
ELISA-Based Analysis Reveals an Anti-SARS-CoV-2 Protein Immune Response Profile Associated with Disease Severity

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design and Participants
2.2. Clinical Variables of Interest
2.3. Linear B-Cell Epitope Prediction
2.4. Linear Peptide Library
2.5. Peptide-Base ELISA
2.6. Peptides of Interest
2.7. Statistical Analysis
2.8. Ethics Approval
3. Results
3.1. Characteristics of Participants
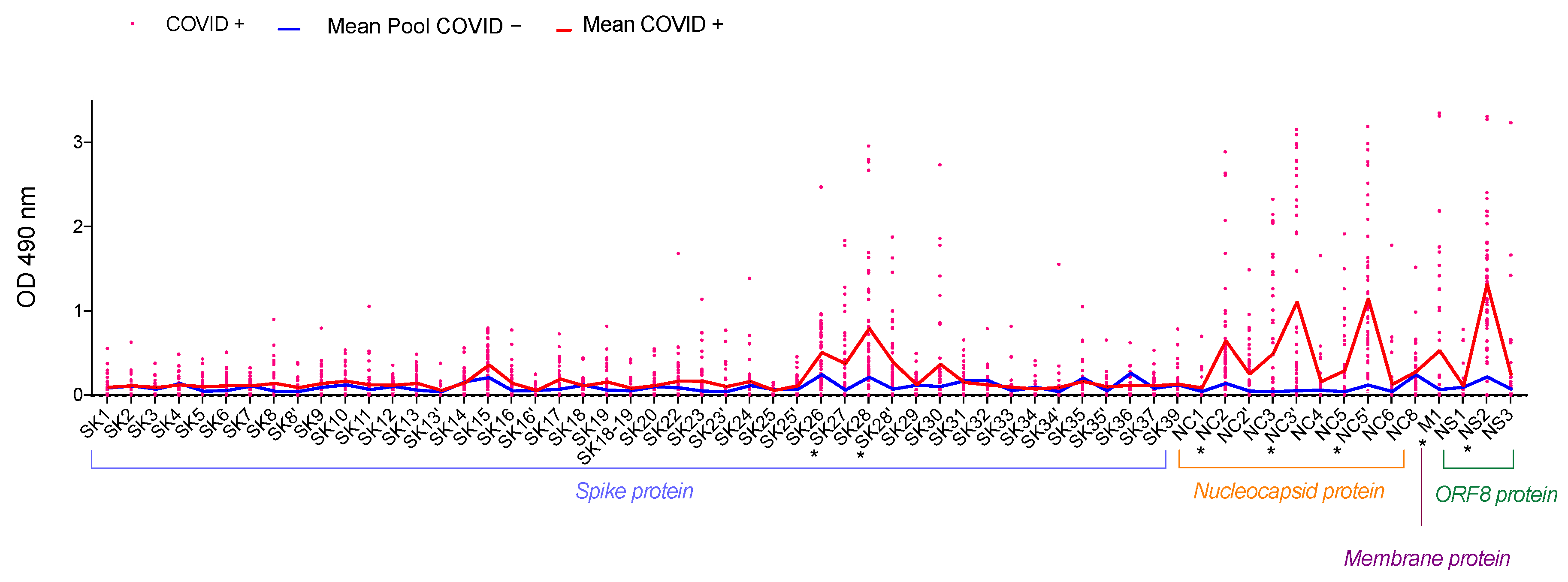
3.2. Selecting the Peptides of Interest
3.3. Analysis of the Selected Peptides
3.4. Analysis of Variables and Peptides Associated with Disease Severity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Q.; Guo, D. Emerging coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Helms, J.; Kremer, S.; Merdji, H.; Clere-Jehl, R.; Schenck, M.; Kummerlen, C.; Collange, O.; Boulay, C.; Fafi-Kremer, S.; Ohana, M.; et al. Neurologic Features in Severe SARS-CoV-2 Infection. N. Engl. J. Med. 2020, 382, 2268–2270. [Google Scholar] [CrossRef]
- Wang, D.; Hu, B.; Hu, C.; Zhu, F.; Liu, X.; Zhang, J.; Wang, B.; Xiang, H.; Cheng, Z.; Xiong, Y.; et al. Clinical Characteristics of 138 Hospitalized Patients With 2019 Novel Coronavirus–Infected Pneumonia in Wuhan, China. JAMA 2020, 323, 1061. [Google Scholar] [CrossRef] [PubMed]
- Rowley, A.H.; Shulman, S.T.; Arditi, M. Immune pathogenesis of COVID-19–related multisystem inflammatory syndrome in children. J. Clin. Investig. 2020, 130, 5619–5621. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Tang, K.; Levin, M.; Irfan, O.; Morris, S.K.; Wilson, K.; Klein, J.D.; Bhutta, Z.A. COVID-19 and multisystem inflammatory syndrome in children and adolescents. Lancet Infect. Dis. 2020, 20, e276–e288. [Google Scholar] [CrossRef]
- Lu, S.; Xie, X.; Zhao, L.; Wang, B.; Zhu, J.; Yang, T.; Yang, G.; Ji, M.; Lv, C.; Xue, J.; et al. The immunodominant and neutralization linear epitopes for SARS-CoV-2. Cell Rep. 2021, 34, 108666. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.-Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e9. [Google Scholar] [CrossRef]
- Astuti, I.; Ysrafil. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An overview of viral structure and host response. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 407–412. [Google Scholar] [CrossRef]
- Chai, J.; Cai, Y.; Pang, C.; Wang, L.; McSweeney, S.; Shanklin, J.; Liu, Q. Structural basis for SARS-CoV-2 envelope protein recognition of human cell junction protein PALS1. Nat. Commun. 2021, 12, 3433. [Google Scholar] [CrossRef]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 27, 671–680.e2. [Google Scholar] [CrossRef]
- Bai, Z.; Cao, Y.; Liu, W.; Li, J. The SARS-CoV-2 Nucleocapsid Protein and Its Role in Viral Structure, Biological Functions, and a Potential Target for Drug or Vaccine Mitigation. Viruses 2021, 13, 1115. [Google Scholar] [CrossRef]
- Dutta, N.K.; Mazumdar, K.; Gordy, J.T. The Nucleocapsid Protein of SARS–CoV-2: A Target for Vaccine Development. J. Virol. 2020, 94, e00647-20. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, Y.; Greig, M.; Liu, G.; Driedger, M.; Langlois, M.-A. Humoral Responses and Serological Assays in SARS-CoV-2 Infections. Front. Immunol. 2020, 11, 610688. [Google Scholar] [CrossRef]
- Ahmad, T.A.; Eweida, A.E.; Sheweita, S.A. B-cell epitope mapping for the design of vaccines and effective diagnostics. Trials Vaccinol. 2016, 5, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Sawyer, L.A. Antibodies for the prevention and treatment of viral diseases. Antivir. Res. 2000, 47, 57–77. [Google Scholar] [CrossRef]
- Farrera-Soler, L.; Daguer, J.-P.; Barluenga, S.; Winssinger, N. Experimental Identification of Immuno-dominant B-cell Epitopes from SARS-CoV-2. Chimia 2021, 75, 276–284. [Google Scholar] [CrossRef]
- Wang, X.; Lam, J.-Y.; Chen, L.; Au, S.W.-N.; To, K.K.W.; Yuen, K.-Y.; Kok, K.-H. Mining of linear B cell epitopes of SARS-CoV-2 ORF8 protein from COVID-19 patients. Emerg. Microbes Infect. 2021, 10, 1016–1023. [Google Scholar] [CrossRef]
- Amrun, S.N.; Lee, C.Y.-P.; Lee, B.; Fong, S.-W.; Young, B.E.; Chee, R.S.-L.; Yeo, N.K.-W.; Torres-Ruesta, A.; Carissimo, G.; Poh, C.M.; et al. Linear B-cell epitopes in the spike and nucleocapsid proteins as markers of SARS-CoV-2 exposure and disease severity. EBioMedicine 2020, 58, 102911. [Google Scholar] [CrossRef]
- Li, Y.; Ma, M.; Lei, Q.; Wang, F.; Hong, W.; Lai, D.; Hou, H.; Xu, Z.; Zhang, B.; Chen, H.; et al. Linear epitope landscape of the SARS-CoV-2 Spike protein constructed from 1051 COVID-19 patients. Cell Rep. 2021, 34, 108915. [Google Scholar] [CrossRef]
- Schwarz, T.; Heiss, K.; Mahendran, Y.; Casilag, F.; Kurth, F.; Sander, L.E.; Wendtner, C.-M.; Hoechstetter, M.A.; Müller, M.A.; Sekul, R.; et al. SARS-CoV-2 Proteome-Wide Analysis Revealed Significant Epitope Signatures in COVID-19 Patients. Front. Immunol. 2021, 12, 629185. [Google Scholar] [CrossRef]
- Marshall, J.C.; Murthy, S.; Diaz, J.; Adhikari, N.K.; Angus, D.C.; Arabi, Y.M.; Baillie, K.; Bauer, M.; Berry, S.; Blackwood, B.; et al. A minimal common outcome measure set for COVID-19 clinical research. Lancet Infect. Dis. 2020, 20, e192–e197. [Google Scholar] [CrossRef]
- Jespersen, M.C.; Peters, B.; Nielsen, M.; Marcatili, P. BepiPred-2.0: Improving sequence-based B-cell epitope prediction using conformational epitopes. Nucleic Acids Res. 2017, 45, W24–W29. [Google Scholar] [CrossRef] [Green Version]
- Ponomarenko, J.; Bui, H.-H.; Li, W.; Fusseder, N.; Bourne, P.E.; Sette, A.; Peters, B. ElliPro: A new structure-based tool for the prediction of antibody epitopes. BMC Bioinform. 2008, 9, 514. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Ji, J.; Ye, J.; Zhao, X.; Wen, J.; Li, W.; Hu, J.; Li, D.; Sun, M.; Zeng, H.; et al. The Structure Analysis and Antigenicity Study of the N Protein of SARS-CoV. Genom. Proteom. Bioinform. 2003, 1, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Poh, C.M.; Carissimo, G.; Wang, B.; Amrun, S.N.; Lee, C.Y.-P.; Chee, R.S.-L.; Fong, S.-W.; Yeo, N.K.-W.; Lee, W.-H.; Torres-Ruesta, A.; et al. Two linear epitopes on the SARS-CoV-2 spike protein that elicit neutralising antibodies in COVID-19 patients. Nat. Commun. 2020, 11, 2806. [Google Scholar] [CrossRef]
- Zamecnik, C.R.; Rajan, J.V.; Yamauchi, K.A.; Mann, S.A.; Loudermilk, R.P.; Sowa, G.M.; Zorn, K.C.; Alvarenga, B.D.; Gaebler, C.; Caskey, M.; et al. ReScan, a Multiplex Diagnostic Pipeline, Pans Human Sera for SARS-CoV-2 Antigens. Cell Rep. Med. 2020, 1, 100123. [Google Scholar] [CrossRef] [PubMed]
- Shrock, E.; Fujimura, E.; Kula, T.; Timms, R.T.; Lee, I.-H.; Leng, Y.; Robinson, M.L.; Sie, B.M.; Li, M.Z.; Chen, Y.; et al. Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370, eabd4250. [Google Scholar] [CrossRef]
- Ladner, J.T.; Henson, S.N.; Boyle, A.S.; Engelbrektson, A.L.; Fink, Z.W.; Rahee, F.; D’ambrozio, J.; Schaecher, K.E.; Stone, M.; Dong, W.; et al. Epitope-resolved profiling of the SARS-CoV-2 antibody response identifies cross-reactivity with an endemic human CoV. Immunology 2020, 2, 100189. [Google Scholar]
- Yi, Z.; Ling, Y.; Zhang, X.; Chen, J.; Hu, K.; Wang, Y.; Song, W.; Ying, T.; Zhang, R.; Lu, H.; et al. Functional mapping of B-cell linear epitopes of SARS-CoV-2 in COVID-19 convalescent population. Emerg. Microbes Infect. 2020, 9, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Huang, X.; Joshi, S.; Guo, C.; Ng, J.; Thakkar, R.; Wu, Y.; Dong, X.; Li, Q.; Pinapati, R.; et al. Immunoreactive peptide maps of SARS-CoV-2 and other human coronaviruses. bioRxiv 2020. [Google Scholar] [CrossRef]
- Klompus, S.; Leviatan, S.; Vogl, T.; Kalka, I.N.; Godneva, A.; Shinar, E.; Weinberger, A.; Segal, E. Cross-reactive antibody responses against SARS-CoV-2 and seasonal common cold coronaviruses. medRxiv 2020. [Google Scholar] [CrossRef]
- Haynes, W.A.; Kamath, K.; Bozekowski, J.; Baum-Jones, E.; Campbell, M.; Casanovas-Massana, A.; Daugherty, P.S.; Dela Cruz, C.S.; Dhal, A.; Farhadian, S.F.; et al. High-resolution mapping and characterization of epitopes in COVID-19 patients. medRxiv 2020. [Google Scholar] [CrossRef]
- Holenya, P.; Lange, P.J.; Reimer, U.; Woltersdorf, W.; Panterodt, T.; Glas, M.; Wasner, M.; Eckey, M.; Drosch, M.; Hollidt, J.-M.; et al. Peptide microarray based analysis of antibody responses to SARS-CoV-2 identifies unique epitopes with potential for diagnostic test development. medRxiv 2020. [Google Scholar] [CrossRef]
- Heffron, A.S.; McIlwain, S.J.; Amjadi, M.F.; Baker, D.A.; Khullar, S.; Sethi, A.K.; Palmenberg, A.C.; Shelef, M.A.; O’Connor, D.H.; Ong, I.M. The landscape of antibody binding in SARS-CoV-2 infection. Immunology 2020, 19, e3001265. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, X.; Wang, D.; Lu, M.; Hou, X.; Wang, H.; Liang, T.; Dai, J.; Duan, H.; Xu, Y.; et al. Proteome-wide analysis of differentially-expressed SARS-CoV-2 antibodies in early COVID-19 infection. medRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Musicò, A.; Frigerio, R.; Mussida, A.; Barzon, L.; Sinigaglia, A.; Riccetti, S.; Gobbi, F.; Piubelli, C.; Bergamaschi, G.; Chiari, M.; et al. SARS-CoV-2 Epitope Mapping on Microarrays Highlights Strong Immune-Response to N Protein Region. Vaccines 2021, 9, 35. [Google Scholar] [CrossRef]
- Premkumar, L.; Segovia-Chumbez, B.; Jadi, R.; Martinez, D.R.; Raut, R.; Markmann, A.; Cornaby, C.; Bartelt, L.; Weiss, S.; Park, Y.; et al. The receptor binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV-2 patients. Sci. Immunol. 2020, 5, eabc8413. [Google Scholar] [CrossRef]
- Gallo Marin, B.; Aghagoli, G.; Lavine, K.; Yang, L.; Siff, E.J.; Chiang, S.S.; Salazar-Mather, T.P.; Dumenco, L.; Savaria, M.C.; Aung, S.N.; et al. Predictors of COVID-19 severity: A literature review. Rev. Med. Virol. 2021, 31, 1–10. [Google Scholar] [CrossRef]
- Chidambaram, V.; Tun, N.L.; Haque, W.Z.; Majella, M.G.; Sivakumar, R.K.; Kumar, A.; Hsu, A.T.-W.; Ishak, I.A.; Nur, A.A.; Ayeh, S.K.; et al. Factors associated with disease severity and mortality among patients with COVID-19: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0241541. [Google Scholar] [CrossRef]
- Williamson, E.J.; Walker, A.J.; Bhaskaran, K.; Bacon, S.; Bates, C.; Morton, C.E.; Curtis, H.J.; Mehrkar, A.; Evans, D.; Inglesby, P.; et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature 2020, 584, 430–436. [Google Scholar] [CrossRef]
- Carrillo, J.; Izquierdo-Useros, N.; Ávila-Nieto, C.; Pradenas, E.; Clotet, B.; Blanco, J. Humoral immune responses and neutralizing antibodies against SARS-CoV-2; implications in pathogenesis and protective immunity. Biochem. Biophys. Res. Commun. 2021, 538, 187–191. [Google Scholar] [CrossRef]
- Chan, J.F.-W.; Kok, K.-H.; Zhu, Z.; Chu, H.; To, K.K.-W.; Yuan, S.; Yuen, K.-Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, Y.; Li, Y.; Huang, F.; Luo, B.; Yuan, Y.; Xia, B.; Ma, X.; Yang, T.; Yu, F.; et al. The ORF8 protein of SARS-CoV-2 mediates immune evasion through down-regulating MHC-Ι. Proc. Natl. Acad. Sci. USA 2021, 118, e2024202118. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.M.; Fink, K.; Schmid, M.A.; Cathcart, A.; Spreafico, R.; Havenar-Daughton, C.; Lanzavecchia, A.; Corti, D.; Virgin, H.W. A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 2020, 584, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.S.; Wheatley, A.K.; Kent, S.J.; DeKosky, B.J. Antibody-dependent enhancement and SARS-CoV-2 vaccines and therapies. Nat. Microbiol. 2020, 5, 1185–1191. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Clinical Data | Disease Severity | |||
|---|---|---|---|---|
| Mild N (%) 84 (39) | Moderate N (%) 42 (20) | Severe N (%) 87 (41) | Total N (%) 213 (100) | |
| Median age (interquartile range) (years) | 65 (45.7–87.0) | 83.5 (58.5–89.5) | 69 (61.0–75.0) | 69.0 (55.0–84.0) |
| Male | 27 (32) | 15 (36) | 57 (66) | 99 (46) |
| Female | 57 (68) | 27 (64) | 30 (34) | 114 (54) |
| Cardiovascular disease | 21 (25) | 18 (43) | 37 (42) | 76 (35) |
| Hypertension | 29 (34) | 25 (60) | 55 (63) | 109 (51) |
| Diabetes mellitus | 17 (20) | 9 (21) | 18 (21) | 44 (21) |
| Lung disease | 10 (12) | 6 (14) | 17 (19) | 33 (16) |
| Renal insufficiency | 5 (6) | 7 (17) | 9 (10) | 21 (10) |
| Dialysis | 0 (0.0) | 0 (0) | 1 (1) | 1 (0.5) |
| Kidney transplantation | 0 (0.0) | 1 (2) | 4 (5) | 5 (2) |
| Liver failure | 4 (5) | 0 (0) | 1 (1) | 5 (2) |
| Obesity | 12 (14) | 10 (24) | 30 (34) | 52 (24) |
| Peptides of Interest | Disease Severity | ||
|---|---|---|---|
| Mild | Moderate | Severe | |
| SK27 | 0.015 (0.000–0.065) | 0.001 (0.000–0.1375) | 0.342 (0.073–0.851) |
| SK28 | 0.138 (0.043–0.293) | 0.124 (0.023–0.716) | 0.591 (0.161–1.48) |
| NC2 | 0.237 (0.065–0.458) | 0.273 (0.123–0.513) | 0.432 (0.235–0.865) |
| NC3′ | 0.068 (0.005–0.649) | 0.236 (0.037–2.101) | 0.749 (0.221–2.522) |
| NC5′ | 0.532 (0.169–1.097) | 0.662 (0.332–1.440) | 1.052 (0.502–2.298) |
| M1 | 0.000 (0.000–0.235) | 0.000 (0.000–0.085) | 0.130 (0.000–1.012) |
| NS2 | 0.512 (0.072–1.216) | 0.801 (0.476–1.319) | 0.811 (0.467–1.138) |
| Variables of Interest | Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|---|
| Crude OR | p-Value | OR | 95%CI | p-Value | |
| Clinical features | |||||
| Age > 50 years | 6.1 | <10−3 | 3.5 | 1.05–13.36 | 0.049 |
| Male | 3.8 | <10−3 | 2.7 | 1.21–6.08 | 0.02 |
| Cardiovascular diseases | 1.6 | 0.09 | - | - | - |
| Hypertension | 2.3 | 0.003 | 2.5 | 1.03–6.15 | 0.04 |
| Chronic lung diseases | 1.6 | 0.18 | - | - | - |
| Diabetes mellitus | 1.0 | 0.98 | - | - | - |
| Obesity | 2.6 | 0.01 | - | - | - |
| Kidney chronic diseases | 1.1 | 0.85 | - | - | - |
| Liver failure | 0.4 | 0.35 | - | - | - |
| Peptides | |||||
| SK27 > 0.05 | 7.4 | <10−3 | 4.1 | 1.80–9.83 | <10−3 |
| SK28 > 0.3 | 2.6 | <10−3 | - | - | - |
| NC2 > 0.1 | 4.2 | 0.001 | - | - | - |
| NC3′ > 0.17 | 4.9 | <10−3 | 3.9 | 1.70–9.22 | 0.002 |
| NC5′ > 0.35 | 3.4 | 0.001 | - | - | - |
| M1 > 0.9 | 9.1 | <10−3 | 6.9 | 1.94–28.37 | <10−3 |
| NS2 > 2 | 0.1 | 0.06 | 0.1 | 0.002–0.63 | 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrscher, C.; Eymieux, S.; Gaborit, C.; Blasco, H.; Marlet, J.; Stefic, K.; Roingeard, P.; Grammatico-Guillon, L.; Hourioux, C. ELISA-Based Analysis Reveals an Anti-SARS-CoV-2 Protein Immune Response Profile Associated with Disease Severity. J. Clin. Med. 2022, 11, 405. https://doi.org/10.3390/jcm11020405
Herrscher C, Eymieux S, Gaborit C, Blasco H, Marlet J, Stefic K, Roingeard P, Grammatico-Guillon L, Hourioux C. ELISA-Based Analysis Reveals an Anti-SARS-CoV-2 Protein Immune Response Profile Associated with Disease Severity. Journal of Clinical Medicine. 2022; 11(2):405. https://doi.org/10.3390/jcm11020405
Chicago/Turabian StyleHerrscher, Charline, Sébastien Eymieux, Christophe Gaborit, Hélène Blasco, Julien Marlet, Karl Stefic, Philippe Roingeard, Leslie Grammatico-Guillon, and Christophe Hourioux. 2022. "ELISA-Based Analysis Reveals an Anti-SARS-CoV-2 Protein Immune Response Profile Associated with Disease Severity" Journal of Clinical Medicine 11, no. 2: 405. https://doi.org/10.3390/jcm11020405