
The Trimeric Autotransporter Adhesin SadA from Salmonella spp. as a Novel Bacterial Surface Display System
Abstract
:1. Introduction
2. Materials and Methods
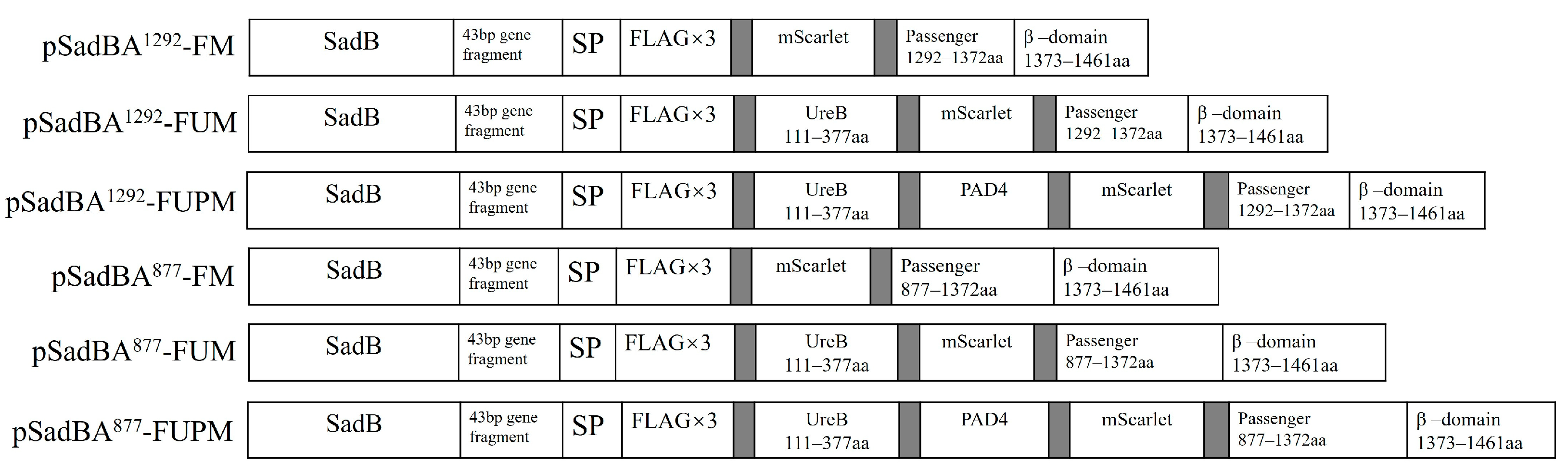
2.1. Plasmid Construction
2.2. Chromosomal Deletion of sadA, ygeA, and murI Genes from S. typhimurium
2.3. Strains and Growth Conditions
2.4. Protein Expression and Analysis
2.5. Proteinase K Treatment of Bacterial Cells
2.6. Immunofluorescence and Flow Cytometric Analyses
2.7. Whole-Cell Dot Blot
2.8. Immunization, Sample Collection, and Specific Antibody Detection by ELISA
2.9. Statistical Analysis
3. Results
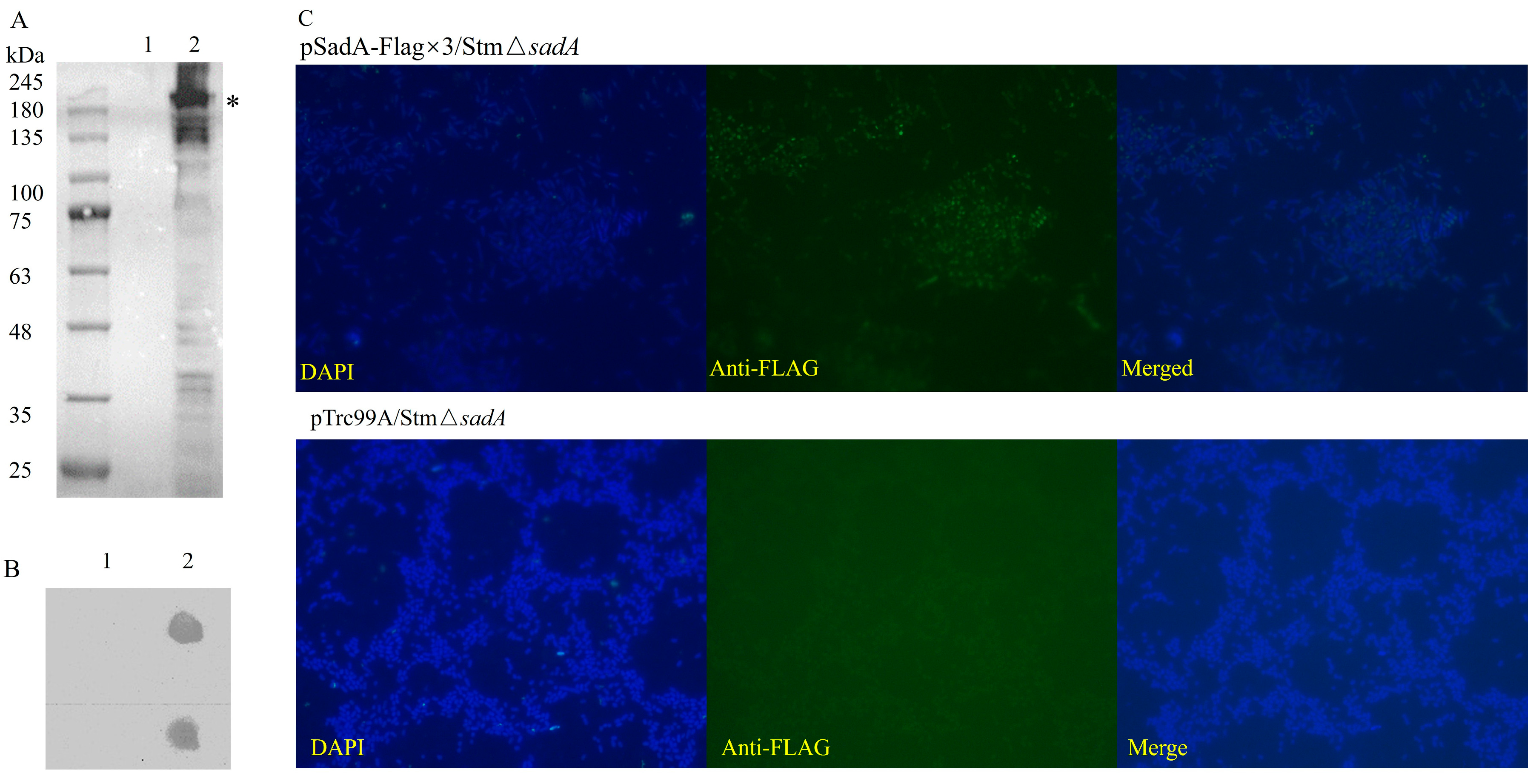
3.1. Full-Length SadA Can Be Displayed on the Surface of S. typhimurium
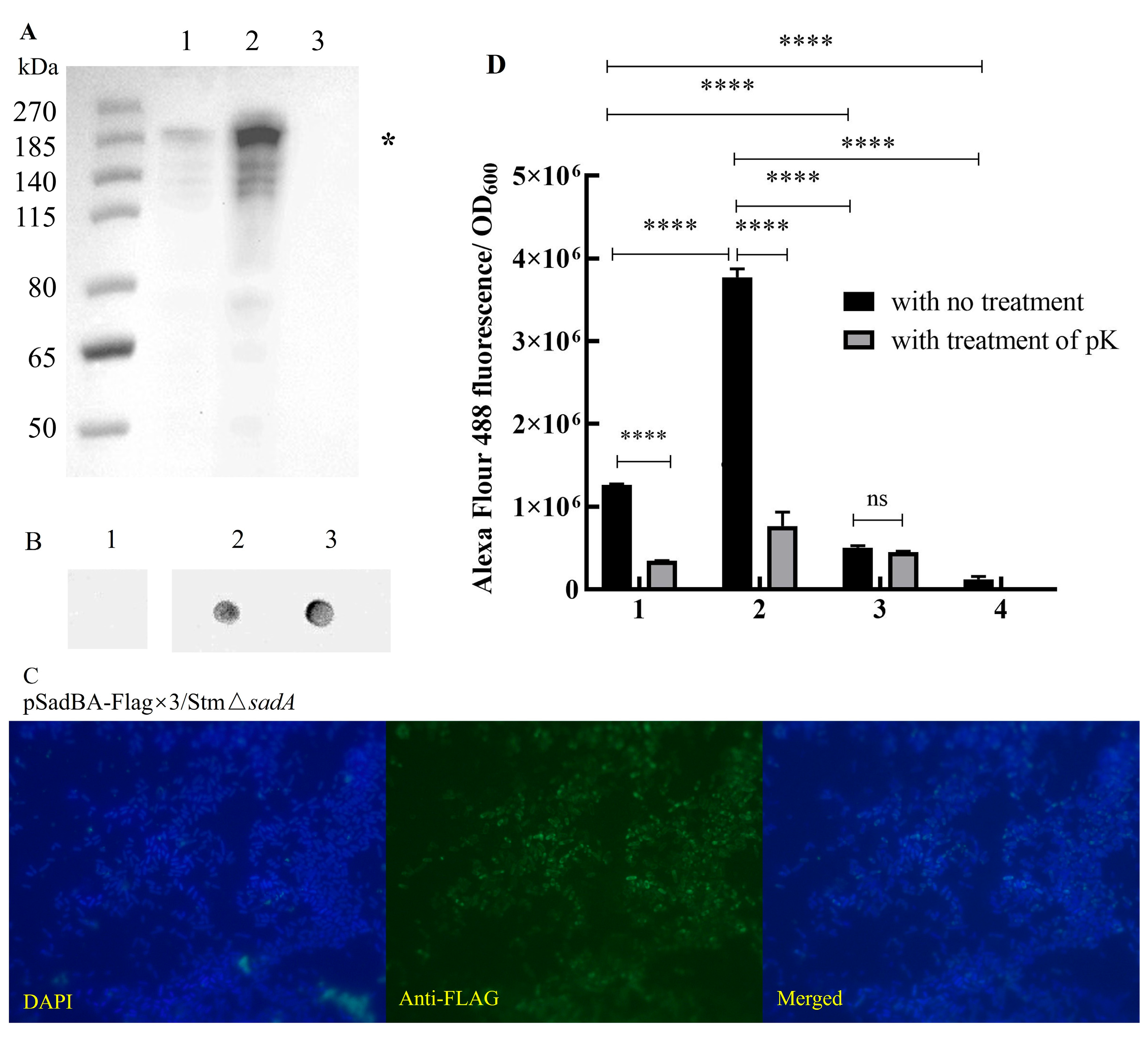
3.2. SadB Enhances the Surface Display of SadA in S. typhimurium
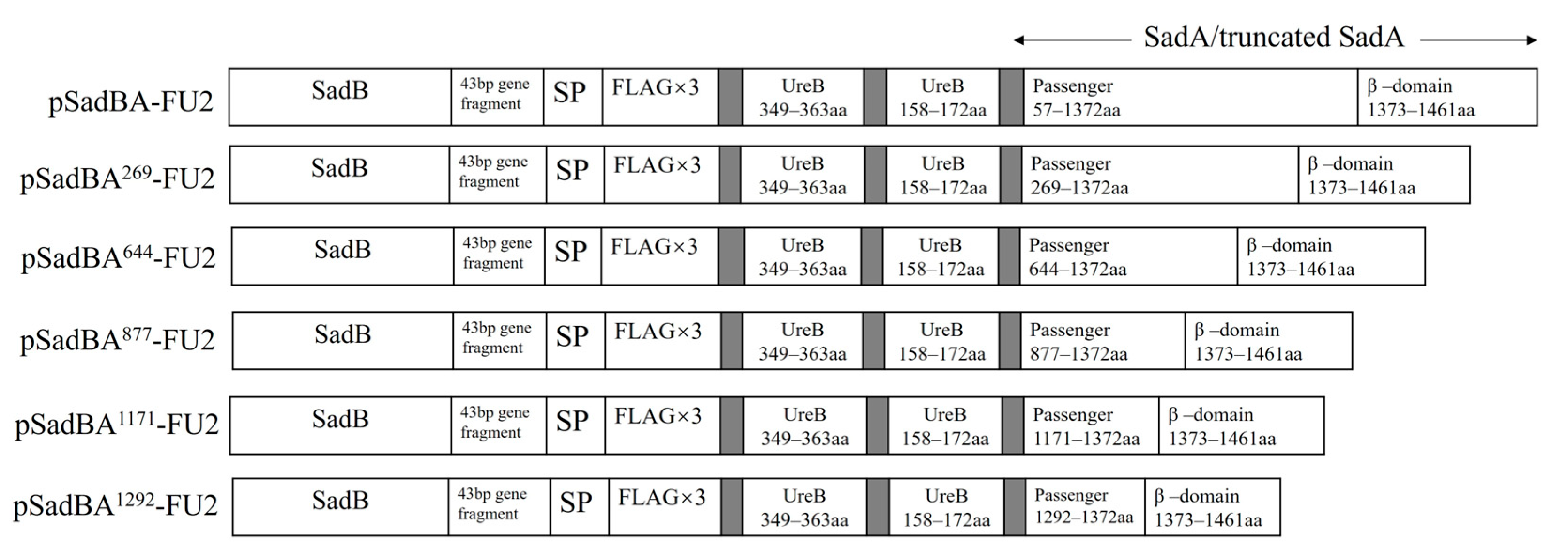
3.3. Epitopes Can Be Displayed on the Cell Surface Using SadAs with Different Sizes as Anchoring Motifs
3.4. Truncated SadAs Can Mediate the Surface Display of Heterologous Proteins on the S. typhimurium Mutant
3.5. Immune Responses Elicited by pSadBA1292-FUM/StmΔygeAΔmurI and pSadBA877-FUM/StmΔygeAΔmurI in Mice Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sevastsyanovich, Y.R.; Withers, D.R.; Marriott, C.L.; Morris, F.C.; Wells, T.J.; Browning, D.F.; Beriotto, I.; Ross, E.; Ali, H.O.; Wardius, C.A.; et al. Antigen localization influences the magnitude and kinetics of endogenous adaptive immune response to recombinant Salmonella vaccines. Infect. Immun. 2017, 85, e00593-17. [Google Scholar] [CrossRef] [PubMed]
- Wiull, K.; Boysen, P.; Kuczkowska, K.; Moen, L.F.; Carlsen, H.; Eijsink, V.G.H.; Mathiesen, G. Comparison of the immunogenic properties of Lactiplantibacillus plantarum carrying the Mycobacterial Ag85B-ESAT-6 antigen at various cellular localizations. Front. Microbiol. 2022, 13, 900922. [Google Scholar] [CrossRef] [PubMed]
- Chmielewski, M.; Kuehle, J.; Chrobok, D.; Riet, N.; Hallek, M.; Abken, H. FimH-based display of functional eukaryotic proteins on bacteria surfaces. Sci. Rep. 2019, 9, 8410. [Google Scholar] [CrossRef] [PubMed]
- Hui, C.Y.; Guo, Y.; Liu, L.; Zheng, H.Q.; Wu, H.M.; Zhang, L.Z.; Zhang, W. Development of a novel bacterial surface display system using truncated OmpT as an anchoring motif. Biotechnol. Lett. 2019, 41, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Han, M.J. Novel bacterial surface display system based on the Escherichia coli protein MipA. J. Microbiol. Biotechnol. 2020, 30, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Zhou, W.; Ding, S.; Lu, Y.; Liu, S.; Cao, J.; Zhang, Z.; Liu, S.; Yu, S. An Escherichia coli carrier vaccine with surface-displayed protein MAP3061c elicits protective immunity against Mycobacterium paratuberculosis in mice. Res. Vet. Sci. 2021, 141, 180–189. [Google Scholar] [CrossRef]
- Vargas, L.M.; Muñoz-López, P.; Becerra-Báez, E.I.; Flores-Martínez, L.F.; Prada-Gracia, D.; Moreno-Vargas, L.M.; Baay-Guzmán, G.J.; Juárez-Hernández, U.; Chávez-Munguía, B.; Cabrera-Muñóz, L.; et al. Live attenuated Salmonella enterica expressing and releasing cell-permeable Bax BH3 peptide through the MisL autotransporter system elicits antitumor activity in a murine xenograft model of human B non-hodgkin’s lymphoma. Front. Immunol. 2019, 10, 2562. [Google Scholar]
- Gustavsson, M.; Do, T.H.; Lüthje, P.; Tran, N.T.; Brauner, A.; Samuelson, P.; Truong, N.H.; Larsson, G. Improved cell surface display of Salmonella enterica serovar enteritidis antigens in Escherichia coli. Microb. Cell Fact. 2015, 14, 47. [Google Scholar] [CrossRef]
- Pardavé-Alejandre, H.D.; Alvarado-Yaah, J.E.; Pompa-Mera, E.N.; Muñoz-Medina, J.E.; Sárquiz-Martínez, B.; Santacruz-Tinoco, C.E.; Manning-Cela, R.G.; Ortíz-Navarrete, V.; López-Macías, C.; González-Bonilla, C.R. Autodisplay of an avidin with biotin-binding activity on the surface of Escherichia coli. Biotechnol. Lett. 2018, 40, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg van Saparoea, H.B.; Houben, D.; de Jonge, M.I.; Jong, W.S.P.; Luirink, J. Display of recombinant proteins on bacterial outer membrane vesicles by using protein ligation. Appl. Environ. Microbiol. 2018, 84, e02567-17. [Google Scholar] [CrossRef] [PubMed]
- Thibau, A.; Dichter, A.A.; Vaca, D.J.; Linke, D.; Goldman, A.; Kempf, V.A.J. Immunogenicity of trimeric autotransporter adhesins and their potential as vaccine targets. Med. Microbiol. Immunol. 2020, 209, 243–263. [Google Scholar] [CrossRef]
- Hoiczyk, E.; Roggenkamp, A.; Reichenbecher, M.; Lupas, A.; Heesemann, J. Structure and sequence ananlysis of Yersinia YadA and Moraxella UspAs reveal a novel class of adhesins. EMBO J. 2000, 19, 5989–5999. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.F.; Kaiser, P.O.; Linke, D.; Schwarz, H.; Riess, T.; Schäfer, A.; Eble, J.A.; Kempf, V.A. Trimeric autotransporter adhesin-dependent adherence of Baronella henselae, Bartonella quintana, and Yersinia enterocolitica to matrix components and endothelial cells under static and dynamic flow conditions. Infect. Immun. 2011, 79, 2544–2553. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, H.; Kanie, J.; Hori, K. On-fiber display of a functional peptide at sites distant from the cell surface using a long bacterionanofiber of a trimeric autotransporter adhesin. Biotchnol. Bioeng. 2019, 116, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.H.; Kuijl, C.; Huynh, D.T.; Jong, W.S.P.; Luirink, J.; van Ulsen, P. Overproducing the BAM complex improves secretion of difficult-to-secrete recombinant autotranporter chimeras. Microb. Cell Factories 2021, 20, 176. [Google Scholar] [CrossRef]
- Chin’ombe, N. Recombinant Salmonella enterica serovar Typhimurium as a vaccine vector for HIV-1 Gag. Viruses 2013, 5, 2062–2078. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, S.I.; Frank, F.M.; Becker, P.D.; Arnaiz, M.; Mirkin, G.A.; Corral, R.S.; Guzmán, C.A.; Malchiodi, E.L. Redirection of the immune response to the functional catalytic domain of the cysteine proteinase cruzipain improves protective immunity against Trypanosoma cruzi infection. J. Infect. Dis. 2010, 202, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Ramani, K.; Cormack, T.; Cartwright, A.N.R.; Alami, A.; Parameswaran, P.; Abdou, M.; Wang, I.; Hilliard-Barth, K.; Argueta, S.; Raghunathan, D.; et al. SadA, a trimeric autotransporter from Salmonella enterica serovar Typhimurium, can promote biofilm formation and provides limited protection against infection. Infect. Immun. 2011, 79, 4342–4352. [Google Scholar]
- Hartmann, M.D.; Grin, I.; Dunin-Horkawicz, S.; Deiss, S.; Linke, D.; Lupas, A.N.; Alvarez, B.H. Complete fiber structures of complex trimeric autotransporter adhesins conserved in enterobacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 20907–20912. [Google Scholar] [CrossRef] [PubMed]
- Grin, I.; Hartmann, M.D.; Sauer, G.; Hernandez Alvarez, B.; Schütz, M.; Wagner, S.; Madlung, J.; Macek, B.; Felipe-Lopez, A.; Hensel, M.; et al. A trimeric lipoprotein assists in trimeric autotransporter biogenesis in enterobacteria. J. Biol. Chem. 2014, 289, 7388–7398. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef]
- Zhang, X.; Sang, S.; Guan, Q.; Tao, H.; Wang, Y.; Liu, C. Oral administration of a Shigella 2aT32-based vaccine expressing UreB-HspA fusion antigen with and without parenteral rUreB-HspA boost confers protection against Helicobacter pylori in mice model. Front. Immunol. 2022, 13, 894206. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wang, Y.C.; Tao, H.X.; Zhan, D.W.; Yuan, S.L.; Wang, P.; Wang, L.C.; Han, X.P.; Li, C.S.; Li, J.K.; et al. Identification of B-cell epitopes in urease B subunit of Helicobacter pylori bound by neutralizing antibodies. Vaccine 2010, 28, 5220–5227. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.; Hatlem, D.; Orwick-Rydmark, M.; Schneider, K.; Floetenmeyer, M.; van Rossum, B.; Leo, J.C.; Linke, D. Insights into the autotransport process of a trimeric autotranporter, Yersinia adhesin A (YadA). Mol. Microbiol. 2019, 111, 844–862. [Google Scholar] [CrossRef] [PubMed]
- Sikdar, R.; Bernstein, H.D. Sequential translocation of polypeptides across the bacterial outer membrane through the trimeric autotransporter pathway. mBio 2019, 10, e01973-19. [Google Scholar] [CrossRef] [PubMed]
- Cotter, S.E.; Surana, N.K.; Grass, S.; St Geme, J.W. 3rd. Trimeric autotransporters require trimerization of the passenger domain for stability and adhesive activity. J. Bacteriol. 2006, 188, 5400–5407. [Google Scholar] [CrossRef] [PubMed]
- Zakowska, D.; Graniak, G.; Rutyna, P.; Naylor, K.; Glowacka, P.; Niemcewicz, M. Protective antigen domain 4 of Bacillus anthracis as a candidate for use as vaccine for anthrax. Ann. Agric. Environ. Med. 2019, 26, 392–395. [Google Scholar] [CrossRef]
- Kiessling, A.R.; Malik, A.; Goldman, A. Recent advances in the understanding of trimeric autotransporter adhesins. Med. Microbiol. Immunol. 2020, 209, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Qu, J.; Mayer, C.; Behrens, S.; Holst, O.; Kleinschmidt, J.H. The trimeric periplasmic chaperone Skp of Escherichia coli forms 1:1 complexes with outer membrane proteins via hydrophobic and electrostatic interactions. J. Mol. Biol. 2007, 374, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Kudva, R.; Denks, K.; Kuhn, P.; Vogt, A.; Müller, M.; Koch, H.G. Protein translocation across the inner membrane of gram-negative bacteria: The Sec and Tat dependent protein transport pathways. Res. Microbiol. 2013, 164, 505–534. [Google Scholar] [CrossRef]
- Meuskens, I.; Michalik, M.; Chauhan, N.; Linke, D.; Leo, J.C. A new strain collection for improved expression of outer membrane proteins. Front. Cell Infect. Microbiol. 2017, 7, 464. [Google Scholar] [CrossRef] [PubMed]
- Veiga, E.; De Lorenzo, V.; Fernandez, L.A. Structural tolerance of bacterial autotransporters for folded passenger protein domains. Mol. Microbiol. 2004, 52, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Ströhle, F.W.; Kranen, E.; Schrader, J.; Maas, R.; Holtmann, D. A simplified process design for P450 driven hydroxylation based on surface displayed enzymes. Biotechnol. Bioeng. 2016, 113, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Olvera, P.; Ruiz-Pérez, F.; Sepulveda, N.V.; Santiago-Machuca, A.; Maldonado-Rodríguez, R.; Garcia-Elorriaga, G.; González-Bonilla, C. Display and release of the Plasmodium falciparum circumsporozoite protein using the autotransporter MisL of Salmonella enterica. Plasmid 2003, 50, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yang, X.; Wang, D.; Yu, J.; Wang, Y. Surface Display of the HPV L1 Capsid Protein by the Autotransporter Shigella IcsA. J. Microbiol. 2014, 52, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Jong, W.S.; Daleke-Schermerhorn, M.H.; Vikstrom, D.; Ten Hagen-Jongman, C.M.; de Punder, K.; van der Wel, N.N.; van de Sandt, C.E.; Rimmelzwaan, G.F.; Follmann, F.; Agger, E.M.; et al. An autotransporter display platform for the development of multivalent recombinant bacterial vector vaccines. Microb. Cell Factories 2014, 13, 162. [Google Scholar] [CrossRef] [PubMed]
- Saurí, A.; Ten Hagen-Jongman, C.M.; van Ulsen, P.; Luirink, J. Estimating the size of the active translocation pore of an autotransporter. J. Mol. Biol. 2012, 416, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Jong, W.S.P.; ten Hagen-Jongman, C.M.; den Blaauwen, T.; Slotboom, D.J.; Tame, J.R.; Wickström, D.; de Gier, J.W.; Otto, B.R.; Luirink, J. Limited tolerance towards folded elements during secretion of the autotransporter Hbp. Mol. Microbiol. 2007, 63, 1524–1536. [Google Scholar] [CrossRef] [PubMed]
- Rooke, J.L.; Icke, C.; Wells, T.J.; Rossiter, A.E.; Browning, D.F.; Morris, F.C.; Leo, J.C.; Schütz, M.S.; Autenrieth, I.B.; Cunningham, A.F.; et al. BamA and Bam D are essential for the secretion of trimeric autotransporter adhesins. Front. Microbiol. 2021, 12, 628879. [Google Scholar] [CrossRef]
- Ding, C.; Ma, J.; Dong, Q.; Liu, Q. Live bacterial vaccine vector and delivery strategies of heterologous antigen: A review. Immunol. Lett. 2018, 197, 70–77. [Google Scholar] [CrossRef]
- Pantophlet, R.; Burton, D.R. Immuofocusing: Antigen engineering to promote the induction of HIV-neutralizing antibodies. Trends Mol. Med. 2003, 9, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Kong, Q.; Curtiss, R. 3rd. New technologies in developing recombinant attenuated Salmonella vaccine vectors. Microb. Pathog. 2013, 58, 17–28. [Google Scholar] [CrossRef] [PubMed]
- García, P.; Moscoso, M.; Fuentes-Valverde, V.; Rodicio, M.R.; Herrera-León, S.; Bou, G. A highly-safe live auxotrophic vaccine protecting against disease caused by non-typhoidal Salmonella Typhimurium in mice. J. Microbiol. Immunol. Infect. 2023, 56, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Cabral, M.P.; Correia, A.; Vilanova, M.; Gärtner, F.; Moscoso, M.; García, P.; Vallejo, J.A.; Pérez, A.; Francisco-Tomé, M.; Fuentes-Valverde, V.; et al. A live auxotrophic vaccine confers mucosal immunity and protection against lethal pneumonia caused by Pseudomonas aeruginosa. PLoS Pathog. 2020, 16, e1008311. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Positive Rate (%) | Average Value | ||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| pTrc99A/StmΔsadA | 0.02% | 0.00% | 0.00% | 0.0067% |
| pSadBA1292-FU2/StmΔsadA | 71.55% | 77.22% | 73.85% | 74.21% |
| pSadBA1171-FU2/StmΔsadA | 41.17% | 32.41% | 41.13% | 38.24% |
| pSadBA877-FU2/StmΔsadA | 78.51% | 83.99% | 82.55% | 81.68% |
| pSadBA644-FU2/StmΔsadA | 50.39% | 52.05% | 55.16% | 52.53% |
| pSadBA269-FU2/StmΔsadA | 85.70% | 86.50% | 87.90% | 86.70% |
| pSadBA-FU2/StmΔsadA | 85.00% | 87.37% | 89.08% | 87.15% |
| pSadBA1292-FM/StmΔsadA | 85.77% | 83.46% | 84.43% | 84.55% |
| pSadBA1292-FUM/StmΔsadA | 76.30% | 75.26% | 74.08% | 75.21% |
| pSadBA1292-FUPM/StmΔsadA | 82.55% | 82.75% | 85.26% | 83.52% |
| pSadBA877-FM/StmΔsadA | 92.47% | 93.36% | 93.43% | 93.09% |
| pSadBA877-FUM/StmΔsadA | 32.04% | 33.12% | 41.23% | 35.46% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sang, S.; Song, W.; Lu, L.; Ou, Q.; Guan, Y.; Tao, H.; Wang, Y.; Liu, C. The Trimeric Autotransporter Adhesin SadA from Salmonella spp. as a Novel Bacterial Surface Display System. Vaccines 2024, 12, 399. https://doi.org/10.3390/vaccines12040399
Sang S, Song W, Lu L, Ou Q, Guan Y, Tao H, Wang Y, Liu C. The Trimeric Autotransporter Adhesin SadA from Salmonella spp. as a Novel Bacterial Surface Display System. Vaccines. 2024; 12(4):399. https://doi.org/10.3390/vaccines12040399
Chicago/Turabian StyleSang, Shuli, Wenge Song, Lu Lu, Qikun Ou, Yiyan Guan, Haoxia Tao, Yanchun Wang, and Chunjie Liu. 2024. "The Trimeric Autotransporter Adhesin SadA from Salmonella spp. as a Novel Bacterial Surface Display System" Vaccines 12, no. 4: 399. https://doi.org/10.3390/vaccines12040399