD+CD27− cells (naïve B cells) | 59 | 28–79 | ||
| % CD19+IgD+CD27+ cells (memory IgM B cells) | 28 | 8–37 | ||
| % CD19+IgD−CD27+ cells (switched memory B cells) | 10 | 5–35 | ||
| % NK cells CD56+ (cells/μL) | 18 (101) | 7–31 (90–600) | ||
| Immunogenetics | ||||
| TCR rearrangements | Oligoclonal | |||
| Gene | Dbsnp | Genomic Location | Protein Change | Associated Phenotype |
| IRF2BP2 | rs763707638 | g.1:234608652: C > G | p.Glu281Asp | Common variable immunodeficiency (CVID) |
| TNFRF13C | rs77874543 | g.22:41926712: G > C | p.Pro21Arg | |
| rs61756766 | g.22:41925447: G > A | p.His159Tyr | ||
| BTNL2 | rs115653647 | g.6:32403216 C > T | p.Gly143Asp | Crohn’s disease susceptibility |
| rs28362680 | g.6:32403039 G > A | p.Ala202Val | ||
| Vaccine | Anti-SARS-CoV-2 IgG Antibodies (BAU/mL) | Anti-CMV IgG Antibodies (AU/mL) |
|---|---|---|
| 1st and 2nd doses mRNA-1273 (Moderna) | 0 | 163 |
| 3rd dose mRNA-1273 (Moderna) | 9.4 | 1026 |
| 4th dose BNT162b2 (Pfizer/BioNTech) | 17.8 | 606 |
| 5th dose BNT162b2 bivalent (Pfizer/BioNTech) | 76.7 | 562 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alba-Cano, T.; Fernández-Cruz, E.; Alonso, R.; Muñoz-Gómez, S.; Pérez de Diego, R.; García Martínez, E.; Sánchez-Mateos, P.; Navarro Caspistegui, J.; Martín López, M.; Gil-Herrera, J. Lack of Specific Immune Response after Five Doses of mRNA SARS-CoV-2 Vaccine in a Patient with CD4+ T-Cell Lymphopenia but Preserved Responses to CMV. Vaccines 2024, 12, 386. https://doi.org/10.3390/vaccines12040386
Alba-Cano T, Fernández-Cruz E, Alonso R, Muñoz-Gómez S, Pérez de Diego R, García Martínez E, Sánchez-Mateos P, Navarro Caspistegui J, Martín López M, Gil-Herrera J. Lack of Specific Immune Response after Five Doses of mRNA SARS-CoV-2 Vaccine in a Patient with CD4+ T-Cell Lymphopenia but Preserved Responses to CMV. Vaccines. 2024; 12(4):386. https://doi.org/10.3390/vaccines12040386
Chicago/Turabian StyleAlba-Cano, Trinidad, Eduardo Fernández-Cruz, Roberto Alonso, Sara Muñoz-Gómez, Rebeca Pérez de Diego, Elena García Martínez, Paloma Sánchez-Mateos, Joaquín Navarro Caspistegui, Mónica Martín López, and Juana Gil-Herrera. 2024. "Lack of Specific Immune Response after Five Doses of mRNA SARS-CoV-2 Vaccine in a Patient with CD4+ T-Cell Lymphopenia but Preserved Responses to CMV" Vaccines 12, no. 4: 386. https://doi.org/10.3390/vaccines12040386
APA StyleAlba-Cano, T., Fernández-Cruz, E., Alonso, R., Muñoz-Gómez, S., Pérez de Diego, R., García Martínez, E., Sánchez-Mateos, P., Navarro Caspistegui, J., Martín López, M., & Gil-Herrera, J. (2024). Lack of Specific Immune Response after Five Doses of mRNA SARS-CoV-2 Vaccine in a Patient with CD4+ T-Cell Lymphopenia but Preserved Responses to CMV. Vaccines, 12(4), 386. https://doi.org/10.3390/vaccines12040386