
Strategies for Enhancement of Live-Attenuated Salmonella-Based Carrier Vaccine Immunogenicity


Abstract
:1. Introduction
2. A summary of S. Typhi-based Carrier Vaccine Performance in Clinical Trials
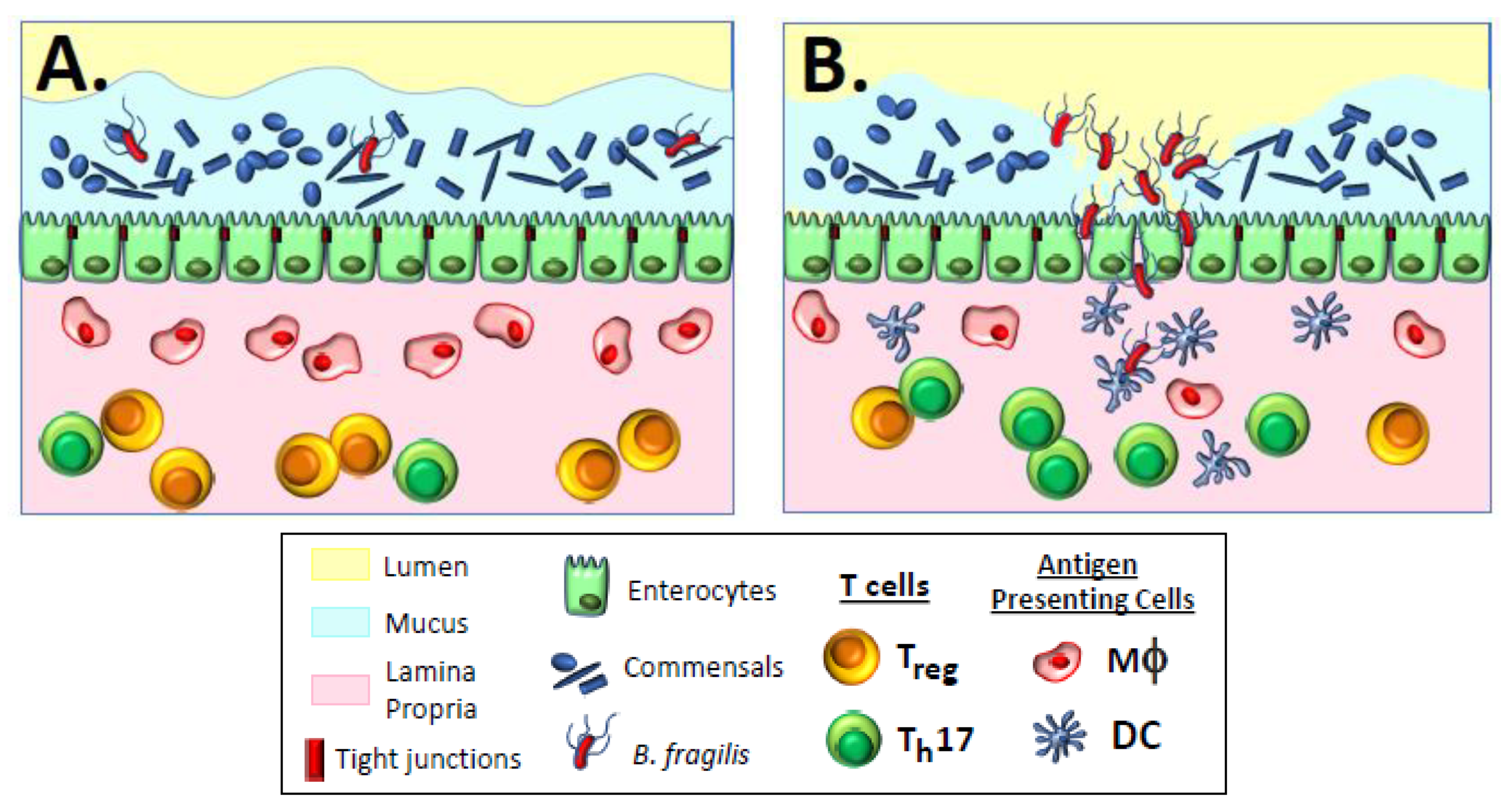
3. Why Are Commensal Bacteria Tolerogenic?
4. Why Are Reactogenic Vaccines Highly Immunogenic?
5. Balancing Attenuation and Immunogenicity in Carrier Vaccines
5.1. Parental Strain Factors
5.2. Antigen Expression Factors
5.3. Immunization Strategies
5.4. Host Factors
6. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nardelli-Haefliger, D.; Kraehenbuhl, J.-P.; Curtiss, R., III; Schodel, F.; Potts, A.; Kelly, S.M.; de Grandi, P. Oral and rectal immunization of adult female volunteers with a recombinant attenuated Salmonella typhi vaccine strain. Infect. Immun. 1996, 64, 5219–5224. [Google Scholar] [CrossRef] [Green Version]
- Tramont, E.C.; Chung, R.; Berman, S.; Keren, D.; Kapfer, C.; Formal, S.B. Safety and antigenicity of typhoid-Shigella sonnei vaccine (strain 5076-1C). J. Infect. Dis. 1984, 149, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Losonsky, G.; Herrington, D.; Berman, S.; Formal, S.B. Prevention of shigellosis by a Salmonella typhi-Shigella sonnei bivalent vaccine. J. Infect. Dis. 1987, 155, 1260–1265. [Google Scholar] [CrossRef] [PubMed]
- Herrington, D.A.; Van De Verg, L.; Formal, S.B.; Hale, T.L.; Tall, B.D.; Cryz, S.J.; Tramont, E.C.; Levine, M.M. Studies in volunteers to evaluate candidate Shigella vaccines: Further experience with a bivalent Salmonella typhi-Shigella sonnei vaccine and protection conferred by previous Shigella sonnei disease. Vaccine 1990, 8, 353–357. [Google Scholar] [CrossRef]
- Tacket, C.O.; Forrest, B.; Morona, R.; Attridge, S.R.; LaBrooy, J.; Tall, B.D.; Reymann, M.; Rowley, D.; Levine, M.M. Safety, immunogenicity, and efficacy against cholera challenge in humans of a typhoid-cholera hybrid vaccine derived from Salmonella typhi Ty21a. Infect. Immun. 1990, 58, 1620–1627. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, C.; Hone, D.M.; Noriega, F.; Tacket, C.O.; Davis, J.R.; Losonsky, G.; Nataro, J.P.; Hoffman, S.; Malik, A.; Nardin, E. Salmonella typhi vaccine strain CVD 908 expressing the circumsporozoite protein of Plasmodium falciparum: Strain construction and safety and immunogenicity in humans. J. Infect. Dis. 1994, 169, 927–931. [Google Scholar] [CrossRef] [PubMed]
- Tacket, C.O.; Galen, J.; Sztein, M.B.; Losonsky, G.; Wyant, T.L.; Nataro, J.; Wasserman, S.S.; Edelman, R.; Chatfield, S.; Dougan, G.; et al. Safety and immune responses to attenuated Salmonella enterica serovar Typhi oral live vector vaccines expressing tetanus toxin fragment C. Clin. Immunol. 2000, 97, 146–153. [Google Scholar] [CrossRef]
- DiPetrillo, M.D.; Tibbetts, T.; Kleanthous, H.; Killeen, K.P.; Hohmann, E.L. Safety and immunogenicity of phoP/phoQ-deleted Salmonella typhi expressing Helicobacter pylori urease in adult volunteers. Vaccine 1999, 18, 449–459. [Google Scholar] [CrossRef]
- Bumann, D.; Metzger, W.G.; Mansouri, E.; Palme, O.; Wendland, M.; Hurwitz, R.; Haas, G.; Aebischer, T.; von Specht, B.U.; Meyer, T.F. Safety and immunogenicity of live recombinant Salmonella enterica serovar Typhi Ty21a expressing urease A and B from Helicobacter pylori in human volunteers. Vaccine 2001, 20, 845–852. [Google Scholar] [CrossRef]
- Metzger, W.G.; Mansouri, E.; Kronawitter, M.; Diescher, S.; Soerensen, M.; Hurwitz, R.; Bumann, D.; Aebischer, T.; Von Specht, B.U.; Meyer, T.F. Impact of vector-priming on the immunogenicity of a live recombinant Salmonella enterica serovar typhi Ty21a vaccine expressing urease A and B from Helicobacter pylori in human volunteers. Vaccine 2004, 22, 2273–2277. [Google Scholar] [CrossRef]
- Khan, S.; Chatfield, S.; Stratford, R.; Bedwell, J.; Bentley, M.; Sulsh, S.; Giemza, R.; Smith, S.; Bongard, E.; Cosgrove, C.A.; et al. Ability of SPI2 mutant of S. typhi to effectively induce antibody responses to the mucosal antigen enterotoxigenic E. coli heat labile toxin B subunit after oral delivery to humans. Vaccine 2007, 25, 4175–4182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bumann, D.; Behre, C.; Behre, K.; Herz, S.; Gewecke, B.; Gessner, J.E.; von Specht, B.U.; Baumann, U. Systemic, nasal and oral live vaccines against Pseudomonas aeruginosa: A clinical trial of immunogenicity in lower airways of human volunteers. Vaccine 2010, 28, 707–713. [Google Scholar] [CrossRef]
- Frey, S.E.; Lottenbach, K.R.; Hill, H.; Blevins, T.P.; Yu, Y.; Zhang, Y.; Brenneman, K.E.; Kelly-Aehle, S.M.; McDonald, C.; Jansen, A.; et al. A Phase 1 dose-escalation trial in adults of three recombinant attenuated Salmonella Typhi vaccine vectors producing Streptococcus pneumoniae surface protein antigen PspA. Vaccine 2013, 31, 4874–4880. [Google Scholar] [CrossRef]
- Mowat, A.M.; Agace, W.W. Regional specialization within the intestinal immune system. Nat. Rev. Immunol. 2014, 14, 667–685. [Google Scholar] [CrossRef]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Sorbara, M.T.; Pamer, E.G. Interbacterial mechanisms of colonization resistance and the strategies pathogens use to overcome them. Mucosal. Immunol. 2019, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Geva-Zatorsky, N.; Sefik, E.; Kua, L.; Pasman, L.; Tan, T.G.; Ortiz-Lopez, A.; Yanortsang, T.B.; Yang, L.; Jupp, R.; Mathis, D.; et al. Mining the Human Gut Microbiota for Immunomodulatory Organisms. Cell 2017, 168, 928–943.e11. [Google Scholar] [CrossRef] [Green Version]
- Daliri, E.B.; Wei, S.; Oh, D.H.; Lee, B.H. The human microbiome and metabolomics: Current concepts and applications. Crit. Rev. Food Sci. Nutr. 2017, 57, 3565–3576. [Google Scholar] [CrossRef]
- Iacob, S.; Iacob, D.G. Infectious Threats, the Intestinal Barrier, and Its Trojan Horse: Dysbiosis. Front. Microbiol. 2019, 10, 1676. [Google Scholar] [CrossRef] [PubMed]
- Pandiyan, P.; Bhaskaran, N.; Zou, M.; Schneider, E.; Jayaraman, S.; Huehn, J. Microbiome Dependent Regulation of T(regs) and Th17 Cells in Mucosa. Front. Immunol. 2019, 10, 426. [Google Scholar] [CrossRef] [Green Version]
- Goto, Y. Epithelial Cells as a Transmitter of Signals from Commensal Bacteria and Host Immune Cells. Front. Immunol. 2019, 10, 2057. [Google Scholar] [CrossRef] [Green Version]
- Bunker, J.J.; Bendelac, A. IgA Responses to Microbiota. Immunity 2018, 49, 211–224. [Google Scholar] [CrossRef] [Green Version]
- Valguarnera, E.; Wardenburg, J.B. Good Gone Bad: One Toxin Away From Disease for Bacteroides fragilis. J. Mol. Biol. 2020, 432, 765–785. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; He, C.; Cong, Y.; Liu, Z. Regulatory immune cells in regulation of intestinal inflammatory response to microbiota. Mucosal. Immunol. 2015, 8, 969–978. [Google Scholar] [CrossRef]
- Alroqi, F.J.; Chatila, T.A. T Regulatory Cell Biology in Health and Disease. Curr. Allergy Asthma Rep. 2016, 16, 27. [Google Scholar] [CrossRef] [PubMed]
- Omenetti, S.; Pizarro, T.T. The Treg/Th17 Axis: A Dynamic Balance Regulated by the Gut Microbiome. Front. Immunol. 2015, 6, 639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kmiec, Z.; Cyman, M.; Slebioda, T.J. Cells of the innate and adaptive immunity and their interactions in inflammatory bowel disease. Adv. Med. Sci. 2017, 62, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sanders, T.J.; Yrlid, U.; Maloy, K.J. Intestinal Mononuclear Phagocytes in Health and Disease. Microbiol Spectrum. 2017. [Google Scholar] [CrossRef]
- Grainger, J.R.; Askenase, M.H.; Guimont-Desrochers, F.; da Fonseca, D.M.; Belkaid, Y. Contextual functions of antigen-presenting cells in the gastrointestinal tract. Immunol. Rev. 2014, 259, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, C.C.; Mowat, A.M. Macrophages in intestinal homeostasis and inflammation. Immunol. Rev. 2014, 260, 102–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, O.; van Baarlen, P.; Wells, J.M. Host-recognition of pathogens and commensals in the mammalian intestine. Curr. Top. Microbiol. Immunol. 2013, 358, 291–321. [Google Scholar]
- Macpherson, A.J.; Uhr, T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 2004, 303, 1662–1665. [Google Scholar] [CrossRef] [Green Version]
- Mathias, A.; Pais, B.; Favre, L.; Benyacoub, J.; Corthesy, B. Role of secretory IgA in the mucosal sensing of commensal bacteria. Gut Microbes 2014, 5, 688–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rol, N.; Favre, L.; Benyacoub, J.; Corthesy, B. The role of secretory immunoglobulin A in the natural sensing of commensal bacteria by mouse Peyer’s patch dendritic cells. J. Biol. Chem. 2012, 287, 40074–40082. [Google Scholar] [CrossRef] [Green Version]
- Flannigan, K.L.; Geem, D.; Harusato, A.; Denning, T.L. Intestinal Antigen-Presenting Cells: Key Regulators of Immune Homeostasis and Inflammation. Am. J. Pathol. 2015, 185, 1809–1819. [Google Scholar] [CrossRef]
- Worbs, T.; Hammerschmidt, S.I.; Forster, R. Dendritic cell migration in health and disease. Nat. Rev. Immunol. 2017, 17, 30–48. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Ning, M.X.; Chen, D.K.; Ma, W.T. Interactions between the Gut Microbiota and the Host Innate Immune Response Against Pathogens. Front. Immunol. 2019, 10, 607. [Google Scholar] [CrossRef] [Green Version]
- Caballero, S.; Pamer, E.G. Microbiota-mediated inflammation and antimicrobial defense in the intestine. Annu. Rev. Immunol. 2015, 33, 227–256. [Google Scholar] [CrossRef] [Green Version]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting edge: Bacterial flagellin activates basolaterally expressed TLR5 to induce epithelial proinflammatory gene expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, F.; Wu, W.; Cao, A.T.; Xue, X.; Yao, S.; Evans-Marin, H.L.; Li, Y.-Q.; Cong , Y. TLR5 mediates CD172alpha(+) intestinal lamina propria dendritic cell induction of Th17 cells. Sci. Rep. 2016, 6, 22040. [Google Scholar] [CrossRef] [Green Version]
- Cullender, T.C.; Chassaing, B.; Janzon, A.; Kumar, K.; Muller, C.E.; Werner, J.J.; Angenent, L.T.; Bell, M.E.; Hay, A.G.; Peterson, D.A.; et al. Innate and adaptive immunity interact to quench microbiome flagellar motility in the gut. Cell Host Microbe 2013, 14, 571–581. [Google Scholar] [CrossRef] [Green Version]
- Killig, M.; Glatzer, T.; Romagnani, C. Recognition strategies of group 3 innate lymphoid cells. Front. Immunol. 2014, 5, 142. [Google Scholar] [CrossRef] [Green Version]
- Satoh-Takayama, N. Heterogeneity and diversity of group 3 innate lymphoid cells: New cells on the block. Int. Immunol. 2016, 28, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, S.; Lochner, M.; Satoh-Takayama, N.; Dulauroy, S.; Berard, M.; Kleinschek, M.; Cua, D.; Di Santo, J.P.; Eberl, G. RORgammat+ innate lymphoid cells regulate intestinal homeostasis by integrating negative signals from the symbiotic microbiota. Nat. Immunol. 2011, 12, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Blander, J.M.; Sander, L.E. Beyond pattern recognition: Five immune checkpoints for scaling the microbial threat. Nat. Rev. Immunol. 2012, 12, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Sander, L.E. Improved vaccines through targeted manipulation of the body’s immunological risk-assessment? Bioessays 2012, 34, 876–884. [Google Scholar] [CrossRef]
- Kasturi, S.P.; Skountzou, I.; Albrecht, R.A.; Koutsonanos, D.; Hua, T.; Nakaya, H.I.; Ravindran, R.; Stewart, S.; Alam, M.; Kwissa, M.; et al. Programming the magnitude and persistence of antibody responses with innate immunity. Nature 2011, 470, 543–547. [Google Scholar] [CrossRef]
- Gause, K.T.; Wheatley, A.K.; Cui, J.; Yan, Y.; Kent, S.J.; Caruso, F. Immunological Principles Guiding the Rational Design of Particles for Vaccine Delivery. ACS Nano 2017, 11, 54–68. [Google Scholar] [CrossRef]
- Kanekiyo, M.; Bu, W.; Joyce, M.G.; Meng, G.; Whittle, J.R.; Baxa, U.; Yamamoto, T.; Narpala, S.; Todd, J.P.; Rao, S.S.; et al. Rational Design of an Epstein-Barr Virus Vaccine Targeting the Receptor-Binding Site. Cell 2015, 162, 1090–1100. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Rondini, S.; Alfini, R.; Lanzilao, L.; Necchi, F.; Negrea, A.; Rossi, O.; Brandt, C.; Clare, S.; Mastroeni, P.; et al. Comparative immunogenicity and efficacy of equivalent outer membrane vesicle and glycoconjugate vaccines against nontyphoidal Salmonella. Proc. Natl. Acad. Sci. USA 2018, 115, 10428–10433. [Google Scholar] [CrossRef] [Green Version]
- Sander, L.E.; Davis, M.J.; Boekschoten, M.V.; Amsen, D.; Dascher, C.C.; Ryffel, B.; Swanson, J.A.; Muller, M.; Blander, J.M. Detection of prokaryotic mRNA signifies microbial viability and promotes immunity. Nature 2011, 474, 385–389. [Google Scholar] [CrossRef] [PubMed]
- Ugolini, M.; Sander, L.E. Dead or alive: How the immune system detects microbial viability. Curr. Opin. Immunol. 2019, 56, 60–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamoto, S.; Nagao-Kitamoto, H.; Kuffa, P.; Kamada, N. Regulation of virulence: The rise and fall of gastrointestinal pathogens. J. Gastroenterol. 2016, 51, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Azimi, T.; Zamirnasta, M.; Sani, M.A.; Soltan Dallal, M.M.; Nasser, A. Molecular Mechanisms of Salmonella Effector Proteins: A Comprehensive Review. Infect. Drug Resist. 2020, 13, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Gan, J.; Giogha, C.; Hartland, E.L. Molecular mechanisms employed by enteric bacterial pathogens to antagonise host innate immunity. Curr. Opin. Microbiol. 2020, 59, 58–64. [Google Scholar] [CrossRef]
- Rogers, A.W.L.; Tsolis, R.M.; Bäumler, A.J. Salmonella versus the Microbiome. Microbiol. Mol. Biol. Rev. 2021, 85, e00027-19. [Google Scholar] [CrossRef]
- Odendall, C.; Kagan, J.C. Activation and pathogenic manipulation of the sensors of the innate immune system. Microbes Infect. Inst. Pasteur 2017, 19, 229–237. [Google Scholar] [CrossRef]
- Kagan, J.C.; Medzhitov, R. Phosphoinositide-mediated adaptor recruitment controls Toll-like receptor signaling. Cell 2006, 125, 943–955. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Svenungsson, B.; Kärnell, A.; Weintraub, A. Prevalence of enterotoxigenic Bacteroides fragilis in adult patients with diarrhea and healthy controls. Clin. Infect. Dis. 1999, 29, 590–594. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Pu, J.; Lu, S.; Bai, X.; Wu, Y.; Jin, D.; Cheng, Y.; Zhang, G.; Zhu, W.; Luo, X.; et al. Species-Level Analysis of Human Gut Microbiota With Metataxonomics. Front. Microbiol. 2020, 11, 2029. [Google Scholar] [CrossRef]
- Hecht, A.L.; Casterline, B.W.; Choi, V.M.; Bubeck Wardenburg, J. A Two-Component System Regulates Bacteroides fragilis Toxin to Maintain Intestinal Homeostasis and Prevent Lethal Disease. Cell Host Microbe 2017, 22, 443–448.e5. [Google Scholar] [CrossRef] [Green Version]
- Galen, J.E.; Gomez-Duarte, O.G.; Losonsky, G.; Halpern, J.L.; Lauderbaugh, C.S.; Kaintuck, S.; Reymann, M.K.; Levine, M.M. A murine model of intranasal immunization to assess the immunogenicity of attenuated Salmonella typhi live vector vaccines in stimulating serum antibody responses to expressed foreign antigens. Vaccine 1997, 15, 700–708. [Google Scholar] [CrossRef]
- Pasetti, M.F.; Pickett, T.E.; Levine, M.M.; Sztein, M.B. A comparison of immunogenicity and in vivo distribution of Salmonella enterica serovar Typhi and Typhimurium live vector vaccines delivered by mucosal routes in the murine model. Vaccine 2000, 18, 3208–3213. [Google Scholar] [CrossRef]
- Pickett, T.E.; Pasetti, M.F.; Galen, J.E.; Sztein, M.B.; Levine, M.M. In vivo characterization of the murine intranasal model for assessing the immunogenicity of attenuated Salmonella enterica serovar Typhi strains as live mucosal vaccines and as live vectors. Infect. Immun. 2000, 68, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tacket, C.O.; Sztein, M.; Losonsky, G.; Wasserman, S.S.; Nataro, J.P.; Edelman, R.; Pickard, D.; Dougan, G.; Chatfield, S.; Levine, M.M.; et al. Safety of live oral Salmonella typhi vaccine strains with deletions in htrA and aroC aroD and immune responses in humans. Infect. Immun. 1997, 65, 452–456. [Google Scholar] [CrossRef] [Green Version]
- Tacket, C.O.; Sztein, M.; Wasserman, S.S.; Losonsky, G.; Kotloff, K.; Wyant, T.L.; Nataro, J.P.; Edelman, R.; Perry, J.G.; Bedford, P.; et al. Phase 2 clinical trial of attenuated Salmonella enterica serovar Typhi oral live vector vaccine CVD 908-htrA in U.S. volunteers. Infect. Immun. 2000, 68, 1196–1201. [Google Scholar] [CrossRef] [Green Version]
- Tacket, C.O.; Hone, D.M.; Losonsky, G.A.; Guers, L.; Edelman, R.; Levine, M.M. Clinical acceptability and immunogenicity of CVD 908 Salmonella typhi vaccine strain. Vaccine 1992, 10, 443–446. [Google Scholar] [CrossRef]
- Tacket, C.O.; Hone, D.M.; Curtiss Iii, R.; Kelly, S.M.; Losonsky, G.; Guers, L.; Harris, A.M.; Edelman, R.; Levine, M.M. Comparison of the safety and immunogenicity of ΔaroC ΔaroD and Δcya Δcrp Salmonella typhi strains in adult volunteers. Infect. Immun. 1992, 60, 536–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galen, J.E.; Curtiss, R., 3rd. The delicate balance in genetically engineering live vaccines. Vaccine. 2014, 32, 4376–4385. [Google Scholar] [CrossRef] [Green Version]
- Germanier, R.; Fuer, E. Isolation and characterization of galE mutant Ty 21a of Salmonella typhi: A candidate strain for a live, oral typhoid vaccine. J. Infect. Dis. 1975, 131, 553–558. [Google Scholar] [CrossRef]
- Kopecko, D.J.; Sieber, H.; Ures, J.A.; Furer, A.; Schlup, J.; Knof, U.; Collioud, A.; Xu, D.; Colburn, K.; Dietrich, G. Genetic stability of vaccine strain Salmonella Typhi Ty21a over 25 years. Int. J. Med. Microbiol. 2009, 299, 233–246. [Google Scholar] [CrossRef]
- Galen, J.E.; Levine, M.M. Can a ‘flawless’ live vector vaccine strain be engineered? Trends Microbiol. 2001, 9, 372–376. [Google Scholar] [CrossRef]
- Galen, J.E.; Pasetti, M.F.; Tennant, S.; Ruiz-Olvera, P.; Sztein, M.B.; Levine, M.M. Salmonella enterica serovar Typhi live vector vaccines finally come of age. Immunol. Cell Biol. 2009, 87, 400–412. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kong, Q.; Curtiss, R., 3rd. New technologies in developing recombinant attenuated Salmonella vaccine vectors. Microb. Pathog. 2013, 58, 17–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roland, K.L.; Brenneman, K.E. Salmonella as a vaccine delivery vehicle. Expert Rev. Vaccines 2013, 12, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.M.; Tacket, C.O.; Sztein, M.B. Host-Salmonella interaction: Human trials. Microbes Infect. 2001, 3, 1271–1279. [Google Scholar] [CrossRef]
- Sztein, M.; Tanner, M.K.; Polotsky, Y.; Orenstein, J.M.; Levine, M.M. Cytotoxic T lymphocytes after oral immunization with attenuated vaccine strains of Salmonella typhi in humans. J. Immunol. 1995, 155, 3987–3993. [Google Scholar]
- Sztein, M.B. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica Serovar Typhi strains used as live oral vaccines in humans. Clin Infect. Dis. 2007, 45, S15–S19. [Google Scholar] [CrossRef] [Green Version]
- Sztein, M.B.; Salerno-Goncalves, R.; McArthur, M.A. Complex adaptive immunity to enteric fevers in humans: Lessons learned and the path forward. Front. Immunol. 2014, 5, 516. [Google Scholar] [CrossRef]
- Brinkmann, U.; Mattes, R.E.; Buckel, P. High-level expression of recombinant genes in Escherichia coli is dependent on the availability of the dnaY gene product. Gene 1989, 85, 109–114. [Google Scholar] [CrossRef]
- Zahn, K. Overexpression of an mRNA dependent on rare codons inhibits protein synthesis and cell growth. J. Bacteriol. 1996, 178, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Metabolic load and heterologous gene expression. Biotechnol. Adv. 1995, 13, 247–261. [Google Scholar] [CrossRef]
- Bailey, J.E. Host-vector interactions in Escherichia coli. In Advances in Biochemical Engineering Biotechnology; Fiechter, A., Ed.; Springer: Berlin, Germany, 1993; pp. 29–77. [Google Scholar]
- Boe, L.; Gerdes, K.; Molin, S. Effects of genes exerting growth inhibition and plasmid stability on plasmid maintenance. J. Bacteriol. 1987, 169, 4646–4650. [Google Scholar] [CrossRef] [Green Version]
- Corchero, J.L.; Villaverde, A. Plasmid maintenance in Escherichia coli recombinant cultures is dramatically, steadily, and specifically influenced by features of the encoded proteins. Biotechnol. Bioeng. 1998, 58, 625–632. [Google Scholar] [CrossRef]
- Galen, J.E.; Nair, J.; Wang, J.Y.; Wasserman, S.S.; Tanner, M.K.; Sztein, M.; Levine, M.M. Optimization of plasmid maintenance in the attenuated live vector vaccine strain Salmonella typhi CVD 908-htrA. Infect. Immun. 1999, 67, 6424–6433. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.; Li, J.; Bacon, A.; Chatfield, S. Oral vaccination against tetanus: Comparison of the immunogenicities of Salmonella strains expressing fragment C from the nirB and htrA promoters. Infect. Immun. 1998, 66, 3080–3087. [Google Scholar] [CrossRef] [Green Version]
- Curtiss, R., III; Galan, J.E.; Nakayama, K.; Kelly, S.M. Stabilization of recombinant avirulent vaccine strains in vivo. Res. Microbiol. 1990, 141, 797–805. [Google Scholar] [CrossRef]
- Tacket, C.O.; Kelley, S.M.; Schodel, F.; Losonsky, G.; Nataro, J.P.; Edelman, R.; Levine, M.M.; Curtiss III, R. Safety and immunogenicity in humans of an attenuated Salmonella typhi vaccine vector strain expressing plasmid-encoded hepatitis B antigens stabilized by the Asd-balanced lethal vector system. Infect. Immun. 1997, 65, 3381–3385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.-F.; Brahmbhatt, H.N.; de Lorenzo, V.; Wehland, J.; Timmis, K.N. Extracellular export of shiga toxin B-subunit/haemolysin A (C-terminus) fusion protein expressed in Salmonella typhimurium aroA- mutant and stimulation of B-subunit specific antibody responses in mice. Microb. Pathog. 1992, 13, 465–476. [Google Scholar] [CrossRef]
- Hess, J.; Gentschev, I.; Miko, D.; Welzel, M.; Ladel, C.; Goebel, W.; Kaufmann, S.H.E. Superior efficacy of secreted over somatic antigen display in recombinant Salmonella vaccine induced protection against listeriosis. Proc. Natl. Acad. Sci. USA 1996, 93, 1458–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzschaschel, B.D.; Guzman, C.A.; Timmis, K.N.; de Lorenzo, V. An Escherichia coli hemolysin transport system-based vector for the export of polypeptides: Export of Shiga-like toxin IIeB subunit by Salmonella typhimurium aroA. Nat. Biotechnol. 1996, 14, 765–769. [Google Scholar] [CrossRef] [PubMed]
- Russmann, H.; Shams, H.; Poblete, F.; Fu, Y.; Galan, J.E.; Donis, R.O. Delivery of epitopes by the Salmonella type III secretion system for vaccine development. Science 1998, 281, 565–568. [Google Scholar] [PubMed] [Green Version]
- Russmann, H.; Igwe, E.I.; Sauer, J.; Hardt, W.D.; Bubert, A.; Geginat, G. Protection against murine listeriosis by oral vaccination with recombinant Salmonella expressing hybrid Yersinia type III proteins. J. Immunol. 2001, 167, 357–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-S.; Shin, K.-S.; Pan, J.-G.; Kim, C.-J. Surface-displayed viral antigens on Salmonella carrier vaccine. Nat. Biotechnol. 2000, 18, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.Y.; Curtiss, R., III. Immune responses dependent on antigen location in recombinant attenuated Salmonella typhimurium vaccines following oral immunization. FEMS Immunol. Med. Microbiol. 2003, 37, 99–104. [Google Scholar] [CrossRef]
- Galen, J.E.; Zhao, L.; Chinchilla, M.; Wang, J.Y.; Pasetti, M.F.; Green, J.; Levine, M.M. Adaptation of the endogenous Salmonella enterica serovar Typhi clyA-encoded hemolysin for antigen export enhances the immunogenicity of anthrax protective antigen domain 4 expressed by the attenuated live-vector vaccine strain CVD 908-htrA. Infect. Immun. 2004, 72, 7096–7106. [Google Scholar] [CrossRef] [Green Version]
- Galen, J.E.; Chinchilla, M.; Pasetti, M.F.; Wang, J.Y.; Zhao, L.; Rciniega-Martinez, I.; Silverman, D.J.; Levine, M.M. Mucosal immunization with attenuated Salmonella enterica serovar Typhi expressing protective antigen of anthrax toxin (PA83) primes monkeys for accelerated serum antibody responses to parenteral PA83 vaccine. J. Infect. Dis. 2009, 199, 326–335. [Google Scholar] [CrossRef] [Green Version]
- Vindurampulle, C.J.; Attridge, S.R. Vector priming reduces the immunogenicity of Salmonella-based vaccines in Nramp1+/+ mice. Infect. Immun. 2003, 71, 2258–2261. [Google Scholar] [CrossRef] [Green Version]
- Vindurampulle, C.J.; Attridge, S.R. Impact of vector priming on the immunogenicity of recombinant Salmonella vaccines. Infect. Immun. 2003, 71, 287–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attridge, S.R.; Vindurampulle, C.J. Vector-primed mice display hypo-responsiveness to foreign antigen presented by recombinant Salmonella regardless of the route of delivery. Microb. Pathog. 2005, 39, 1–7. [Google Scholar] [CrossRef]
- Roberts, M.; Bacon, A.; Li, J.; Chatfield, S. Prior immunity to homologous and heterologous Salmonella serotypes suppresses local and systemic anti-fragment C antibody responses and protection from tetanus toxin in mice immunized with Salmonella strains expressing fragment C. Infect. Immun. 1999, 67, 3810–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.V.; Reece, W.; Gothard, P.; Moorthy, V.; Roberts, M.; Flanagan, K.; Plebanski, M.; Hannan, C.; Hu, J.T.; Anderson, R.; et al. DNA-based vaccines for malaria: A heterologous prime-boost immunisation strategy. Dev. Biol. (Basel) 2000, 104, 171–179. [Google Scholar]
- Ramshaw, I.A.; Ramsay, A.J. The prime-boost strategy: Exciting prospects for improved vaccination. Immunol. Today 2000, 21, 163–165. [Google Scholar] [CrossRef]
- Londono-Arcila, P.; Freeman, D.; Kleanthous, H.; O’Dowd, A.M.; Lewis, S.; Turner, A.K.; Rees, E.L.; Tibbitts, T.J.; Greenwood, J.; Monath, T.P.; et al. Attenuated Salmonella enterica serovar Typhi expressing urease effectively immunizes mice against Helicobacter pylori challenge as part of a heterologous mucosal priming-parenteral boosting vaccination regimen. Infect. Immun. 2002, 70, 5096–5106. [Google Scholar] [CrossRef] [Green Version]
- Vindurampulle, C.J.; Cuberos, L.F.; Barry, E.M.; Pasetti, M.F.; Levine, M.M. Recombinant Salmonella enterica serovar Typhi in a prime-boost strategy. Vaccine 2004, 22, 3744–3750. [Google Scholar] [CrossRef]
- Chinchilla, M.; Pasetti, M.F.; Medina-Moreno, S.; Wang, J.Y.; Gomez-Duarte, O.G.; Stout, R.; Levine, M.M.; Galen, J.E. Enhanced immunity to Plasmodium falciparum circumsporozoite protein (PfCSP) by using Salmonella enterica serovar Typhi expressing PfCSP and a PfCSP-encoding DNA vaccine in a heterologous prime-boost strategy. Infect. Immun. 2007, 75, 3769–3779. [Google Scholar] [CrossRef] [Green Version]
- Galen, J.E.; Wang, J.Y.; Carrasco, J.A.; Lloyd, S.A.; Mellado-Sanchez, G.; Diaz-McNair, J.; Franco, O.; Buskirk, A.D.; Nataro, J.P.; Pasetti, M.F. A Bivalent Typhoid Live Vector Vaccine Expressing both Chromosome- and Plasmid-Encoded Yersinia pestis Antigens Fully Protects against Murine Lethal Pulmonary Plague Infection. Infect. Immun. 2015, 83, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boraschi, D.; Italiani, P. Immunosenescence and vaccine failure in the elderly: Strategies for improving response. Immunol. Lett. 2014, 162, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhivaki, D.; Lo-Man, R. Unique aspects of the perinatal immune system. Nat. Rev. Immunol. 2017, 17, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Stelekati, E.; Wherry, E.J. Chronic bystander infections and immunity to unrelated antigens. Cell Host Microbe 2012, 12, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Uche, I.V.; MacLennan, C.A.; Saul, A. A Systematic Review of the Incidence, Risk Factors and Case Fatality Rates of Invasive Nontyphoidal Salmonella (iNTS) Disease in Africa (1966 to 2014). PLoS Negl. Trop. Dis. 2017, 11, e0005118. [Google Scholar] [CrossRef] [Green Version]
- Qadri, F.; Bhuiyan, T.R.; Sack, D.A.; Svennerholm, A.M. Immune responses and protection in children in developing countries induced by oral vaccines. Vaccine 2013, 31, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.M. Immunogenicity and efficacy of oral vaccines in developing countries: Lessons from a live cholera vaccine. BMC Biol. 2010, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Prendergast, A.J. Malnutrition and vaccination in developing countries. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donowitz, J.R.; Petri, W.A., Jr. Pediatric small intestine bacterial overgrowth in low-income countries. Trends Mol Med. 2015, 21, 6–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilmartin, A.A.; Petri, W.A., Jr. Exploring the role of environmental enteropathy in malnutrition, infant development and oral vaccine response. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140143. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Petri, W.A., Jr. Environmental Enteropathy: Elusive but Significant Subclinical Abnormalities in Developing Countries. EBioMedicine. 2016, 10, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Suharyono; Simanjuntak, C.; Witham, N.; Punjabi, N.; Heppner, D.G.; Losonsky, G.; Totosudirjo, H.; Rifai, A.R.; Clemens, J.; Lim, Y.L.; et al. Safety and immunogenicity of single-dose live oral cholera vaccine CVD 103-HgR in 5-9-year-old Indonesian children. Lancet 1992, 340, 689–694. [Google Scholar] [CrossRef]
- Gotuzzo, E.; Butron, B.; Seas, C.; Penny, M.; Ruiz, R.; Losonsky, G.; Lanata, C.F.; Wasserman, S.S.; Salazar, E.; Kaper, J.B.; et al. Safety, immunogenicity, and excretion pattern of single-dose live oral cholera vaccine CVD 103-HgR in Peruvian adults of high and low socioeconomic levels. Infect. Immun. 1993, 61, 3994–3997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos, R.; Fasano, A.; Wasserman, S.S.; Prado, V.; San Martin, O.; Abrego, P.; Losonsky, G.A.; Alegria, S.; Levine, M.M. Effect of small bowel bacterial overgrowth on the immunogenicity of single-dose live oral cholera vaccine CVD 103-HgR. J. Infect. Dis. 1999, 180, 1709–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, S.M.; Amadi, B.; Mwiya, M.; Nkamba, H.; Tomkins, A.; Goldblatt, D. Dendritic cell anergy results from endotoxemia in severe malnutrition. J. Immunol. 2009, 183, 2818–2826. [Google Scholar] [CrossRef] [Green Version]
- Pasetti, M.F.; Levine, M.M.; Sztein, M.B. Animal models paving the way for clinical trials of attenuated Salmonella enterica serovar Typhi live oral vaccines and live vectors. Vaccine 2003, 21, 401–418. [Google Scholar] [CrossRef]
- Jiang, B.; Li, Z.; Ou, B.; Duan, Q.; Zhu, G. Targeting ideal oral vaccine vectors based on probiotics: A systematical view. Appl. Microbiol. Biotechnol. 2019, 103, 3941–3953. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.L.; Hruby, D.E. Commensal bacteria as a novel delivery system for subunit vaccines directed against agents of bioterrorism. Adv. Drug Deliv. Rev. 2005, 57, 1392–1402. [Google Scholar] [CrossRef]


{kind=link}
| Vaccine A | Year(s) of Clinical Trial | Parent Strain; Relevant Genotype | Foreign Antigen | Expression (Stabilization) Method ‡ | CFU Per Dose; Number of Doses # | Foreign Antigen—Specific Igg Sero- Conversion Rates | Heterologous Booster Given? (Route) B | Ref |
|---|---|---|---|---|---|---|---|---|
| 5076-1C w/ plasmid | 1984, 1987, 1990 | Ty21a; Str R | Shigella sonnei O-antigen | P (ND) | ~109; 3 doses | 28/77 | NO | [2,3,4] |
| EX645 w/ plasmid | 1990 | Ty21a; ΔthyA rfaTy2::rfaE. coli RifR | O-antigen from Vibrio cholera O1 serotype Inaba | P (thyA) | ~1010; 3 doses | 1/14 | NO | [5] |
| CVD 908 | 1994 | Ty2; ΔrpoS ΔaroC ΔaroD | csp1 | C (NR) | 5 × 107; 2 doses | 2/10 | NO | [6] |
| χ4632(pYA3167) | 1996 | Ty2; Δcya Δ(crp-cdt) Δasd | HBcAg-pre-S1-pre-S2 2 | P (asd) | 3 × 109; 3 doses | 0/7 | NO | [1] |
| CVD 908-htrA w/ plasmid | 2000 | Ty2; ΔrpoS ΔaroC ΔaroD ΔhtrA | toxC3 | P (ND) | 108 to 109; 1 dose | 3/9 | NO | [7] |
| Ty1033 w/ plasmid | 2000 | Ty2; ΔrpoS ΔphoP ΔphoQ ΔpurB | ureA-ureB4 | P (purB) | >1010; 1 dose | 0/7 | NO | [8] |
| Ty21a(pDB1) | 2001, 2004 | Ty21a; ΔthyA | ureA-ureB | P (thyA) | ~6–9 × 109; 3 doses 1–2 × 1010; 3 doses | 0/9 0/9 | NO | [9,10] |
| TSB7 | 2007 | Ty2; ΔrpoS ΔaroC::PssaG-eltB ΔssaV | eltB5 | C (NR) | ~108; 2 doses ~109; 2 doses | 7/12 15/22 | NO | [11] |
| CVD 908 w/ plasmid | 2010 | Ty2; ΔrpoS ΔaroC ΔaroD ΔthyA | oprF-oprI fusion protein 6 | P (thyA) | ~108; 1 dose | 13/16 | YES (IM) | [12] |
| Ty21a w/ plasmid | 2010 | Ty21a; ΔthyA | oprF-oprI fusion protein 6 | P (thyA) | ~1010; 3 doses | 15/16 | YES (IM) | [12] |
| χ9639(pYA4088) C | 2013 | Ty2; rpoS− | pspA7 | P (asd) | 107–1010 in escalating single doses | 0/20 | NO | [13] |
| χ9640(pYA4088) C | 2013 | Ty2; rpoS+ | pspA | P (asd) | 107–1010 in escalating single doses | 0/20 | NO | [13] |
| Factor | Potential Problem |
|---|---|
| 1. Over-attenuation of the carrier strain itself | Reduced colonization/contact with immune inductive sites |
| 2. Ability of carrier to elicit humoral, cellular, and/or mucosal immunity | Failure to elicit biologically relevant foreign-pathogen-specific immunity |
| 3. Expression level of the foreign antigen; chromosomal versus plasmid-based expression | Metabolic burden over-attenuates carrier vaccine |
| 4. Location of the synthesized foreign antigen; cytoplasmic versus surface or exported | Intracellular degradation of antigen(s) reduces in situ delivery to immune inductive sites |
| 5. Inherent immunogenicity of the foreign antigen(s) | Insufficient stimulation of biologically relevant immunity |
| 6. Immunization strategy; dosing, timing, and homologous versus heterologous vaccination | Insufficient stimulation of biologically relevant immunity |
| 7. Host intestinal microbiota | Reduced colonization/contact with immune inductive sites |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galen, J.E.; Wahid, R.; Buskirk, A.D. Strategies for Enhancement of Live-Attenuated Salmonella-Based Carrier Vaccine Immunogenicity. Vaccines 2021, 9, 162. https://doi.org/10.3390/vaccines9020162
Galen JE, Wahid R, Buskirk AD. Strategies for Enhancement of Live-Attenuated Salmonella-Based Carrier Vaccine Immunogenicity. Vaccines. 2021; 9(2):162. https://doi.org/10.3390/vaccines9020162
Chicago/Turabian StyleGalen, James E., Rezwanul Wahid, and Amanda D. Buskirk. 2021. "Strategies for Enhancement of Live-Attenuated Salmonella-Based Carrier Vaccine Immunogenicity" Vaccines 9, no. 2: 162. https://doi.org/10.3390/vaccines9020162
APA StyleGalen, J. E., Wahid, R., & Buskirk, A. D. (2021). Strategies for Enhancement of Live-Attenuated Salmonella-Based Carrier Vaccine Immunogenicity. Vaccines, 9(2), 162. https://doi.org/10.3390/vaccines9020162