

Identification of Allelochemicals with Differential Modes of Phytotoxicity against Cuscuta campestris
 , ,
, ,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Chemicals
2.2. In Vitro Experiments for Screening of Allelopathy against Cuscuta Seedling Growth
2.3. Calculations and Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimentel, D.; Zuniga, R.; Morrison, D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol. Econ. 2005, 52, 273–288. [Google Scholar] [CrossRef]
- Parker, C.; Riches, C.R. Parasitic Weeds of the World: Biology and Control; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Fernández-Aparicio, M.; Flores, F.; Rubiales, D. The effect of Orobanche crenata infection severity in faba bean, field pea and grass pea productivity. Front. Plant Sci. 2016, 7, 1049. [Google Scholar] [CrossRef] [Green Version]
- Goldwasser, Y.; Miryamchik, H.; Sibony, M.; Rubin, B. Detection of resistant chickpea (Cicer arietinum) genotypes to Cuscuta campestris (field dodder). Weed Res. 2012, 52, 122–130. [Google Scholar] [CrossRef]
- Córdoba, E.M.; Fernández-Aparicio, M.; González-Verdejo, C.I.; López-Grau, C.; Muñoz-Muñoz, M.V.; Nadal, S. Search for Resistant Genotypes to Cuscuta campestris Infection in two legume species, Vicia sativa and Vicia ervilia. Plants 2021, 10, 738. [Google Scholar] [CrossRef]
- Kaštier, P.; Krasylenko, Y.A.; Martincová, M.; Panteris, E.; Šamaj, J.; Blehová, A. Cytoskeleton in the parasitic plant Cuscuta during germination and pre-haustorium formation. Front. Plant Sci. 2018, 9, 794. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Aparicio, M.; Delavault, P.; Timko, M.P. Management of infection by parasitic weeds: A Review. Plants 2020, 9, 1184. [Google Scholar] [CrossRef]
- Haidar, M.A.; Orr, G.L.; Westra, P. Effects of light and mechanical stimulation on coiling and pre-haustoria formation in Cuscuta spp. Weed Res. 1997, 37, 219–228. [Google Scholar] [CrossRef]
- Vaughn, K.C. Attachment of the parasitic weed dodder to the host. Protoplasma 2002, 219, 227–237. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Soriano, G.; Masi, M.; Carretero, P.; Vilariño-Rodríguez, S.; Cimmino, A. (4Z)-Lachnophyllum lactone, an acetylenic furanone from Conyza bonariensis, identified for the first time with allelopathic activity against Cuscuta campestris. Agriculture 2022, 12, 790. [Google Scholar] [CrossRef]
- Nadler-Hassar, T.; Shaner, D.L.; Nissen, S.; Westra, P.; Rubin, B. Are herbicide-resistant crops the answer to controlling Cuscuta? Pest Manag. Sci. 2009, 65, 811–816. [Google Scholar] [CrossRef]
- Dayan, F.E.; Duke, S.O. Natural compounds as next-generation herbicides. Plant Physiol. 2014, 166, 1090–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Charudattan, R.; Duke, S.O.; Fennimore, S.A.; Marrone, P.; Slaughter, D.C.; Swanton, C.; Zollinger, R. Weed management in 2050: Perspectives on the future of weed science. Weed Sci. 2018, 66, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Khanh, T.D.; Cong, L.C.; Xuan, T.D.; Lee, S.J.; Kong, D.S.; Chung, I.M. Weed-suppressing potential of dodder (Cuscuta hygrophilae) and its phytotoxic constituents. Weed Sci. 2008, 56, 119–127. [Google Scholar] [CrossRef]
- Azimi, A.A.; Hashemloian, B.D. Allelopathy and anti-mitotic effects of Cuscuta campestris and Cuscuta monogyna extracts on plant cell division. J. Med. Plants By-Prod. 2017, 6, 131–138. [Google Scholar]
- Zermane, N.; Vurro, M.; Boari, A.; Avolio, F.; Andolfi, A.; Evidente, A. Towards broomrape and field dodder management using natural metabolites from plants. In Proceedings of the Abstract of 11th World Congress on Parasitic Plants, Martina Franca, Italy, 7–12 June 2011; p. 82. [Google Scholar]
- Andolfi, A.; Zermane, N.; Cimmino, A.; Avolio, F.; Boari, A.; Vurro, M.; Evidente, A. Inuloxins A–D, phytotoxic bi-and tri-cyclic sesquiterpene lactones produced by Inula viscosa: Potential for broomrapes and field dodder management. Phytochemistry 2013, 86, 112–120. [Google Scholar] [CrossRef]
- Vurro, M.; Boari, A.; Evidente, A.; Andolfi, A.; Zermane, N. Natural metabolites for parasitic weed management. Pest Manag. Sci. 2009, 65, 566–571. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products—A Biosynthetic Approach; Wiley and Sons Ltd.: Chicester, UK, 2009. [Google Scholar]
- Evidente, A.; Cimmino, A.; Fernandez-Aparicio, M.; Andolfi, A.; Rubiales, D.; Motta, A. Polyphenols, including the new peapolyphenols A–C, from pea root exudates stimulate Orobanche foetida seed germination. J. Agric. Food Chem. 2010, 58, 2902–2907. [Google Scholar] [CrossRef] [Green Version]
- Gaertner, E.E. Studies of seed germination, seed identification, and host relationship in Dodders, Cuscuta spp. Mem. Cornell Agric. Exp. Stn. 1950, 294, 1–56. [Google Scholar]
- Cala, A.; Zorrilla, J.G.; Rial, C.; Molinillo, J.M.G.; Varela, R.M.; Macías, F.A. Easy access to alkoxy, amino, carbamoyl, hydroxy, and thiol derivatives of sesquiterpene lactones and evaluation of their bioactivity on parasitic weeds. J. Agric. Food Chem. 2019, 67, 10764–10773. [Google Scholar] [CrossRef]
- Zorrilla, J.G.; Rial, C.; Varela, R.M.; Molinillo, J.M.; Macías, F.A. Strategies for the synthesis of canonical, non-canonical and analogues of strigolactones, and evaluation of their parasitic weed germination activity. Phytochem. Rev. 2022, 21, 1627–1659. [Google Scholar] [CrossRef]
- Soriano, G.; Siciliano, A.; Fernández-Aparicio, M.; Cala Peralta, A.; Masi, M.; Moreno-Robles, A.; Guida, M.; Cimmino, A. Iridoid glycosides isolated from Bellardia trixago identified as inhibitors of Orobanche cumana radicle growth. Toxins 2022, 14, 559. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Aparicio, M.; Bernard, A.; Falchetto, L.; Marget, P.; Chauvel, B.; Steinberg, C.; Morris, C.E.; Gibot-Leclerc, S.; Boari, A.; Vurro, M.; et al. Investigation of amino acids as herbicides for control of Orobanche minor parasitism in red clover. Front. Plant Sci. 2017, 8, 842. [Google Scholar] [CrossRef]
- Kuruma, M.; Suzuki, T.; Seto, Y. Tryptophan derivatives regulate the seed germination and radicle growth of a root parasitic plant, Orobanche minor. Bioorg. Med. Chem. Lett. 2021, 43, 128085. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Cimmino, A.; Evidente, A.; Rubiales, D. Inhibition of Orobanche crenata seed germination and radicle growth by allelochemicals identified in cereals. J. Agric. Food Chem. 2013, 61, 9797–9803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanardo, D.I.L.; Lima, R.B.; Ferrarese, M.D.L.L.; Bubna, G.A.; Ferrarese-Filho, O. Soybean root growth inhibition and lignification induced by p-coumaric acid. Environ. Exp. Bot. 2009, 66, 25–30. [Google Scholar] [CrossRef]
- Bharathalakshmi; Jayachandra. Presowing hardening of the host with phenolic acids reduces induction of seed germination in the root parasite Striga asiatica. Int. J. Pest Manag. 1980, 26, 309–312. [Google Scholar]
- Huffman, C.W.; Godar, E.M.; Torgeson, D.C. Inhibition of plant growth by halogenated benzoic acids. J. Agric. Food Chem. 1967, 15, 976–981. [Google Scholar] [CrossRef]
- Jeschke, P. The unique role of halogen substituents in the design of modern agrochemicals. Pest Manag. Sci. 2010, 66, 10–27. [Google Scholar] [CrossRef] [PubMed]
- Macias, F.A.; Chinchilla, N.; Arroyo, E.; Molinillo, J.M.G.; Marin, D.; Varela, R.M. Combined strategy for phytotoxicity enhancement of benzoxazinones. J. Agric. Food Chem. 2010, 58, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Mejías, F.J.; Durán, A.G.; Zorrilla, J.G.; Varela, R.M.; Molinillo, J.M.G.; Valdivia, M.M.; Macías, F.A. Acyl derivatives of eudesmanolides to boost their bioactivity: An explanation of behavior in the cell membrane using a molecular dynamics approach. ChemMedChem 2021, 16, 1297–1307. [Google Scholar] [CrossRef]
- Borgati, T.F.; Alves, R.B.; Teixeira, R.R.; Freitas, R.P.D.; Perdigão, T.G.; Silva, S.F.D.; dos Santos, A.A.; Bastidas, A.D.J.O. Synthesis and phytotoxic activity of 1, 2, 3-triazole derivatives. J. Braz. Chem. Soc. 2013, 24, 953–961. [Google Scholar] [CrossRef]
- Yoneyama, K.; Takeuchi, Y.; Ogasawara, M.; Konnai, M.; Sugimoto, Y.; Sassa, T. Cotylenins and fusicoccins stimulate seed germination of Striga hermonthica (Del.) Benth and Orobanche minor Smith. J. Agric. Food Chem. 1998, 46, 1583–1586. [Google Scholar] [CrossRef]
- Bar Nun, N.; Mayer, A.M. Smoke chemicals and coumarin promote the germination of the parasitic weed Orobanche aegyptiaca. Isr. J. Plant Sci. 2005, 53, 97–101. [Google Scholar] [CrossRef]
- Serghini, K.; de Luque, A.P.; Castejón-Muñoz, M.; García-Torres, L.; Jorrín, J.V. Sunflower (Helianthus annuus L.) response to broomrape (Orobanche cernua Loefl.) parasitism: Induced synthesis and excretion of 7-hydroxylated simple coumarins. J. Exp. Bot. 2001, 52, 2227–2234. [Google Scholar] [CrossRef]
- Zorrilla, J.G.; Cala, A.; Rial, C.R.; Mejías, F.J.; Molinillo, J.M.; Varela, R.M.; Macías, F.A. Synthesis of active strigolactone analogues based on eudesmane-and guaiane-type sesquiterpene lactones. J. Agric. Food Chem. 2020, 68, 9636–9645. [Google Scholar] [CrossRef]
- Wada, S.; Cui, S.; Yoshida, S. Reactive oxygen species (ROS) generation is indispensable for haustorium formation of the root parasitic plant Striga hermonthica. Front. Plant Sci. 2019, 10, 328. [Google Scholar] [CrossRef] [Green Version]
- Cui, S.; Wada, S.; Tobimatsu, Y.; Takeda, Y.; Saucet, S.B.; Takano, T.; Yoshida, S. Host lignin composition affects haustorium induction in the parasitic plants Phtheirospermum japonicum and Striga hermonthica. New Phytol. 2018, 218, 710–723. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.S.; Barbosa, L.C.D.A.; de Alvarenga, E.S.; Demuner, A.J.; da Silva, A.A. Synthesis and phytotoxicity evaluation of substituted para-benzoquinones. Aust. J. Chem. 2003, 56, 625–630. [Google Scholar] [CrossRef]
- González-Ibarra, M.; Farfán, N.; Trejo, C.; Uribe, S.; Lotina-Hennsen, B. Selective herbicide activity of 2, 5-di (benzylamine)-p-benzoquinone against the monocot weed Echinochloa crusgalli. An in vivo analysis of photosynthesis and growth. J. Agric. Food Chem. 2005, 53, 3415–3420. [Google Scholar] [CrossRef]
- Dayan, F.E.; Howell, J.L.; Weidenhamer, J.D. Dynamic root exudation of sorgoleone and its in planta mechanism of action. J. Exp. Bot. 2009, 60, 2107–2117. [Google Scholar] [CrossRef] [Green Version]
- Zorrilla, J.G.; Rial, C.; Varela, R.M.; Molinillo, J.M.; Macías, F.A. Facile synthesis of anhydrojudaicin and 11, 13-dehydroanhydrojudaicin, two eudesmanolide-skeleton lactones with potential allelopathic activity. Phytochem. Lett. 2019, 31, 229–236. [Google Scholar] [CrossRef]
- Yoder, J.I. Host-plant recognition by parasitic Scrophulariaceae. Curr. Opin. Plant Biol. 2001, 4, 359–365. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Masi, M.; Cimmino, A.; Evidente, A. Effects of benzoquinones on radicles of Orobanche and Phelipanche species. Plants 2021, 10, 746. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, Y.; Simier, P.; Arfaoui, A.; Sifi, B.; Delavault, P.; Zourgui, L.; Belhadj, O. Induction of phenolic compounds in pea (Pisum sativum L.) inoculated by Rhizobium leguminosarum and infected with Orobanche crenata. J. Phytopathol. 2007, 155, 728–734. [Google Scholar] [CrossRef]
- Fernández-Aparicio, M.; Masi, M.; Cimmino, A.; Vilariño, S.; Evidente, A. Allelopathic effect of quercetin, a flavonoid from Fagopyrum esculentum roots in the radicle growth of Phelipanche ramosa: Quercetin natural and semisynthetic analogues were used for a structure-activity relationship investigation. Plants 2021, 10, 543. [Google Scholar] [CrossRef] [PubMed]
- Dear, J.; Aronoff, S. Relative kinetics of chlorogenic and caffeic acids during the onset of boron deficiency in sunflower. Plant Physiol. 1965, 40, 458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yordi, E.G.; Pérez, E.M.; Matos, M.J.; Villares, E.U. Antioxidant and pro-oxidant effects of polyphenolic compounds and structure-activity relationship evidence. Nutr. Well-Being Health 2012, 2, 23–48. [Google Scholar]
- Dinkel, D.H. Chlorogenic acid associated with physiological internal necrosis of potato tubers. Am. Potato J. 1963, 40, 149–153. [Google Scholar] [CrossRef]
- Krumsri, R.; Iwasaki, A.; Suenaga, K.; Kato-Noguch, H. Phytotoxic effects of Senna garrettiana and identification of phytotoxic substances for the development of bioherbicides. Agriculture 2022, 12, 1338. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Calomme, M.; Pieters, L.; Vlietinck, A.J.; Berghe, D.V. Structure-activity relationship of flavonoids as antioxidant and pro-oxidant compounds. Stud. Nat. Prod. Chem. 2000, 22, 307–341. [Google Scholar]
- Sunohara, Y.; Nakano, K.; Matsuyama, S.; Oka, T.; Matsumoto, H. Cuminaldehyde, a cumin seed volatile component, induces growth inhibition, overproduction of reactive oxygen species and cell cycle arrest in onion roots. Sci. Hortic. 2021, 289, 110493. [Google Scholar] [CrossRef]
- Khamphio, M.; Barusrux, S.; Weerapreeyakul, N. Sesamol induces mitochondrial apoptosis pathway in HCT116 human colon cancer cells via pro-oxidant effect. Life Sci. 2016, 158, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Kato-Noguchi, H. Isolation and identification of an allelopathic substance in Pisum sativum. Phytochemistry 2003, 62, 1141–1144. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
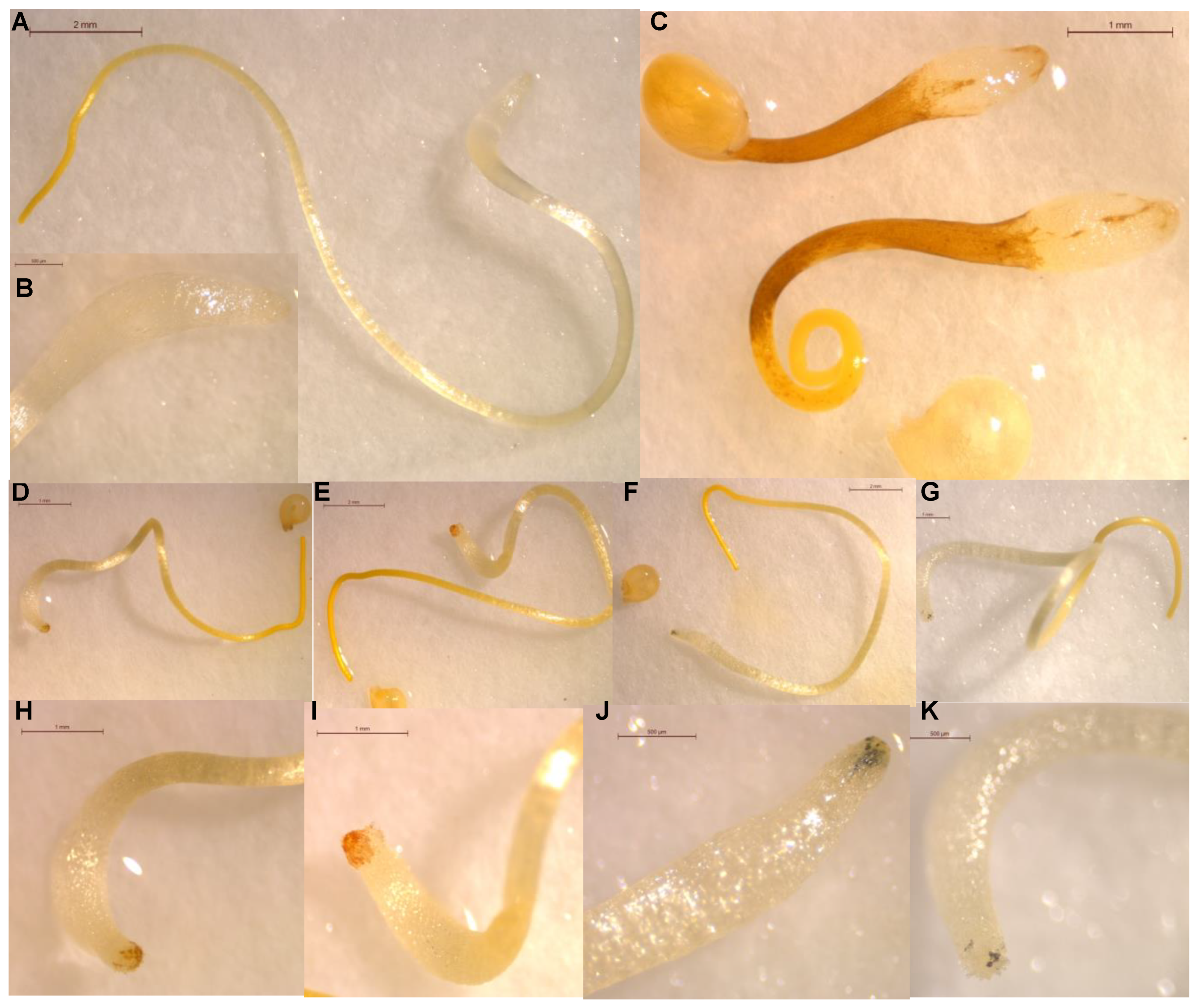{kind=link}
{kind=link}
| CLogP | CLogP | CLogP | CLogP | ||||
| 1 | −3.424 | 8 | 0.975 | 15 | 1.564 | 22 | 1.236 |
| 2 | 1.966 | 9 | 1.421 | 16 | 0.208 | 23 | 1.186 |
| 3 | 2.376 | 10 | 1.352 | 17 | 2.445 | 24 | 0.491 |
| 4 | −1.556 | 11 | 1.623 | 18 | 2.420 | 25 | 2.845 |
| 5 | 1.158 | 12 | 1.355 | 19 | 2.046 | 26 | 0.710 |
| 6 | 1.903 | 13 | 1.885 | 20 | 2.616 | 27 | 2.021 |
| 7 | 1.572 | 14 | 0.158 | 21 | 2.766 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Robles, A.; Cala Peralta, A.; Soriano, G.; Zorrilla, J.G.; Masi, M.; Vilariño-Rodríguez, S.; Cimmino, A.; Fernández-Aparicio, M. Identification of Allelochemicals with Differential Modes of Phytotoxicity against Cuscuta campestris. Agriculture 2022, 12, 1746. https://doi.org/10.3390/agriculture12101746
Moreno-Robles A, Cala Peralta A, Soriano G, Zorrilla JG, Masi M, Vilariño-Rodríguez S, Cimmino A, Fernández-Aparicio M. Identification of Allelochemicals with Differential Modes of Phytotoxicity against Cuscuta campestris. Agriculture. 2022; 12(10):1746. https://doi.org/10.3390/agriculture12101746
Chicago/Turabian StyleMoreno-Robles, Antonio, Antonio Cala Peralta, Gabriele Soriano, Jesús G. Zorrilla, Marco Masi, Susana Vilariño-Rodríguez, Alessio Cimmino, and Mónica Fernández-Aparicio. 2022. "Identification of Allelochemicals with Differential Modes of Phytotoxicity against Cuscuta campestris" Agriculture 12, no. 10: 1746. https://doi.org/10.3390/agriculture12101746
APA StyleMoreno-Robles, A., Cala Peralta, A., Soriano, G., Zorrilla, J. G., Masi, M., Vilariño-Rodríguez, S., Cimmino, A., & Fernández-Aparicio, M. (2022). Identification of Allelochemicals with Differential Modes of Phytotoxicity against Cuscuta campestris. Agriculture, 12(10), 1746. https://doi.org/10.3390/agriculture12101746