The Effect of Osmopriming on Seed Germination and Early Seedling Characteristics of Carum carvi L.

 ,
, 
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Condition
2.2. Priming Experiment in Petri Dishes
2.3. Priming Experiment in Soil
2.4. Statistics
3. Results and Discussion
3.1. Priming Experiment in Petri Dishes
3.2. PEG Priming Experiment in Soil
3.3. Hydro-priming Experiment in Soil
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ruszkowska, J. Main chemical constituents of Carum. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 38–60. [Google Scholar]
- Sadowska, A.; Obidoska, G. Pharmacological uses and toxicology of caraway. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 186–196. [Google Scholar]
- Toxopeus, H.; Lubberts, H. A century of breeding caraway in the Netherlands. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 117–143. [Google Scholar]
- Samojlik, I.; Lakic, N.; Mimica-Dukic, N.; Đaković-S#x160;vajcer, K.; Bozin, B. Antioxidant and hepatoprotective potential of essential oils of coriander (Coriandrum sativum L.) and caraway (Carum carvi L.)(Apiaceae). J. Agr. Food Chem. 2010, 58, 8848–8853. [Google Scholar] [CrossRef] [PubMed]
- Javed, R.; Hanif, M.A.; Rehman, R.; Hanif, M.; Tung, B.T. Caraway. In Medicinal Plants of South Asia; Asif Hanif, M., Mumtaz Khan, M., Nawaz, H.J., Byrne, H., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 87–100. [Google Scholar]
- Bouwmeester, H.H. Regulation of essential oil formation of caraway. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 92–116. [Google Scholar]
- Németh, É. Questions of the generative development in caraway. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 79–91. [Google Scholar]
- Omidbaigi, R. Production and Processing of Medicinal Plants II; Astan Quds Publication: Tehran, Iran, 2005; p. 99. ISBN 964-02-395-5. (In Farsi) [Google Scholar]
- Weglarz, Z. Production of biennial caraway for seed and essential oil. In Caraway -The Genus Carum; Németh, É., Ed.; Harwood Academic Publisher: Amsterdam, The Netherlands, 1998; pp. 144–157. [Google Scholar]
- Tzortzakis, N.G. Effect of pre-sowing treatment on seed germination and seedling vigour in endive and chicory. Horti. Sci. (Prague) 2009, 36, 117–125. [Google Scholar] [CrossRef]
- Galambosi, B.; Peura, P. Agrobotanical features and oil content of wild and cultivated forms of caraway (Carum carvi L.). J. Essent. Oil Res. 1996, 8, 389–397. [Google Scholar] [CrossRef]
- Bewleyl, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Heydecker, W.; Coolbear, P. Seed treatment for improved performance survey and attempted prognosis. Seed Sci. Technol. 1977, 13, 299–335. [Google Scholar]
- Bradford, K.J. Manipulation of seed water relations via osmotic priming to improve germination under stress condition. HortScience (USA) 1989, 21, 1105–1112. [Google Scholar]
- Khan, A.A. Preplant physiological seed conditioning. Hort. Rev. 1992, 13, 131–172. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Pre-sowing seed treatment –A shotgun approach to improve germination, plant growth, and crop yield under saline and non-saline conditions. Adv. Agron. 2005, 88, 223–271. [Google Scholar]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef] [PubMed]
- Ehrenshaft, M.; Brambl, R. Respiration and mitochondrial biogenesis in germinating embryos of maize. Plant Physiol. 1990, 93, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Attucci, S.; Carde, J.P.; Raymond, P.; Saint Gès, V.; Spiteri, A.; Pradet, A. Oxidative phosphorylation by mitochondria extracted from dry sunflower seeds. Plant Physiol. 1991, 95, 390–398. [Google Scholar] [CrossRef] [PubMed]
- Thornton, J.M.; Collins, A.R.S.; Powell, A.A. The effect of aerated hydration on DNA synthesis in embryos of Brassica oleracea L. Seed Sci. Res. 1993, 3, 195–199. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Rehman, H.; Saleem, B.A. Seed priming enhances the performance of late sown wheat (Triticum aestivum L.) by improving chilling tolerance. J. Agron. Crop Sci. 2008, 194, 55–60. [Google Scholar] [CrossRef]
- Giri, G.S.; Schillinger, W.F. Seed priming winter wheat for germination, emergence and yield. Crop Sci. 2003, 43, 2135–2141. [Google Scholar] [CrossRef]
- Mazor, L.; Perl, M.; Negbi, M. Changes in some ATP-dependent activities in seeds during treatment with polyethylene glycol and during the redrying process. J. Exp. Bot. 1984, 35, 1119–1127. [Google Scholar] [CrossRef]
- Bujalski, W.; Nienow, A.W. Large-scale osmotic priming of onion seeds: A comparison of different strategies for oxygenation. Sci. Hortic. 1991, 46, 13–24. [Google Scholar] [CrossRef]
- Michel, B.E.; Kaufmann, M.R. The osmotic potential of polyethylene glycol 6000. Plant Physiol. 1973, 51, 914–916. [Google Scholar] [CrossRef]
- Demir, I.; Mavi, K. Effect of salt and osmotic stresses on the germination of Pepper seeds of different maturation stages. Braz. Arc. Biol. Techn. 2008, 51, 897–902. [Google Scholar] [CrossRef]
- Maguire, J.D. Speed of germination-aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 1962, 2, 176–177. [Google Scholar] [CrossRef]
- IBM Corp. IBM SPSS Statistics for Windows, Version 25.0; IBM Corp: Armonk, NY, USA, 2017. [Google Scholar]
- Roberts, E.; Smith, R.D. Dormancy and the pentose phosphate pathway. In The Physiology and Biochemistry of Seed Dormancy and Germination; Khan, A., Ed.; North-Holland Publishing Co: Amsterdam, The Netherlands, 1977; pp. 385–411. [Google Scholar]
- Pérez-Fernández, M.A.; Calvo-Magro, E.; Montanero-Fernández, J.; Oyola-elasco, J.A. Seed germination in response to chemicals: Effect of nitrogen and pH in the media. J. Environ. Biol. 2006, 27, 13–20. [Google Scholar]
- Blandino, C.; Fernández-Pascual, E.; Marin, M.; Vernet, A.; Pritchard, H.W. Seed ecology of the geophyte Conopodium majus (Apiaceae), indicator species of ancient woodland understories and oligotrophic meadows. Plant Biol. 2019, 21, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Tahaei, A.; Soleymani, A.; Shams, M. Seed germination of medicinal plant, fennel (Foeniculum vulgare Mill), as affected by different priming techniques. Appl. Biochem. Biotech. 2016, 180, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Ahmadvand, G.; Soleimani, F.; Saadatian, B.; Pouya, M. Effect of seed priming with potassium nitrate on germination and emergence traits of two soybean cultivars under salinity stress conditions. Am. Eurasian J. Agric. Environ. Sci. 2012, 12, 769–774. [Google Scholar]
- Espanany, A.; Fallah, S.; Tadayyon, A. Seed priming improves seed germination and reduces oxidative stress in black cumin (Nigella sativa) in presence of cadmium. Ind. Crops Prod. 2016, 79, 195–204. [Google Scholar] [CrossRef]
- Kumar, S.; Hemantaranjan, A.; Mondal, S.; Bose, B. Impact of KNO3 Primed seeds on the performance of late sown sesame (Sesamum indicum L.). IJBSM 2016, 7, 950–954. [Google Scholar] [CrossRef]
- Patil, K.; Ravat Anilkumar, L.; Trivedi, V.; Hirpara, A.; Sasidharan, N. Effect of seed priming treatment in chickpea (Cicer arietinum L.). IJCS 2018, 6, 1064–1069. [Google Scholar]
- Alizadeh, M.A.; Sajjadi Jaghargh, S.S.; Sharifi, R.S.; Calagari, M.; Sedghi, M. Effect of seed priming and moist chilling on emergence traits of six populations (Anthemis haussknechtii Boiss. & Reut. and Anthemis pseudocotula Boiss.) in greenhouse condition. J. Med. Plants Prod. 2019, 8, 41–51. [Google Scholar]
- Damalas, C.A.; Koutroubas, S.D.; Fotiadis, S. Hydro-priming effects on seed germination and field performance of faba bean in spring sowing. Agriculture 2019, 9, 201. [Google Scholar] [CrossRef]
- Karssen, C.M.; Haigh, A.; Toorn, P.; Eges, R. Physiological mechanisms involved in seed priming. In Recent Advances in the Development and Germination of Seeds; Taylorson, R.B., Ed.; Plenum Press: New York, NY, USA; NLondon, UK, 1990; pp. 269–280. [Google Scholar]
- Gray, D.; Steckel, J.R.A.; Hands, L.J. Responses of vegetable seeds to controlled hydration. Ann. Bot. 1990, 66, 227–235. [Google Scholar] [CrossRef]
- Vandelook, F.; Van Assche, J.A. Deep complex morphophysiological dormancy in Sanicula europaea (Apiaceae) fits a recurring pattern of dormancy types in genera with an Arcto-Tertiary distribution. Botany 2008, 86, 1370–1377. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; de Casas, R.R.; Bradford, K.; Burghardt, L.; et al. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef] [PubMed]



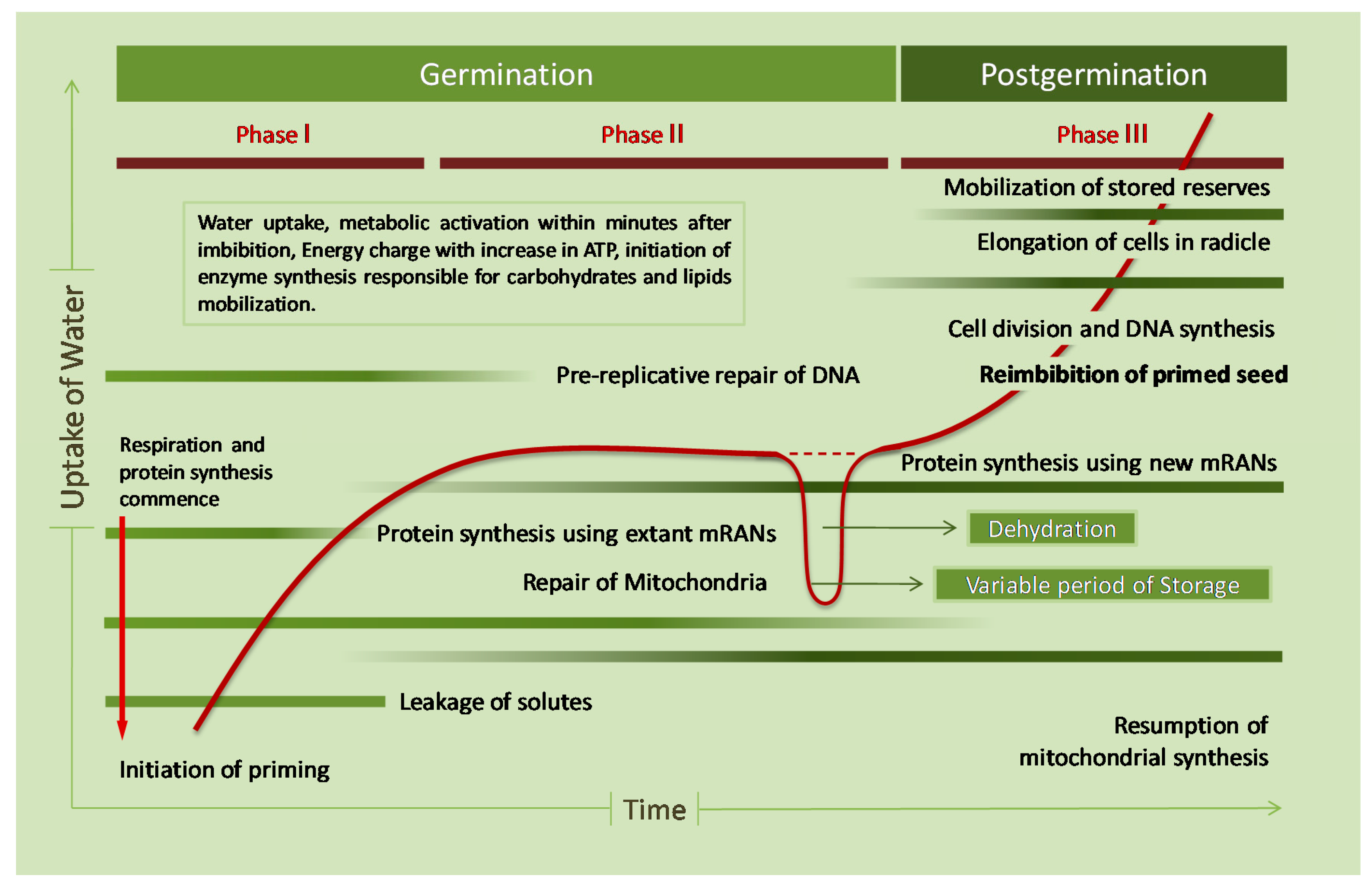{kind=link}
{kind=link}
| Treatment | Time | Germination percent | Germination rate (GR) | Seedling length (cm) | Plumule fresh weight (mg) | Plumule dry weight (mg) |
|---|---|---|---|---|---|---|
| PEG 5% | 12 h | 54.00 ± 19.73 | 1.64 ± 0.27 | 8.95±0.89 | 29.14 ± 5.45 | 1.62 ± 0.06 |
| 24 h | 46.00 ± 14.79 | 1.41 ± 0.43 | 7.48 ± 0.96A | 23.98 ± 1.64 | 1.92 ± 0.06 | |
| 36 h | 51.00 ± 2.00 | 1.44 ± 0.09 | 6.58 ± 0.87 | 24.67 ± 3.40 | 1.93 ± 0.07 | |
| PEG* 10% | 12 h | 53.00 ± 15.10 | 1.62 ± 0.40 | 7.90 ± 0.35B | 25.70 ± 1.36 | 1.79 ± 0.07 |
| 24 h | 51.00 ± 11.94 | 1.50 ± 0.24 | 7.96 ± 0.48AB | 24.49 ± 4.39 | 1.86 ± 0.15B | |
| 36 h | 61.00 ± 11.94 | 1.71 ± 0.31 | 7.40 ± 0.28A | 23.43 ± 2.72 | 2.00 ± 0.10B | |
| PEG 20% | 12 h | 52.00 ± 8.64 | 1.46 ± 0.28 | 7.86 ± 0.65A | 21.37 ± 5.26 | 1.92 ± 0.20 |
| 24 h | 54.00 ± 6.93 | 1.62 ± 0.11 | 7.61 ± 0.51 | 25.80 ± 4.04 | 1.70 ± 0.12B | |
| 36 h | 48.00 ± 13.47 | 1.42 ± 0.34 | 6.66 ± 0.59A | 26.43 ± 3.46 | 2.01 ± 0.12B | |
| KNO3* 0.5% | 12 h | 52.00 ± 9.80 | 1.61 ±0.27 | 9.11 ± 0.26 | 20.70 ± 2.83 | 1.78 ± 0.27 |
| 24 h | 55.00 ± 10.52 | 1.61 ±0.36 | 8.93 ± 0.28B | 26.20 ± 5.80 | 2.02 ± 0.31 | |
| 36 h | 48.00 ± 16.97 | 1.43 ±0.42 | 7.77 ± 0.47a | 27.31 ± 6.24 | 2.08 ± 0.22 | |
| KNO3 1% | 12 h | 48.00 ± 9.80 | 1.47 ± 0.22 | 9.47 ± 0.40B | 25.40 ± 1.87 | 1.92 ± 0.18 |
| 24 h | 39.00 ± 8.25 | 1.38 ± 0.19 | 9.03 ± 0.66B | 24.18 ± 3.44 | 2.02 ± 0.21B | |
| 36 h | 51.00 ± 8.25 | 1.61 ± 0.22 | 9.48 ± 0.53bB | 24.00 ± 3.52 | 2.01 ± 0.20AB | |
| KNO3 2% | 12 h | 47.00 ± 13.22 | 1.42 ± 0.36 | 9.96 ± 0.76B | 24.55 ± 2.59 | 2.04 ± 0.19 |
| 24 h | 57.00 ± 16.77 | 1.71 ± 0.35 | 9.08 ± 0.96 | 27.13 ± 0.94 | 2.13 ± 0.06B | |
| 36 h | 49.00 ± 10.52 | 1.52 ± 0.31 | 9.12 ± 0.28bB | 30.13 ± 2.53 | 2.11 ± 0.14A | |
| KCL 1% | 12 h | 54.00 ± 17.74 | 1.71 ± 0.52 | 8.32 ± 0.66b | 25.93 ± 3.47 | 2.10 ± 0.12 |
| 24 h | 48.00 ± 7.12 | 1.43 ± 0.37 | 8.15 ± 0.32bAB | 31.24 ± 2.48 | 1.86 ± 0.18 | |
| 36 h | 51.00 ± 8.25 | 1.53 ± 0.28 | 7.98 ± 0.68 | 26.94 ± 2.68 | 2.05 ± 0.10 | |
| KCL* 2% | 12 h | 40.0 ± 11.78 | 1.31 ± 0.18 | 8.31 ± 0.28bA | 27.11 ± 1.08 | 1.68 ± 0.07 |
| 24 h | 52.00 ± 10.83 | 1.56 ± 0.36 | 6.73 ± 0.83aA | 23.11 ± 2.36 | 1.80 ± 0.13A | |
| 36 h | 46.00 ± 7.66 | 1.46 ± 0.24 | 7.23 ± 0.15A | 24.88 ± 1.22 | 1.46 ± 0.08A | |
| KCL 4% | 12 h | 55.00 ± 3.83 | 1.72 ± 0.07 | 7.77 ± 0.16aA | 27.71 ± 4.59 | 1.66 ± 0.21 |
| 24 h | 48.00 ± 6.53 | 1.46 ± 0.20 | 7.66 ± 0.66b | 26.90 ± 2.45 | 1.82 ± 0.07A | |
| 36 h | 46.00 ± 13.27 | 1.46 ± 0.38 | 8.01 ± 1.00 AB | 29.84 ± 3.66 | 1.72 ± 0.04B | |
| Control | 46.00 ± 10.58 | 1.42 ± 0.34 | 6.66 ± 0.32 | 24.48 ± 3.73 | 1.62 ± 0.19 |
| Treatment | Time | Germination percent | Germination rate | Plumule length (cm) | Plumule fresh weight (mg) | Plumule dry weight (mg) |
|---|---|---|---|---|---|---|
| PEG 5% | 12 h | 56.63 ± 3.35B | 1.61 ± 0.03B | 5.89 ± 0.27 | 94 ± 0.006b | 12.2 ± 0.1 |
| 24 h | 63.30 ± 3.30B | 1.91 ± 0.03C | 6.17 ± 0.09 | 105 ± 0.007b | 11.1 ± 0.4 | |
| 36 h | 33.30 ± 3.30A | 0.84 ± 0.05A | 6.01 ± 0.31 | 074 ± 0.006a | 9.8 ± 0.12 | |
| PEG 10% | 12 h | 36.63 ± 3.35A | 1.44 ± 0.04A | 6.08 ± 0.52 | 92 ± 0.001b | 11.2 ± 0.2 |
| 24 h | 56.63 ± 3.35B | 1.59 ± 0.02B | 5.78 ± 0.54 | 79 ± 0.006a | 10.8 ± 0.3 | |
| 36 h | 43.30 ± 3.30A | 1.39 ± 0.04A | 5.94 ± 0.72 | 101 ± 0.004b | 13.8 ± 0.12 | |
| PEG 20% | 12 h | 49.97 ± 3.35A | 1.36 ± 0.02A | 6.19 ± 0.29 | 64 ± 0.001a | 9.5 ± 0.7 |
| 24 h | 53.30 ± 3.30A | 1.49 ± 0.02B | 5.31 ± 0.23 | 81 ± 0.008a | 11.0 ± 0.12 | |
| 36 h | 49.97 ± 3.35A | 1.55 ± 0.05B | 5.62 ± 0.23 | 80 ± 0.011ab | 10.8 ± 0.7 | |
| Control | 39.97 ± 3.35 | 1.01 ± 0.02 | 4.49 ± 0.05 | 53 ± 0.1 | 7.0 ± 0.04 |
| Treatment | Time | Germination percent | Germination rate | Plumule length (cm) | Plumule fresh weight (mg) | Plumule dry weight (mg) |
|---|---|---|---|---|---|---|
| H2O | 12 h | 53.30 ± 3.35* | 1.62 ± 0.01*** | 5.75 ± 0.12*** | 85.66 ± 0.9** | 12.1 ± 0.05*** |
| 24 h | 53.30 ± 3.30* | 1.52 ± 0.02*** | 5.61 ± 0.42 | 81.33 ± 0.05*** | 11.0 ± 0.06** | |
| 36 h | 46.63 ± 3.30 | 1.50 ± 0.03*** | 4.73 ± 0.37 | 75.33 ± 0.57** | 9.8 ± 0.08* | |
| Control | 39.97 ± 3.35 | 1.01±0.02 | 4.49 ± 0.05 | 53.0 ± 0.1 | 7.0 ± 0.04 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirmazloum, I.; Kiss, A.; Erdélyi, É.; Ladányi, M.; Németh, É.Z.; Radácsi, P. The Effect of Osmopriming on Seed Germination and Early Seedling Characteristics of Carum carvi L. Agriculture 2020, 10, 94. https://doi.org/10.3390/agriculture10040094
Mirmazloum I, Kiss A, Erdélyi É, Ladányi M, Németh ÉZ, Radácsi P. The Effect of Osmopriming on Seed Germination and Early Seedling Characteristics of Carum carvi L. Agriculture. 2020; 10(4):94. https://doi.org/10.3390/agriculture10040094
Chicago/Turabian StyleMirmazloum, Iman, Attila Kiss, Éva Erdélyi, Márta Ladányi, Éva Zámboriné Németh, and Péter Radácsi. 2020. "The Effect of Osmopriming on Seed Germination and Early Seedling Characteristics of Carum carvi L." Agriculture 10, no. 4: 94. https://doi.org/10.3390/agriculture10040094
APA StyleMirmazloum, I., Kiss, A., Erdélyi, É., Ladányi, M., Németh, É. Z., & Radácsi, P. (2020). The Effect of Osmopriming on Seed Germination and Early Seedling Characteristics of Carum carvi L. Agriculture, 10(4), 94. https://doi.org/10.3390/agriculture10040094