
Advances in Nucleic Acid Universal Influenza Vaccines

Abstract
1. Introduction
2. A Brief Description of Influenza Viruses
2.1. Influenza Epidemiology
2.2. Influenza Virus Typology and Structural Characteristics
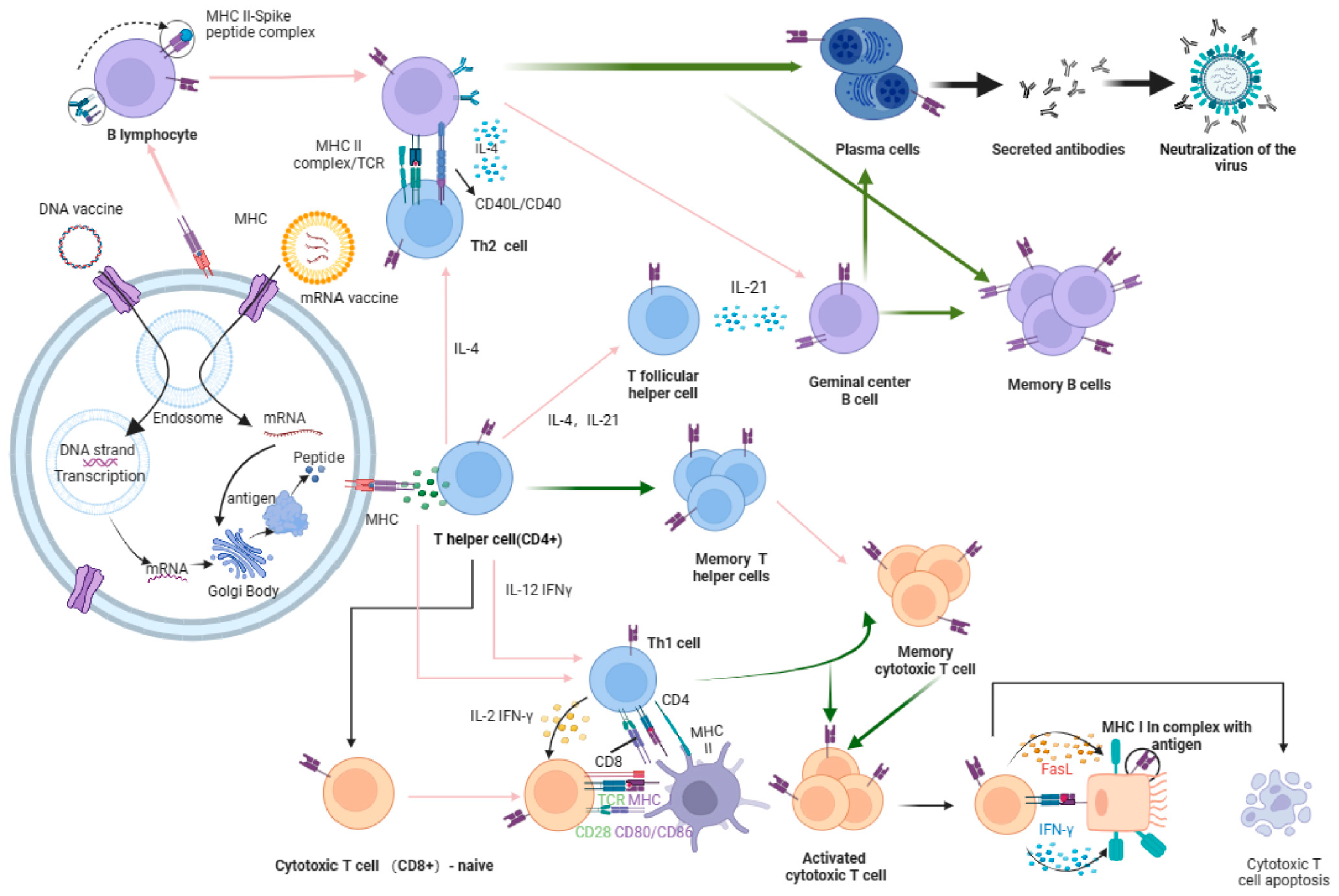
3. Nucleic Acid Vaccines
3.1. Advantages and Disadvantages of Nucleic Acid Vaccines
3.2. Universal Influenza DNA Vaccines
3.3. Development of Universal Influenza mRNA Vaccines

{kind=link}
| Authors | Published Year | Antigens | Animal Model | Clinical Trial |
|---|---|---|---|---|
| Freyn et al. [52] | 2020 | HA stalk, NA, M2, NP, | Mice | |
| Arevalo et al. [55] | 2022 | HA from 20 subtypes | Mice | |
| McMahon et al. [53] | 2022 | HA stalk, NA, M2, NP | Mice | |
| Ven et al. [56] | 2022 | NP, M1, PB1 | Ferret | |
| Zhu et al. [57] | 2022 | HA, MI, NP | Mice | |
| Pardi et al. [54] | 2022 | B/Yamagata/16/1988-like lineage HA B/Victoria/2/1987-like lineage HA, NA, NP, and M2 | Mice | |
| Widge et al. [58] | 2023 | HA stabilized stem | Phase 1 | |
| Lee et al. [16] | 2023 | HA of (A/H1N1, A/H3N2, B/Victoria, and B/Yamagata) | phase 1/2 | |
| Xiong et al. [59] | 2023 | M2e, HA stalk, NP, | Mice |
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Global Influenza Programme. Available online: https://www.who.int/teams/global-influenza-programme/surveillance-and-monitoring/burden-of-disease (accessed on 9 September 2015).
- Saunders-Hastings, P.; Krewski, D. Reviewing the History of Pandemic Influenza: Understanding Patterns of Emergence and Transmission. Pathogens 2016, 5, 66. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Kash, J.C.; Morens, D.M. The 1918 influenza pandemic: 100 years of questions answered and unanswered. Sci. Transl. Med. 2019, 11, eaau5485. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Past Reported Global Human Cases with Highly Pathogenic Avian Influenza A(H5N1) (HPAI H5N1) by Country, 1997–2023; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2023.
- Chang, P.; Sadeyen, J.-R.; Bhat, S.; Daines, R.; Hussain, A.; Yilmaz, H.; Iqbal, M. Risk assessment of the newly emerged H7N9 avian influenza viruses. Emerg. Microbes Infect. 2023, 12, 2172965. [Google Scholar] [CrossRef]
- Krammer, F. The human antibody response to influenza A virus infection and vaccination. Nat. Rev. Immunol. 2019, 19, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.P.; Santos, M.; Martins, A.; Felgueiras, M.; Santos, R. Seasonal Influenza Vaccine Effectiveness in Persons Aged 15–64 Years: A Systematic Review and Meta-Analysis. Vaccines 2023, 11, 1322. [Google Scholar] [CrossRef]
- Harrington, W.N.; Kackos, C.M.; Webby, R.J. The evolution and future of influenza pandemic preparedness. Exp. Mol. Med. 2021, 53, 737–749. [Google Scholar] [CrossRef]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Meliopoulos, V.A.; Wang, W.; Lin, X.; Stucker, K.M.; Halpin, R.A.; Stockwell, T.B.; Schultz-Cherry, S.; Wentworth, D.E.; Dermody, T.S. Reversion of Cold-Adapted Live Attenuated Influenza Vaccine into a Pathogenic Virus. J. Virol. 2016, 90, 8454–8463. [Google Scholar] [CrossRef]
- Wang, W.-C.; Sayedahmed, E.E.; Sambhara, S.; Mittal, S.K. Progress towards the Development of a Universal Influenza Vaccine. Viruses 2022, 14, 1684. [Google Scholar] [CrossRef]
- Li, J.; Arevalo, M.T.; Chen, Y.; Chen, S.; Zeng, M. T-cell-mediated cross-strain protective immunity elicited by prime-boost vaccination with a live attenuated influenza vaccine. Int. J. Infect. Dis. 2014, 27, 37–43. [Google Scholar] [CrossRef]
- Melo, A.; de Macedo, L.S.; Invencao, M.; de Moura, I.A.; da Gama, M.; de Melo, C.M.L.; Silva, A.J.D.; Batista, M.V.A.; Freitas, A.C. Third-Generation Vaccines: Features of Nucleic Acid Vaccines and Strategies to Improve Their Efficiency. Genes 2022, 13, 2287. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Xia, F.; Chen, H.; Cui, B.; Feng, Y.; Zhang, P.; Chen, J.; Luo, M. A Guide to Nucleic Acid Vaccines in the Prevention and Treatment of Infectious Diseases and Cancers: From Basic Principles to Current Applications. Front. Cell Dev. Biol. 2021, 9, 633776. [Google Scholar] [CrossRef] [PubMed]
- William Ho, M.G.; Li, F.; Li, Z.; Zhang, X.-Q.; Xu, X. Next-Generation Vaccines Nanoparticle-Mediated DNA and mRNA Delivery. Adv. Healthc. Mater. 2021, 10, e2001812. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.T.; Nachbagauer, R.; Ensz, D.; Schwartz, H.; Carmona, L.; Schaefers, K.; Avanesov, A.; Stadlbauer, D.; Henry, C.; Chen, R.; et al. Safety and immunogenicity of a phase 1/2 randomized clinical trial of a quadrivalent, mRNA-based seasonal influenza vaccine (mRNA-1010) in healthy adults: Interim analysis. Nat. Commun. 2023, 14, 3631. [Google Scholar] [CrossRef] [PubMed]
- Boktor, S.W.; Hafner, J.W. Influenza. 2023 Jan 23. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Nuwarda, R.F.; Alharbi, A.A.; Kayser, V. An Overview of Influenza Viruses and Vaccines. Vaccines 2021, 9, 1032. [Google Scholar] [CrossRef] [PubMed]
- Kuchipudi, S.; Nissly, R. Novel Flu Viruses in Bats and Cattle: “Pushing the Envelope” of Influenza Infection. Vet. Sci. 2018, 5, 71. [Google Scholar] [CrossRef] [PubMed]
- Fodor, E.; te Velthuis, A.J.W. Structure and Function of the Influenza Virus Transcription and Replication Machinery. Cold Spring Harb. Perspect. Med. 2020, 10, a038398. [Google Scholar] [CrossRef] [PubMed]
- Boivin, S.; Cusack, S.; Ruigrok, R.W.H.; Hart, D.J. Influenza A Virus Polymerase: Structural Insights into Replication and Host Adaptation Mechanisms. J. Biol. Chem. 2010, 285, 28411–28417. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.E.; Flyer, D.C.; Raghunandan, R.; Liu, Y.; Wei, Z.; Wu, Y.; Kpamegan, E.; Courbron, D.; Fries, L.F.; Glenn, G.M. Development of influenza H7N9 virus like particle (VLP) vaccine: Homologous A/Anhui/1/2013 (H7N9) protection and heterologous A/chicken/Jalisco/CPA1/2012 (H7N3) cross-protection in vaccinated mice challenged with H7N9 virus. Vaccine 2013, 31, 4305–4313. [Google Scholar] [CrossRef]
- Manicassamy, B.; Kang, H.-J.; Chu, K.-B.; Lee, D.-H.; Lee, S.-H.; Park, B.R.; Kim, M.-C.; Kang, S.-M.; Quan, F.-S. Influenza M2 virus-like particle vaccination enhances protection in combination with avian influenza HA VLPs. PLoS ONE 2019, 14, e0216871. [Google Scholar] [CrossRef]
- Schwartzman, L.M.; Cathcart, A.L.; Pujanauski, L.M.; Qi, L.; Kash, J.C.; Taubenberger, J.K.; Dermody, T.S. An Intranasal Virus-Like Particle Vaccine Broadly Protects Mice from Multiple Subtypes of Influenza A Virus. mBio 2015, 6, e01044-15. [Google Scholar] [CrossRef] [PubMed]
- Nicole, M.; Bouvier, P.P. The biology of influenza viruses. Vaccine 2008, 26, 49–53. [Google Scholar]
- Lemey, P.; Langat, P.; Raghwani, J.; Dudas, G.; Bowden, T.A.; Edwards, S.; Gall, A.; Bedford, T.; Rambaut, A.; Daniels, R.S.; et al. Genome-wide evolutionary dynamics of influenza B viruses on a global scale. PLoS Pathog. 2017, 13, e1006749. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.C.; Wilson, I.A. Influenza Hemagglutinin Structures and Antibody Recognition. Cold Spring Harb. Perspect. Med. 2020, 10, a038778. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Liu, S.; Du, L.; Jiang, S. A new role of neuraminidase (NA) in the influenza virus life cycle: Implication for developing NA inhibitors with novel mechanism of action. Rev. Med. Virol. 2016, 26, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Sempere Borau, M.; Stertz, S. Entry of influenza A virus into host cells—Recent progress and remaining challenges. Curr. Opin. Virol. 2021, 48, 23–29. [Google Scholar] [CrossRef]
- Peukes, J.; Xiong, X.; Erlendsson, S.; Qu, K.; Wan, W.; Calder, L.J.; Schraidt, O.; Kummer, S.; Freund, S.M.V.; Kräusslich, H.-G.; et al. The native structure of the assembled matrix protein 1 of influenza A virus. Nature 2020, 587, 495–498. [Google Scholar] [CrossRef]
- Lee, S.-Y.; Kang, J.-O.; Chang, J. Nucleoprotein vaccine induces cross-protective cytotoxic T lymphocytes against both lineages of influenza B virus. Clin. Exp. Vaccine Res. 2019, 8, 54–63. [Google Scholar] [CrossRef]
- Mosmann, T.R.; McMichael, A.J.; LeVert, A.; McCauley, J.W.; Almond, J.W. Opportunities and challenges for T cell-based influenza vaccines. Nat. Rev. Immunol. 2024. [Google Scholar] [CrossRef]
- Lee, Y.-N.; Kim, M.-C.; Lee, Y.-T.; Kim, Y.-J.; Kang, S.-M. Mechanisms of Cross-protection by Influenza Virus M2-based Vaccines. Immune Netw. 2015, 15, 213–221. [Google Scholar] [CrossRef]
- Lee, J.-S.; Chowdhury, M.Y.E.; Moon, H.-J.; Choi, Y.-K.; Talactac, M.R.; Kim, J.-H.; Park, M.-E.; Son, H.-Y.; Shin, K.-S.; Kim, C.-J. The highly conserved HA2 protein of the influenza a virus induces a cross protective immune response. J. Virol. Methods 2013, 194, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, F.; Yin, L.; Jiang, H.; Lu, X.; Bi, Y.; Zhang, W.; Shi, Y.; Burioni, R.; Tong, Z.; et al. Structural basis for a human broadly neutralizing influenza A hemagglutinin stem-specific antibody including H17/18 subtypes. Nat. Commun. 2022, 13, 7603. [Google Scholar] [CrossRef] [PubMed]
- Bajic, G.; Tonouchi, K.; Adachi, Y.; Suzuki, T.; Kuroda, D.; Nishiyama, A.; Yumoto, K.; Takeyama, H.; Suzuki, T.; Hashiguchi, T.; et al. Structural basis for cross-group recognition of an influenza virus hemagglutinin antibody that targets postfusion stabilized epitope. PLoS Pathog. 2023, 19, e1011554. [Google Scholar] [CrossRef] [PubMed]
- Taheri, M.; Nemattalab, M.; Mahjoob, M.; Hasan-alizadeh, E.; Zamani, N.; Nikokar, I.; Evazalipour, M.; Soltani Tehrani, B.; Shenagari, M. Toward a universal influenza virus vaccine: Some cytokines may fulfill the request. Cytokine 2021, 148, 155703. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, P.; Tchitchek, N.; Joly, C.; Rodriguez Pozo, A.; Stimmer, L.; Langlois, S.; Hocini, H.; Gosse, L.; Pejoski, D.; Cosma, A.; et al. Vaccine Inoculation Route Modulates Early Immunity and Consequently Antigen-Specific Immune Response. Front. Immunol. 2021, 12, 645210. [Google Scholar] [CrossRef]
- Saiz, J.-C. Animal and Human Vaccines against West Nile Virus. Pathogens 2020, 9, 1073. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct Gene Transfer into Mouse Muscle in Vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Maruggi, G.; Shan, H.; Li, J. Advances in mRNA Vaccines for Infectious Diseases. Front. Immunol. 2019, 10, 594. [Google Scholar] [CrossRef] [PubMed]
- Gabriela Baldeon Vaca, M.M.; Cadete, A.; Hsiao, C.J.; Golding, A.; Jeon, A.; Jacquinet, E.; Azcue, E.; Guan, C.M.; Sanchez-Felix, X.; Pietzsch, C.A.; et al. Intranasal mRNA-LNP vaccination protects hamsters from SARS-CoV-2 infection. Sci. Adv. 2023, 9, eadh1655. [Google Scholar] [CrossRef]
- Alexandra Suberi, M.K.G.; Mao, T.; Israelow, B.; Reschke, M.; Grundler, J.; Akhtar, L.; Lee, T.; Shin, K.; Piotrowski-Daspit, A.S.; Homer, R.J.; et al. Polymer nanoparticles deliver mRNA to the lung for mucosal vaccination. Sci. Transl. Med. 2023, 15, eabq0603. [Google Scholar] [CrossRef]
- Thapa, H.B.; Müller, A.M.; Camilli, A.; Schild, S. An Intranasal Vaccine Based on Outer Membrane Vesicles Against SARS-CoV-2. Front. Microbiol. 2021, 12, 752739. [Google Scholar] [CrossRef] [PubMed]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol. Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, S.; Shahsavandi, S.; Maddadgar, O. Improvement influenza HA2 DNA vaccine cellular and humoral immune responses with Mx bio adjuvant. Biologicals 2017, 46, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Khanna, M.; Sharma, S.; Kumar, B.; Rajput, R. Protective Immunity Based on the Conserved Hemagglutinin Stalk Domain and Its Prospects for Universal Influenza Vaccine Development. BioMed Res. Int. 2014, 2014, 546274. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Li, R.; Deng, Y.; Lu, N.; Chen, H.; Meng, X.; Wang, W.; Wang, X.; Yan, K.; Qi, X.; et al. Protective Efficacy of the Conserved NP, PB1, and M1 Proteins as Immunogens in DNA- and Vaccinia Virus-Based Universal Influenza A Virus Vaccines in Mice. Clin. Vaccine Immunol. 2015, 22, 618–630. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wang, X.; Zheng, M.; Xiong, F.; Liu, X.; Zhou, L.; Tan, W.; Chen, Z. Immunization with DNA prime-subunit protein boost strategy based on influenza H9N2 virus conserved matrix protein M1 and its epitope screening. Sci. Rep. 2020, 10, 4144. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Wang, H.; Chen, J.; Shao, Z.; He, B.; Chen, J.; Lan, J.; Chen, Q.; Chen, Z. Protection against homo and hetero-subtypic influenza A virus by optimized M2e DNA vaccine. Emerg. Microbes Infect. 2019, 8, 45–54. [Google Scholar] [CrossRef]
- Hollý, J.; Tomčíková, K.; Vozárová, M.; Fogelová, M.; Jakubcová, L.; Varečková, E.; Kostolanský, F. DNA vaccine targeting the ectodomain of influenza M2 protein to endolysosome pathway enhances anti-M2e protective antibody response in mice. Acta Virol. 2021, 65, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Freyn, A.W.; Ramos da Silva, J.; Rosado, V.C.; Bliss, C.M.; Pine, M.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; de Souza Ferreira, L.C.; Weissman, D.; et al. A Multi-Targeting, Nucleoside-Modified mRNA Influenza Virus Vaccine Provides Broad Protection in Mice. Mol. Ther. 2020, 28, 1569–1584. [Google Scholar] [CrossRef]
- McMahon, M.; O’Dell, G.; Tan, J.; Sárközy, A.; Vadovics, M.; Carreño, J.M.; Puente-Massaguer, E.; Muramatsu, H.; Bajusz, C.; Rijnink, W.; et al. Assessment of a quadrivalent nucleoside-modified mRNA vaccine that protects against group 2 influenza viruses. Proc. Natl. Acad. Sci. USA 2022, 119, e2206333119. [Google Scholar] [CrossRef]
- Pardi, N.; Carreno, J.M.; O’Dell, G.; Tan, J.; Bajusz, C.; Muramatsu, H.; Rijnink, W.; Strohmeier, S.; Loganathan, M.; Bielak, D.; et al. Development of a pentavalent broadly protective nucleoside-modified mRNA vaccine against influenza B viruses. Nat. Commun. 2022, 13, 4677. [Google Scholar] [CrossRef] [PubMed]
- Arevalo, C.P.; Bolton, M.J.; Le Sage, V.; Ye, N.; Furey, C.; Muramatsu, H.; Alameh, M.-G.; Pardi, N.; Drapeau, E.M.; Parkhouse, K.; et al. A multivalent nucleoside-modified mRNA vaccine against all known influenza virus subtypes. Science 2022, 378, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Koen van de Ven, J.L.; van Dijken, H.; Muramatsu, H.; de Melo, C.V.B.; Lenz, S.; Peters, F.; Beattie, M.B.; Lin, P.J.C.; Ferreira, J.A.; van den Brand, J.; et al. A universal influenza mRNA vaccine candidate boosts T cell responses and reduces zoonotic influenza virus disease in ferrets. Sci. Adv. 2022, 8, eadc9937. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Wei, L.; Dong, C.; Wang, Y.; Kim, J.; Ma, Y.; Gonzalez, G.X.; Wang, B.-Z. cGAMP-adjuvanted multivalent influenza mRNA vaccines induce broadly protective immunity through cutaneous vaccination in mice. Mol. Ther. Nucleic Acids 2022, 30, 421–437. [Google Scholar] [CrossRef] [PubMed]
- Widge, A.T.; Hofstetter, A.R.; Houser, K.V.; Awan, S.F.; Chen, G.L.; Burgos Florez, M.C.; Berkowitz, N.M.; Mendoza, F.; Hendel, C.S.; Holman, L.A.; et al. An influenza hemagglutinin stem nanoparticle vaccine induces cross-group 1 neutralizing antibodies in healthy adults. Sci. Transl. Med. 2023, 15, eade4790. [Google Scholar] [CrossRef] [PubMed]
- Xiong, F.; Zhang, C.; Shang, B.; Zheng, M.; Wang, Q.; Ding, Y.; Luo, J.; Li, X. An mRNA-based broad-spectrum vaccine candidate confers cross-protection against heterosubtypic influenza A viruses. Emerg. Microbes Infect. 2023, 12, 2256422. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Lim, J.M.; Yu, B.; Song, S.; Neeli, P.; Sobhani, N.; Pavithra, K.; Bonam, S.R.; Kurapati, R.; Zheng, J.; et al. The next-generation DNA vaccine platforms and delivery systems: Advances, challenges and prospects. Front. Immunol. 2024, 15, 1332939. [Google Scholar] [CrossRef]
- Li, J.; Song, M.; Guo, D.; Yi, Y. Safety and Considerations of the COVID-19 Vaccine Massive Deployment. Virol. Sin. 2021, 36, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Ren, W.; Wu, T.; Wang, Q.; Luo, M.; Yi, Y.; Li, J. Real-World Safety of COVID-19 mRNA Vaccines: A Systematic Review and Meta-Analysis. Vaccines 2023, 11, 1118. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zeng, X.; Ren, W.; Liao, P.; Zhu, B. Protective efficacy of a universal influenza mRNA vaccine against the challenge of H1 and H5 influenza A viruses in mice. mLife 2023, 2, 308–316. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Ren, W.; Wang, Q.; Li, J. Advances in Nucleic Acid Universal Influenza Vaccines. Vaccines 2024, 12, 664. https://doi.org/10.3390/vaccines12060664
Xu L, Ren W, Wang Q, Li J. Advances in Nucleic Acid Universal Influenza Vaccines. Vaccines. 2024; 12(6):664. https://doi.org/10.3390/vaccines12060664
Chicago/Turabian StyleXu, Liang, Weigang Ren, Qin Wang, and Junwei Li. 2024. "Advances in Nucleic Acid Universal Influenza Vaccines" Vaccines 12, no. 6: 664. https://doi.org/10.3390/vaccines12060664
APA StyleXu, L., Ren, W., Wang, Q., & Li, J. (2024). Advances in Nucleic Acid Universal Influenza Vaccines. Vaccines, 12(6), 664. https://doi.org/10.3390/vaccines12060664