Abstract
Sweet chestnut fruits are popular fruits commercialized as fresh or processed ready-to-eat products. The major post-harvest problems associated with stored chestnut fruits are fungal rots, which cause major losses in fruit quality. The aims of this work were to determine the incidence, abundance and diversity of rots and fungi in three chestnut varieties (Longal, Judia and Martaínha) of Portugal, collected from an industrial plant, and to identify the stages of storage and processing where fungi and rots are more significant. Thirty-three chestnut samples from the three varieties were collected from different stages of industrial processing. Nuts were internally and externally inspected for damage, infestation and infection, and internal fungi were isolated and molecularly identified. The variety Martaínha was identified as the least susceptible to fungal growth, while Longal was the most susceptible. A high diversity of fungi was detected and identified. The dominant fungi were Mucor racemosus, Penicillium spp. (the causal agents of green rots), Ciboria batschiana (black rot) and Botrytis cinerea (gray rot). Gnomoniopsis smithogilvyi, the causal agent of brown rot, was also frequently detected. Sterilization with hydrothermal bath was effective in the elimination or reduction of most of the rot-causing fungi. These results could serve as a baseline for better monitoring fungal development and chestnut decay, and to develop effective management measures to control post-harvest chestnut rots.
1. Introduction
Chestnuts (Castanea sativa Mill.) play an important role in human nutrition owing to their nutrient composition and their potentially beneficial effects on health. The chestnut fruit is characterized by high content of water, starch and unsaturated fatty acids, and low fat content. Chestnuts are considered a good source of energy, especially because they are cholesterol-free. The fruits are also rich in fiber, vitamins and polyphenols, with gallic and ellagic acids predominating among the hydrolyzable and condensed tannins [1]. They are recommended as part of a gluten-free diet in case of celiac disease, and they help in the reduction of coronary heart disease and cancer [1,2].
Portugal is the fourth largest producer of sweet chestnut in Europe and seventh in the world, with an annual production of 24.7 thousand tons and an orchard area of 51 thousand hectares [3]. Trás-os-Montes, in the northeast of the country, is the first Portuguese chestnut producer region, with 80% of the national production. The chestnuts produced in this region are of high quality, and are recognized by the European Union with the Protected Denomination of Origin “Castanha da Terra Fria”.
Chestnut fruit is a seasonal product, commercialized as fresh or processed ready-to-use products. The high moisture and the rich nutrient content of the chestnuts represent storage problems since these conditions are supportive of infestation by insects and infection by a wide spectrum of spoilage fungi [4,5], resulting in major losses in fruit quality, potentially leading to completely unmarketable fruits. These conditions make it necessary for chestnuts to be controlled at both pre- and post-harvest stages.
The major post-harvest problems associated with these fruits are rots, which can be caused by several fungal agents. Traditionally, Penicillium sp., Aspergillus sp., Fusarium sp., Phomopsis castanea, Acrospeira mirabilis, Ciboria batschiana (syn. Sclerotinia pseudotuberosa), Phoma castanea and Phomopsis endogena have been reported in many different countries worldwide as the most important fungi responsible for chestnut rots and fruit post-harvest decay [5,6,7,8,9,10,11,12]. In the past decade, however, a steep rise in the incidence of rotten nuts has been extensively observed by chestnut growers in some regions of Europe and Australasia [13,14,15,16], and the fungus Gnomoniopsis smithogilvyi L.A. Shuttlew., E.C.Y. Liew and D.I. Guest (2012) (syn. Gnomoniopsis castaneae Tamietti 2012) has been identified as a significant rot-causing agent [7,16,17,18,19,20,21].
Chestnut rots are mainly expressed post-harvest, and nuts show steep decay during storage. Even though many fungi can be detected superficially on the shell [5,6,12], most of the fungi responsible for internal nut rot are considered endophytic to the plant. After harvest and during storage, the fungi develop internally and affect the kernel of the nuts, resulting in internally rotten nuts that are superficially apparently healthy [7,12]. For instance, while Penicillium sp., Sclerotinia sp. and Trichoderma sp. have been shown to start developing nut rot symptoms even before harvest, others such as Coniophora sp., Botryosphaeria sp., Botryotinia fuckeliana (anamorph Botrytis cinerea) and Giberella sp. (anamorph Fusarium sp.) seem to develop only during storage [12].
In this work, we intended to determine the incidence, abundance and diversity of rots and fungi in three chestnut varieties (Longal, Judia and Martaínha) of Trás-os-Montes, Portugal, collected from an industrial plant, and to identify the stages of storage and processing where fungi and rots become more significant.
2. Materials and Methods
2.1. Collection of Samples
The study was performed on European chestnuts (C. sativa) received, processed and stored in a local industry in Braganca, Trás-os-Montes, Portugal, during the growing season in 2018–2019. Samples were collected in different processing stages, as detailed in Table 1, from three varieties (Longal, Judia and Martaínha), for a total of 33 samples. The sampling plan was adjusted to the availability of the industry, and this resulted in a different number of samples for the two factors—stage of processing and variety—under study.

Table 1.
Description of chestnuts sampling points, storage period and sampled varieties.
The samples (of approximately 1 kg of chestnuts) were collected from 1-ton wood palox pallets from the storage room (stages of storage/process P2, P3, P6, P7, P8 and P9) or from the rejection palox (P10), and packaged separately in paper envelopes. The envelopes were immediately sealed and transported to the laboratory. Chestnut samples were stored in a cold chamber at 4 °C for a maximum of 2 days.
2.2. General Inspection of Chestnut Samples
Fifty chestnuts per sample (for a total of 1650 chestnuts) were randomly selected for external inspection. The external quality of fruits was evaluated by checking for visual defects, i.e., visible signs of pre- and post-harvest damage, including cracks, signs of infestation (presence of exit holes caused by the chestnut weevil) and signs of fungal infection (visible mold growth), as described by Overy et al. [22]. The observed symptoms were described for each chestnut to determine the percentage of external infection and infestation per sample.
After external inspection, the 50 fruits were cleaned under running tap water, surface-disinfected by immersing in 10% commercial bleach for 2 min, washed three times with sterile distilled water and blot-dried in sterile absorbent paper. Chestnuts were aseptically bisected longitudinally from the stylar end to hilum with a sterile knife and visually inspected for the presence of internal molds and insect larvae. Whenever signs of rot were present, the type of rot was identified. The percentage of rotten surface of each chestnut was visually estimated to determine the level of infection: Level 0 (L0): no visible mold infection; Level 1 (L1): 1–25% of chestnut internal surface covered by molds, Level 2 (L2): 26–50%, Level 3 (L3): 51–75%, Level 4 (L4): 76–100%, as proposed by Donis-González et al. [12]. The disease severity index, or McKinney index [23], was also determined. This index takes into account the intensity of the rot (severity), its frequency and the maximum possible value, according to the equation:
where d is the category of rot intensity scored for chestnuts, f is the rot frequency, N is the total number of examined chestnuts (healthy and rotted) and D is the highest category of rot intensity that occurred.
2.3. Isolation of Fungi from Chestnuts
From each sample, 10 chestnuts with symptoms of kernel infection were sorted for the isolation and identification of contaminating fungi. Five explants of the rotten parts were cut aseptically and plated on solidified potato dextrose agar (PDA; Biolife, Milan, Italy) in 9 cm Petri dishes. For samples with less than 10 nuts showing visible signs of fungal infection, non-infected chestnuts were used to complete the 10 chestnuts per sample. Plates with the explants were sealed with parafilm and incubated at 25 °C for 6 days.
After the incubation period, each different growing colony was isolated and transferred the number of times necessary to obtain pure cultures into 6 cm Petri dishes containing 10 mL of Malt Extract Agar (MEA: malt extract 20 g/L, glucose 20 g/L, peptone 1 g/L, agar 20 g/L, autoclaved for 15 min at 121 °C) for morphological and molecular identification. Plates were incubated using the same conditions as before. A total of 544 fungi were isolated from the samples. Fungal isolates were morphologically identified on the basis of their cultural characteristics such as colony size, shape, opacity, color, form, surface growth, elevation and margin consistency, and grouped by morphotypes. From these, 78 isolates representative of all morphotypes were selected for molecular identification. All pure isolates (spores and mycelium) were preserved in 30% glycerol with 0.05% of Tween 80 at −20 °C until further analysis [5].
2.4. Molecular Identification of Fungal Isolates
The total genomic DNA was extracted according to the SDS protocol described by Rodrigues et al. [24]. The DNA was dissolved in 30 to 50 μL of ultrapure water depending on the yield and stored at 20 °C. The universal primers ITS1-F (5′ CTTGGTCATTTAGAGGAAGTAA-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [25,26], which amplify a 600 bp segment of the ITS1/5.8S/ITS2 region of the rRNA gene, were used for sequencing purposes. PCR amplifications were performed as described by Rodrigues et al. [24]. PCR products were purified with the commercial GF-1 PCR cleanup kit (Vivantis Technologies, Malaysia), according to the instructions of the manufacturer. Sequence data were obtained by Sanger sequencing. The sequences were manually corrected by comparison with the chromatogram and aligned with the NCBI (National Centre for Biotechnology Information) GenBank database (http://www.ncbi.nlm.nih.gov/ (accessed on 10 November 2019) using the BLAST algorithm to identify the fungi.
2.5. Fungi Guild Classification
Each taxon (genus) was assigned to an ecological guild, based on FUNGuild [27]. The genera were distributed into their respective guild class: animal pathogen, endophyte, hyperparasitic, plant pathogen, plant saprotroph, mycoparasitic, saprobiotic, soil saprotroph and wood saprotroph. Some fungi do not exclusively belong to one guild, and were put in a class with the combined guilds.
2.6. Statistical Analyses
For the comparison of means of quantitative variables, samples were tested for homogeneity of variances by Levene’s test. Since samples failed this criterion, variances were analyzed by one-way ANOVA, and multiple comparisons between samples pairs were computed using Tamhane’s T2 test. The mean differences were significant at p < 0.05.
Fungal incidence was determined as the percentage of samples contaminated with a given species. Fungal frequency was determined as the percentage of isolates of a given species compared with the total number of isolates. The Shannon–Wiener diversity index (H’) and the Pielou evenness index (J’) were calculated by PRIMER 5.2.2 [28]:
where pi is the proportion of each species in the community. The logarithmic e base was used.
where H’obs. is the observed diversity and H’max (=lnS) is the maximum possible diversity.
H’ = −∑[(pi) × ln(pi)]
J’ = H′obs./H′max
For statistical analyses, quantitative variables were tested for normality (Shapiro–Wilks and Kolmogorov–Smirnov tests). The variables with a normal distribution were analyzed by one-way ANOVA, followed by Tukey’s Honest Significant Difference (Tukey-HSD) test. The variables that did not achieve normal distribution despite transformation were analyzed by the non-parametric Kruskal–Wills ANOVA and Median test for multiple independent groups, followed by the multiple comparisons Mann–Whitney U test between independent groups.
To determine which fungi species contribute the most to the discrimination among the two a priori categorical groups (chestnut variety or processing stage), the raw data were analyzed by the multiple discriminant analysis (MDA)—first with all species and later without punctual species (after removal of 9 species that were isolated only once). For this purpose, a forward stepwise analysis with maximum 0.010 tolerance was computed. Parametric and non-parametric tests and MDA were analyzed with STATISTICA version 10 [29]. The null hypotheses were rejected when p > 0.05.
3. Results and Discussion
3.1. Chestnut Overall Inspection
The results of damage observed in chestnut samples after external inspection are presented in Figure 1. Damaged chestnuts were present in all samples from the three varieties with different percentages. The highest percentages of damaged chestnuts were detected in the variety Longal for all the analyzed processing stages. When varieties were considered in bulk (Longal, Judia and Martaínha, independently of the processing stage), significant differences were observed between Longal and Judia (p < 0.05).
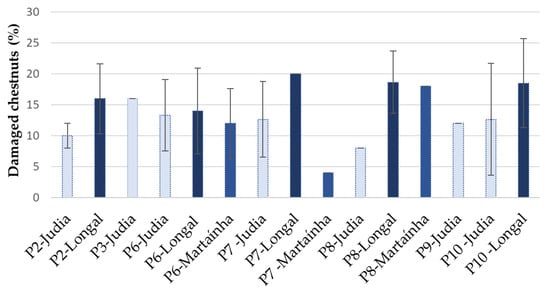
Figure 1.
Percentage of external damaged chestnuts at different processing stages for the chestnut varieties Longal (dark blue), Judia light blue) and Martaínha (intermediate blue) (mean ± SD; n = 50). P2, P3—sterilized chestnuts (0 and 15 days storage); P6—unsterilized chestnuts in reception; P7, P8, P9—unsterilized chestnuts after storage (15, 30 and 45 days, respectively); P10—sterilized chestnuts rejected by sorting.
The lowest percentage of damage was observed in Martaínha, stage P7. When comparing external damage among stages of processing, regardless of the variety, no significant differences were observed (p > 0.998). This result could be due to the high differences among chestnut varieties for the same processing stage.
The observed external and internal symptoms of infestation were described and counted to determine the percentage of infestation, as summarized in Figure 2. Attention is drawn to the fact that these results were only analyzed as trends; they were not statistically computed, given the low and different number of replicas among processing stages.
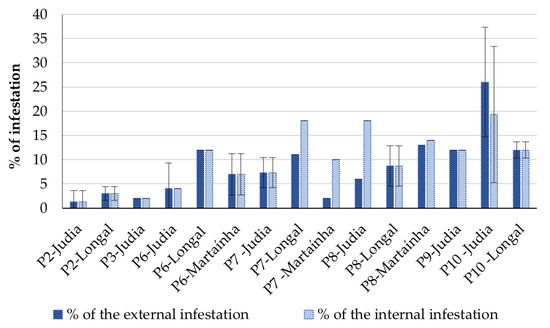
Figure 2.
Percentage of external and internal infestation at different processing stages for the varieties Longal, Judia and Martaínha (mean ± SD; n = 50). P2, P3—sterilized chestnuts (0 and 15 days storage); P6—unsterilized chestnuts in reception; P7, P8, P9—unsterilized chestnuts after storage (15, 30 and 45 days, respectively); P10—sterilized chestnuts rejected by sorting.
The highest percentage of internal infestation was registered in samples from Judia, processing stage P10. This result was expected, because the processing stage P10 corresponds to the chestnuts rejected based on visual defects. The sterilized chestnuts from processing stages P2 and P3 (0 and 15 days of storage after sterilization) presented the lowest percentage of chestnut infestation. On the other hand, the chestnuts stored immediately after reception without going through the sterilization process (P6 to P9) showed a higher percentage of internal infestation. Storage time showed to have a negative impact on infestation trend in non-sterilized chestnuts, since internal infestation increased concomitantly with the augmentation of the period of storage.
The higher average of infestation was detected for the variety Longal compared to Judia, in all processing stages except for P10.
3.2. Level of Chestnut Rot
The level of internal fungal infection of chestnuts was estimated by determining the percentage of rotten chestnuts showing different levels of visible infection. The results are presented in Figure 3 (levels of infection) and Figure 4 (McKinney index). Samples stored without sterilization generally presented higher levels of infection, reflecting in higher fruit decay. Comparing samples among varieties, the variety Judia showed the lowest percentage of infection during the processing stages P2, P6, P7 and P10, while Longal presented a higher percentage of infection for Level 2 or more, with a concomitant higher McKinney index. Of the three varieties tested, Longal was the most sensitive to fungal infection and rot decay, when similar stages of processing are considered. This variety has been associated with the highest contents of protein, fat and starch [30,31,32], which potentially stimulate fungal establishment and development, while showing the lowest contents of total phenolics, ellagic and gallic acids [32]. Phenolics in general, and particularly ellagic and gallic acids, have shown to play a key role in plant defense against fungal pathogens by enhancing plant antioxidative responses or ergosterol inhibition, among other modes of action [33,34].
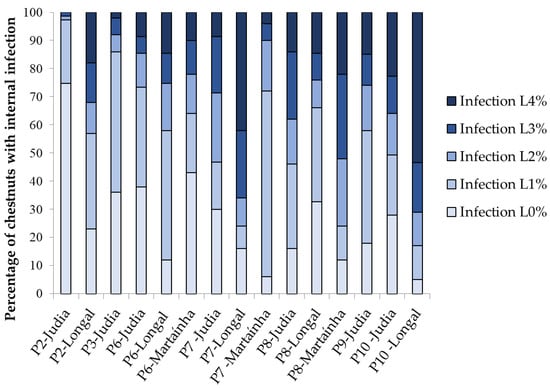
Figure 3.
Percentage of chestnuts showing internal fungal infection at different processing stages for the varieties Longal, Judia and Martaínha. Levels of infection: L0—no visible mold infection; L1—1–25% of chestnut internal surface covered by molds; L2—26–50%; L3—51–75%; L4: 76–100%.
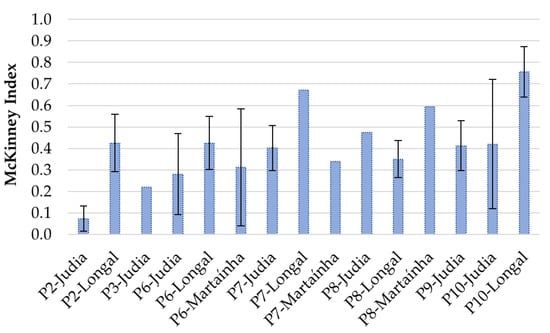
Figure 4.
McKinney index for the infected (rot) chestnuts recorded at different processing stages for the varieties Longal, Judia and Martaínha.
Comparing non-sterilized Judia samples with 0, 15 and 30 days of storage (P6, P7 and P8), there is a clear increase in the level of infection throughout time, showing that the storage conditions allow for the proliferation of fungi and rot. For Longal, the trend is not evident, since a decrease in visual rot (higher % L0 and L1, lower McKinney index) is observed from P7 to P8.
The observed level of decay (as determined by the McKinney index) correlated positively and significantly with all the previously mentioned visual parameters, namely the level of external damage (r = 0.579, p = 0.000), external and internal signs of infestation (r = 0.647, p = 0.000 and r = 0.593, p = 0.000, respectively) and with the external level of fungal infection (r = 0.585, p = 0.000). These correlations were also reported by others [22,35]. Internal fungal infection and rot decay are not fully dependent on shell damage and oviposition wounds, since some fungi responsible for internal infection are considered endophytes. However, such shell breaches provide penetration sites to external fungi, and the oviposition inside the kernel can have a role as a vector of fungal spores [22,36].
3.3. Fungal Species Frequency and Incidence
A total of 557 fungal isolates were obtained from chestnut samples. Of these, 78 isolates were sequenced for molecular identification by the ITS region. The fungal species identified in this study, with reference to the most similar strain used for identification, are given in Table S1, along with the GenBank accession numbers of the comparison strains and the percentage of similarity. The number of isolates of each species molecularly identified is also shown. From the sequenced isolates, 37 different species were identified, scattered among 18 families and 23 genera: Alternaria, Aureobasidium, Botryotinia, Botrytis, Ciboria, Cladosporium, Coleophoma, Coniella, Curvibasidium, Cytospora, Didymella, Fusarium, Gnomoniopsis, Lanzia, Mollisia, Mucor, Penicillium, Peniophora, Phacidium, Pilidium, Sporothrix, Stromatina and Trichoderma.
Along the storage and the processing stages, the frequency of the species varied greatly (Figure 5a). Globally, in all samples, the results showed a clear predominance of ascomycetous molds (91.9%): of the 37 species identified, only three belonged to other phyla, namely Mucormycota (Mucor racemosus) and Basidiomycota (Curvibasidium cygneicollum and Peniophora meridionalis). These proportions are comparable with those previously reported [36,37].
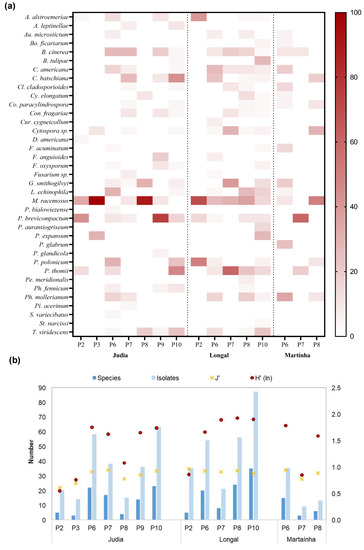
Figure 5.
(a) Heat map of the frequency of the fungal species (in percentage of analyzed chestnuts), obtained for each storage and processing stages of the Judia, Longal and Martaínha varieties; (b) their respective total number of isolates and species, and diversity indexes (Shannon–Wiener, H’, and Pielou, J’).
Mucor racemous, a cosmopolitan fungus widely distributed in nature, was the most prevalent species (between 24.2% and 100% frequency), and it was present in all stages of production, even after chestnut sterilization (P2, P3 and P10). The resistance of the Mucoraceae to warm bath was also reported by Morales-Rodriguez et al. [38].
The second highest frequency was detected for the genus Penicillium, able to proliferate throughout storage in both sterilized and non-sterilized chestnuts. These fungi have a strong conidial proliferation, and are the causal agents of the highly aggressive and destructive green and blue rots (Figure 6a), as also described by Donis-Gonzalez et al. [12]. Of the eight Penicillium species identified in this study, P. brevicompactum, P. thomii and P. polonicum were the dominant ones. Others have previously reported similar results, with Penicillium species showing up to 100% incidence [11,12,22,35]. Prencipe et al. [39] also identified P. bialowiezense, P. brevicompactum, P. glandicola and P. polonicum as chestnut contaminants. More recently, Morales-Rodriguez et al. [36] reported P. brevicompactum as one of the most abundant contaminants of chestnuts. This Penicillium species is not only a saprotroph capable of causing green rot in fruits, but is also associated with the production of several mycotoxins in chestnuts and in other fruit species [39,40,41,42,43].
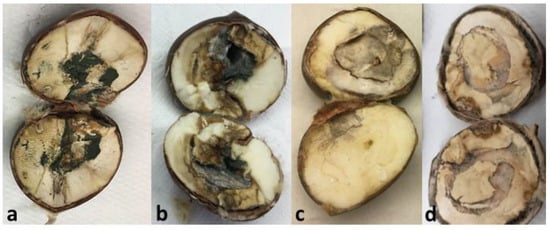
Figure 6.
Chestnuts with visible and advanced rot caused by Penicillium brevicompactum (a), Ciboria batschiana (b), Botrytis cinerea (c) and Gnomoniopsis smithogilvyi (d). The rot-causing agents were isolated and molecularly identified.
B. cinerea, generally associated with fruit decay and considered the causal agent of gray rot in chestnuts [11,12] (Figure 6b), was also frequently isolated. C. batschiana, showing the same frequency of detection as B. cinerea, has been identified as the causal agent of chestnut black rot (Figure 6c) and is responsible for causing severe post-harvest losses by disease [12,17]. It has been reported in Greece [44], Italy [9,17] and the USA [12]. C. batschiana and B. cinerea, with an average frequency of 10.6%, were detected in all the stages except in the sterilized samples. Nevertheless, C. batschiana achieved the highest frequency in rejected chestnuts (21.4%, in P10, Judia), which can mean that a significant level of black rotten chestnuts is eliminated by sorting.
G. smithogilvyi is currently considered the major causal agent of chestnut brown rot in several countries in Europe, and also in New Zealand and Australia. It has been found in different climatic regions in Switzerland [20], and has been reported to be a ubiquitous pathogen in Australia [14,45], in Italy [16,17], in Switzerland [20] and in the United Kingdom [46]. G. smithogilvyi causes characteristic texture degradation and brown rot in the endosperm [17], showing conidiomata when in advanced infection levels (Figure 6d). G. smithogilvyi, which is reported in this study for the first time in post-harvest chestnuts from Portugal, was more frequently detected in non-sterilized samples with 15 or more days of storage, showing up to 40% incidence, and, interestingly, it was absent from sterilized samples (Figure 5a). In laboratory trials, the hydrothermal bath was also identified as the critical phase for the inactivation of G. smithogilvyi in chestnut fruits [38].
Along the processing stages, the highest number of isolates were, as expected, recovered from the rejected chestnuts (P10), for Judia (63) and Longal (87) varieties. The maximum percentage of fungal frequency was detected during this stage for P. thomii (28.6%), T. viridescens (22.9%), the causal agent of black rot C. batschiana (21.4%), and B. tulipae (17.1%). The causal agent of brown rot, G. smithogilvyi, although not highly frequent (10.0%), showed a relatively important position in sample contamination. By contrast, P2 showed the lowest level of fungal contamination, with Mucor, Penicillium and Alternaria contributing with the highest percentages. The sterilization process by hydrothermal bath seems to be ineffective for the elimination of these three fungi.
When P2 (sterilized) and P6 (non-sterilized) samples are compared, it is evident that the sterilization process has an important effect on fungal contamination. Overall, the results show a high percentage of fungal incidence in non-sterilized samples (P6 to P9), although time of storage could not be clearly associated with increased level of contamination. These inconsistencies may be related to the number of repetitions per sample, which, due to industrial limitations in providing all kinds of samples, was not constant for all processing stages and chestnut varieties. As previously reported, the higher number of samples may justify the higher species richness [47].
Non-sterilized chestnut samples (P6, P7, P8 and P9) were compared in order to determine the effect of the storage period. Samples with the longest storage period (45 days, P9) showed higher levels of fungal infection (Figure 3) and rot incidence than samples with shorter storage periods (P6, P7 and P8). As also reported by Washington et al. [34], the storage period was found to influence the proliferation of fungi. In general, the decay incidence increased with longer storage period and seems to also depend on cultivar [35]. At harvest, the chestnuts already infected by rot-causing fungi will colonize the fruit during storage [34]. Storage time also contributed to the elimination of the fungi that are less resistant to the storage conditions. By comparing the richness (number of different fungal species, Figure 5b), the diversity of fungi decreased with the storage period, from 27 species in P6 to 11 species in P9. Moreover, a decrease in both the abundance and diversity of fungal communities on chestnuts during storage period was recently reported [37].
Shannon–Wiener diversity (H’) revealed a similar pattern of total species (Figure 5b), with the highest values being reached in P6 to P10, and the lowest values in P2 and P3 samples. The evenness index (J’) ranged between 0.609 and 0.964 in Judia (P2) and Longal (P2), respectively, being virtually constant in the Longal and Martaínha samples. Contrary to this, in Judia samples, J’ underwent greater variations.
Rapid, accurate, real-time and non-destructive methods capable of monitoring, detecting and quantifying rot incidence and severity, and even of differentiating the type of rot or causal agent during storage and processing, would significantly benefit the chestnut supply chain manufacturing process, even more so if we consider that the onset of decay starts long before the onset of visible signs [48,49]. Devices such as molecular- or biochemical-based sensors have been developed for the timely detection of various post-harvest pathogenic fungi or specific decay-related derivative metabolites, such as volatiles [50,51,52,53], and some are even able to differentiate different rotting fungi [50,52].
Many of these methods have been developed and tested with success at controlled laboratorial or small-scale levels [49], and now need to be scaled up to the agri-food industries. Valuable information on decay development along the supply chain would facilitate rational management decisions.
3.4. Fungal Species Richness and Diversity among Chestnut Varieties
The distribution among chestnut varieties of the frequency of the 37 fungal species is displayed in Figure 7a. The results show that M. racemosus f. sphaerosporus had the highest incidence in all varieties with identical values. P. brevicompactum attacked the varieties Judia and Martaínha with almost the same incidence (21.7% and 20%, respectively) and was less present in Longal (7.7%). T. viridescens, P. expansum and Con. fragaria showed similar incidences in the varieties Judia and Longal.
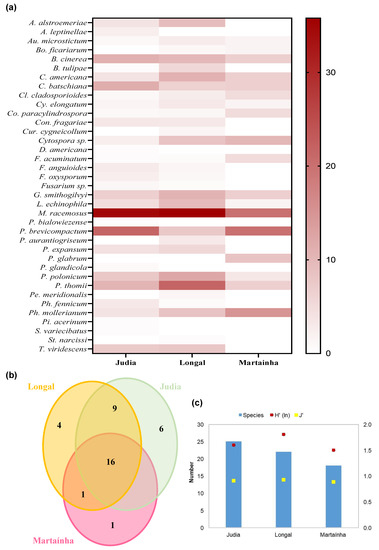
Figure 7.
(a) Heat map of the frequency of the fungal species (in percentage of analyzed chestnuts), obtained for each variety (Judia, Longal and Martaínha); (b) Venn diagram showing the unique and shared number of species in the 3 varieties; (c) total number of species and Shannon–Wiener, H’, and Pielou, J’, indexes for the common processing stages (P6, P7 and P8).
For the Judia variety, the highest incidence of fungi affecting chestnuts was for M. racemosus and P. brevicompactum, with nearly 22% each, followed by C. batschiana, B. cinerea and P. thomii. The Longal variety presented higher frequencies of M. racemosus, P. thomii, P. polonicum, C. americana, and B. cinerea. In Martaínha, besides M. racemosus and P. brevicompactum, the dominant species included Ph. mollerianum, P. glabrum and Cytospora sp.
Some fungi seem to have preference for certain varieties. C. americana showed higher incidence in Longal, while Judia was more contaminated by C. batschiana. If Penicillium species are considered, P. thomii and P. polonicum were predominant in Longal, while P. brevicompactum was more common in Judia and Martaínha.
Sixteen (43.2%) of the thirty-seven total species are common to the three chestnut varieties (Figure 7b). Among these shared species are fungi responsible for several rots: black rot (C. batschiana), brown rot (G. smithogilvy), gray rot (B. cinerea and M. racemus) and green rot (Penicillium spp.). Species such as Au. microstictum and C. americana, associated with mummified chestnuts, and Cl. cladosporioides, Cy. elongatum, Co. paracylindrospora, Cytospora sp., F. acuminatum, L. echinophila and Ph. mollerianum were also isolated from the three varieties. Judia and Longal varieties shared nine species (24.3%): A. alstroemeriae, B. tulipae, Con. fragariae, F. anguioides, F. oxysporum, Fusarium sp., P. expansum, Ph. fennicum and Trichoderma viridescens. One species (Bo. ficariarum) was shared by Longal and Martaínha, while none were shared by Judia and Martaínha. The remaining species (11–29.7%) were unique of each variety: (i) P. glabrum in Martaínha; (ii) A. leptinellae, Didymella americana, P. bialowiezense, P. glandicola, Pilidium acerinum and Sporothrix variecibatus in Judia; and (iii) Cur. cygneicollum, Penicillium aurantiogriseum, Peniophora meridionalis and Stromatinia narcissi in Longal. All the unique species from Longal and five species from Judia were isolated only once. Curiously, all the Basidiomycota species (Cur. cygneicollum and Peniophora meridionalis) came from Longal chestnuts. In a recent work [54], Cur. cygneicollum was found in galled twigs of Castanea molissima.
Many of the species isolated in this work were reported by others in association with C. sativa tree nuts, crust, leaves and pollen [55], and in C. sativa galls [56]. Contrary to Kačániová et al. [55], who isolated several Aspergillus spp. from nuts in Slovakian chestnut orchards, no Aspergillus spp. were recovered from chestnuts in this study.
Techniques traditionally applied in plant pathology diagnostics show many limitations, namely those associated with fungal cultivation methods, thus hampering the identification of the true fungal diversity [57]. The development of further research using high-throughput DNA sequencing (HTS) and metagenomics technologies will allow an in-depth characterization of the fungal community composition, including the detection of the uncultivable ones.
For the common processing stages (P6 to P8), Longal had the highest H’ and J’ values (Figure 7c), followed by Judia and Martaínha. The evenness index (J’) was lower in Martaínha (0.885), indicating the presence of more dominant fungal species than in Judia ((J’ = 0.906) and in Longal (J’ = 0.927).
3.5. Mold Guilds According to Stage Processing and Chestnut Variety
Nine functional groups were associated with the species identified (Figure 8; Table S2) [58,59,60,61,62,63]. Overall, Saprobiotic, Saprobiotic–Plant pathogen, and Plant pathogen were the most representative guilds in all processing stages of the three varieties (Figure 8a). Except for P2, the other processing stages ranged from three to six guild types. P2 had the lowest guild diversity, dominating the Saprobiotic, followed by Plant pathogen (Judia and Longal) and Endophyte–Plant pathogen–Wood saprotroph (only in Judia) guilds. Judia was the only variety showing the guild Mycoparasitic (P6), and the complex groups Animal pathogen–Endophyte–Plant pathogen–Wood saprotroph (P7 and P10) and Animal pathogen–Endophyte–Plant pathogen–Soil saprotroph (P6). Both P6 and P7 were not submitted to sterilization, and P10 corresponds to sterilized but rejected chestnuts.
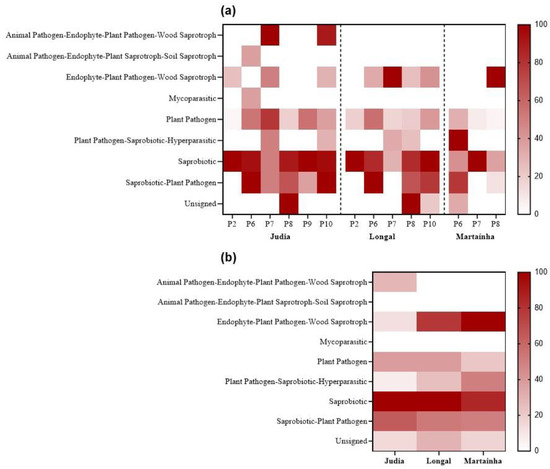
Figure 8.
Heat maps of the frequency of the fungal guilds obtained (a) for each processing stage for the chestnut varieties (Judia, Longal and Martaínha); (b) per variety.
Figure 8b displays the mean values of guild frequencies among chestnut varieties. The two guilds Saprobiotics and Saprobiotic–Plant pathogen dominated in the three varieties. Contrary, the Plant pathogen–Saprobiotic–Hyperparasitic guild exhibited a decreasing representativeness from Martaínha > Longal > Judia, a pattern also followed by the Endophyte–Plant pathogen–Wood saprotroph guild.
Not many studies report the fungal guilds associated with chestnuts. In green leaves and necrotic galls sampled in chestnut orchards, in Italy, 10 functional groups were found and the dominating ones were Endophytes and Plant pathogens [56].
3.6. Mold Community Analysis According to Processing Stage and Chestnut Variety
For MDA analysis, the species that appeared only once were removed. Of a total of selected 30 species, 26 were included in the model, while 4 were not (A. alstroemeria, F. oxysporum, Ph. mollerianum and L. echinophila). Using the processing stage as the discriminant factor (Figure 9a), the MDA of the mold species frequency showed distinct communities among the stages (Wilks λ = 0.00000, F = 6.7902, p < 0.0001). The first two root functions, explaining 95.8% of the total discriminant power, showed that all samples were clearly separated (Figure 9a). The squared Mahalanobis distances were significant (p < 0.05) between P6 (without sterilization, day 0) and all the stages (with and without sterilization, days 15, 30 and 45), between P8 and P9, and P9 and P10. The mycobiota composition varied between sterilized/unsterilized samples and storage periods. These differences are probably a consequence of physicochemical changes in chestnuts. Throughout chestnut storage, parameters such as starch, total soluble sugars, reducing sugars, soluble protein and moisture contents were found to decrease [37]. These authors found a strong correlation between fungal community and chestnut quality: fungal decay incidence, starch and malondialdehyde content changes along storage. In addition, hydrothermal bath provokes changes in the chemical composition of the fruits, decreasing the starch concentration and increasing the concentration of sucrose [10].
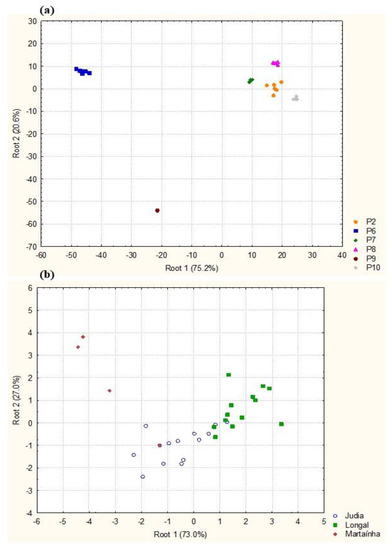
Figure 9.
Diagrams for the first two roots obtained by MDA of mold species composition in processing stages (a) and chestnut varieties (b). P2—sterilized chestnuts; P6—unsterilized chestnuts in reception; P7, P8, P9—unsterilized chestnuts after storage (15, 30 and 45 days, respectively); P10—sterilized chestnuts rejected by sorting.
MDA analysis suggests that mold communities are different among the three chestnut varieties (Figure 9b, Wilks λ = 0.11219, F = 2.7492 p < 0.01). Of the 30 species, 13 were responsible for significant differences, particularly the species P. glabrum, Cytospora sp., C. americana, Cy. elongatum, A. leptinella, Cl. cladosporioides, B. tulipae, Bo. ficariarum and Ph. fennicium. The squared Mahalanobis distances were highly significant between Longal and Martaínha (p = 0.0041) and between Longal and Judia (p = 0.01905). This analysis suggests differences in the fungal communities associated with chestnut varieties.
As previously mentioned, the three varieties tested show divergent chemical and nutritional compositions [30,31,32], which potentially define the establishment and development of different mycobiota based on their own nutritional requirements. For instance, Judia and Longal have higher protein and fat content than Martaínha [31,32], which might explain the higher incidence of proteolytic and lipolytic fungi such as Penicillium spp. in those varieties. On the other hand, Longal has been associated with higher starch contents [31], which has been reported as an excellent substrate for G. smithogilvyi [64]. Differences in fungal incidence and severity of colonization were also reported by Sieber et al. [11].
4. Conclusions
In the industry, the storage of chestnuts is a major problem due to the growth of a wide spectrum of spoilage fungi, which lead to the appearance of rots. As a result, chestnuts lose their fruit quality and commercial value.
Among the dominant species identified in this study, some were previously associated with chestnut rot or decay: B. cinerea, the causal agent of gray rot; G. smithogilvyi, the main causal agent of chestnut brown rot in the world; C. batschiana, identified as the agent of chestnut black rot and responsible for serious post-harvest losses; Penicillium spp., responsible for green rot, particularly P. brevicompactum, which is also associated with the production of several mycotoxins in chestnuts.
The results also suggest that the mycobiota diversity differed with the chestnut cultivars, processing stages and storage periods. The variety Martaínha was identified as the least prone to fungal growth, while Longal was the most susceptible to infection. Sterilization with hydrothermal bath appeared to be effective in eliminating or reducing most rot-causing fungi, such as G. smithogilvyi, C. batschiana and B. cinerea, although other fungi (Mucor, Penicillium and Alternaria) seem to be more resistant to this sterilization method.
The nut–fungal interactions in different varieties and processing stages can serve as a baseline in monitoring fungal development and chestnut decay, in order to develop management measures and effective treatments to control the post-harvest fruit rot.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agriculture12111930/s1, Table S1: Fungal species identified in this study, with reference to the most similar strain used for identification, GenBank accession numbers of the comparison strains and percentage of similarity. The number of isolates of each species molecularly identified is also shown. Table S2: List of fungal species identified in this study, and their reported geographical distribution, guild, associated disease and hosts.
Author Contributions
Conceptualization, P.R., A.S. and J.G.-L.; methodology, P.R. and J.O.D.; validation, P.R. and A.S.; formal analysis, P.R. and A.S.; investigation, P.R. and J.O.D.; resources, P.R.; data curation, P.R.; writing—original draft preparation, P.R., J.O.D. and A.S.; writing—review and editing, P.R., and A.S.; supervision, P.R.; funding acquisition, J.G.-L. All authors have read and agreed to the published version of the manuscript. This research work is part of the Master’s thesis of Jihen Oueslati Driss.
Funding
This work was funded by the project “ValorCast—Valorização da castanha e otimização da sua comercialização”, ref. PDR2020-101-032030, funded by Fundo Europeu Agrícola de Desenvolvimento Rural (FEADER) and the Portuguese government, under the scope of Ação 1.1 “Grupos Operacionais”, Medida 1. “Inovação”, PDR 2020—Programa de Desenvolvimento Rural do Continente. The Foundation for Science and Technology (FCT, Portugal) and FEDER under Program PT2020 provided financial support to CIMO (UID/AGR/00690/2020), SusTEC (LA/P/0007/2020), CITAB (UID/AGR/04033/2020) and Inov4Agro (LA/P/0126/2020).
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors are grateful to Sortegel, Portugal, for providing the chestnut samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gonçalves, B.; Borges, O.; Costa, H.S.; Bennett, R.; Santos, M.; Silva, A.P. A metabolite of chestnut (Castanea sativa Mill.) upon cooking: Proximate analysis, fiber, organic acids, and phenolics. Food Chem. 2010, 122, 154–160. [Google Scholar] [CrossRef]
- Vasconcelos, M.C.B.M.; Bennett, R.N.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. The composition of European chestnut (Castanea sativa Mill.) and association with health effects: Fresh and processed products. J. Sci. Food Agric. 2010, 90, 1578–1589. [Google Scholar] [CrossRef] [PubMed]
- INE. Estatísticas Agrícolas 2020; Instituto Nacional de Estatística: Lisbon, Portugal, 2021. [Google Scholar]
- Rodrigues, P.; Venâncio, A.; Lima, N. Mycobiota and mycotoxins of almonds and chestnuts with special reference to aflatoxins. Food Res. Int. 2012, 48, 76–90. [Google Scholar] [CrossRef]
- Rodrigues, P.; Venâncio, A.; Lima, N. Incidence and diversity of the fungal genera Aspergillus and Penicillium in Portuguese almonds and chestnuts. Eur. J. Plant Pathol. 2013, 137, 197–209. [Google Scholar] [CrossRef]
- Wells, J.M.; Payne, J.A. Toxigenic Aspergillus and Penicillium Isolate from Weevil-Damaged Chestnuts. Appl. Microbiol. 1975, 30, 536–540. [Google Scholar] [CrossRef] [PubMed]
- Lione, G.; Danti, R.; Fernandez-Conradi, P.; Ferreira-Cardoso, J.V.; Lefort, F.; Marques, G.; Meyer, J.B.; Prospero, S.; Radócz, L.; Robin, C.; et al. The emerging pathogen of chestnut Gnomoniopsis castaneae: The challenge posed by a versatile fungus. Eur. J. Plant Pathol. 2019, 153, 671–685. [Google Scholar] [CrossRef]
- Washington, W.S.; Allen, A.D.; Dooley, L.B. Preliminary studies on Phomopsis castanea and other organisms associated with healthy and rotted chestnut fruit in storage. Australas. Plant Pathol. 1997, 26, 37–43. [Google Scholar] [CrossRef]
- Vettraino, A.M.; Paolacci, A.; Vannini, A. Endophytism of Sclerotinia pseudotuberosa: PCR assay for specific detection in chestnut tissues. Mycol. Res. 2005, 109, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Jermini, M.; Conedera, M.; Sieber, T.; Sassella, A.; Schärer, H.; Jelmini, G.; Höhn, E. Influence of fruit treatments on perishability during cold storage of sweet chestnuts. J. Sci. Food Agri. 2006, 86, 877–885. [Google Scholar] [CrossRef]
- Sieber, T.N.; Jermini, M.; Conedera, M. Effects of the harvest method on the infestation of chestnuts (Castanea sativa) by insects and molds. J. Phytopathol. 2007, 155, 497–504. [Google Scholar] [CrossRef]
- Donis-González, I.R.; Guyer, D.E.; Fulbright, D.W. Quantification and identification of microorganisms found on shell and kernel of fresh edible chestnuts in Michigan. J. Sci. Food Agric. 2016, 96, 4514–4522. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.C.; Agri, M. The life cycle, pathology, and taxonomy of two different nut rot fungi in chestnut. Austral. Nutgrow. 2008, 22, 11–15. [Google Scholar]
- Smith, H.C.; Ogilvy, D. Nut rot in chestnuts. Austral. Nutgrow. 2008, 2, 10–15. [Google Scholar]
- Gentile, S.; Valentino, D.; Visentin, I.; Tamietti, G. An epidemic of Gnomonia pascoe on nuts of Castanea sativa in the Cuneo area. In Proceedings of the 1st European Congress on Chestnut—Castanea 2009. Eds. G. Bounous and G.L. Beccaro. Acta Hortic. 2010, 866, 363–368. [Google Scholar] [CrossRef]
- Visentin, I.; Gentile, S.; Valentino, D.; Gonthier, P.; Tamietti, G.; Cardinale, F. Gnomoniopsis castanea sp. nov. (Gnomoniaceae, Diaper- Thales) as a causal agent of nut rot in sweet chestnut. J. Plant Pathol. 2012, 94, 411–419. [Google Scholar]
- Maresi, G.; Oliveira Longa, C.M.; Turchetti, T. Brown rot on nuts of Castanea sativa Mill: An emerging disease and its causal agent. iForest 2013, 6, 294–301. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Liew, E.C.Y.; Guest, D.I. Survey of the incidence of chestnut rot in south-eastern Australia. Australas. Plant Pathol. 2013, 42, 63–72. [Google Scholar] [CrossRef]
- Dar, M.A.; Rai, M. Biological and phylogenetic analyses evidencing the presence of Gnomoniopsis sp. in India causing canker of chestnut trees: A new report. Indian For. 2013, 139, 37–42. [Google Scholar]
- Dennert, F.G.; Broggini, G.A.L.; Gessler, C.; Storari, M. Gnomoniopsis castanea is the main agent of chestnut nut rot in Switzerland. Phytopathol. Mediterr. 2015, 54, 41–53. [Google Scholar]
- Shuttleworth, L.A.; Guest, D.I. The infection process of chestnut rot, an important disease caused by Gnomoniopsis smithogilvyi (Gnomoniaceae, Diaporthales) in Oceania and Europe. Australas. Plant Pathol. 2017, 46, 397–405. [Google Scholar] [CrossRef]
- Overy, D.P.; Seifert, K.A.; Savard, M.E.; Frisvad, J.C. Spoilage fungi and their mycotoxins in commercially marketed chestnuts. Int. J. Food Microbiol. 2003, 88, 69–77. [Google Scholar] [CrossRef]
- McKinney, H.H. Influence of soil temperature and moisture on infection of wheat seedlings by Helminthosporium sativum. J. Agric. Res. 1923, 26, 195–218. [Google Scholar]
- Rodrigues, P.; Venâncio, A.; Lima, N. Toxic reagents and expensive equipment: Are they really necessary for the extraction of good quality fungal DNA? Lett. Appl. Microbiol. 2018, 66, 32–37. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Burns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocol: A Guide to Methods and Applications; Innis, M.A., Gelfald, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Molec. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v5: User Manual/Tutorial; PRIMER-E Ltd., Ed.; Plymouth Marine Laboratory: Plymouth, UK, 2001. [Google Scholar]
- StatSoft. STATISTICA (Data Analysis Software System); Version 10; StatSoft Inc.: Tulsa, OK, USA, 2011; Available online: https://www.statistica.com/en/ (accessed on 15 October 2021).
- Costa, R.; Ribeiro, C.; Valdiviesso, T.; Afonso, S.; Costa, R.; Borges, O.; Carvalho, J.S.; Costa, H.A.; Assunção, A.; Fonseca, L.; et al. Variedades de Castanha Das Regiões Centro e Norte de Portugal; Instituto Nacional dos Recursos Biológicos: Oeiras, Portugal, 2008. [Google Scholar]
- Barreira, J.C.M.; Casal, S.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P.; Pereira, J.A. Nutritional, fatty acid and triacylglycerol profiles of Castanea sativa Mill. Cultivars: A Compositional and Chemometric Approach. J. Agric. Food Chem. 2009, 57, 2836–2842. [Google Scholar] [CrossRef]
- Vasconcelos, M.C.; Bennett, R.N.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. Industrial processing effects on chestnut fruits (Castanea sativa Mill.). 2. Crude protein, free amino acids and phenolic phytochemicals. Int. J. Food Sci. Technol. 2009, 44, 2613–2619. [Google Scholar] [CrossRef]
- El-Nagar, A.; Elzaawely, A.A.; Taha, N.A.; Nehela, Y. The antifungal activity of gallic acid and its derivatives against Alternaria solani, the causal agent of tomato early blight. Agronomy 2020, 10, 1402. [Google Scholar] [CrossRef]
- El-Shahir, A.A.; El-Wakil, D.A.; Abdel Latef, A.A.H.; Youssef, N.H. Bioactive compounds and antifungal activity of leaves and fruits methanolic extracts of Ziziphus spina-christi L. Plants 2022, 11, 746. [Google Scholar] [CrossRef]
- Wells, J.M.; Payne, J.A. Mycoflora and Market Quality of Chestnuts Treated with Hot Water to Control the Chestnut Weevil. Plant Dis. 1980, 64, 999–1001. [Google Scholar] [CrossRef]
- Morales-Rodriguez, C.; Sferrazza, I.; Aleandri, M.P.; Valle, M.D.; Mazzetto, T.; Speranza, S.; Contarini, M.; Vannini, A. Fungal community associated with adults of the Chestnut gallwasp Dryocosmus kuriphilus after emergence from galls: Taxonomy and functional ecology. Fungal Biol. 2019, 123, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Yang, S.; Litao, P.; Zeng, K.; Feng, B.; Jingjing, Y. Compositional shifts in fungal community of chestnuts during storage and their correlation with fruit quality. Postharvest Biol. Technol. 2022, 191, 111983. [Google Scholar] [CrossRef]
- Morales-Rodriguez, C.; Bastianelli, G.; Caccia, R.; Bedini, G.; Massantini, R.; Moscetti, R.; Thomidis, T.; Vannini, A. Impact of ‘brown rot’ caused by Gnomoniopsis castanea on chestnut fruits during the post-harvest process: Critical phases and proposed solutions. Fungal Biol. 2022, 102, 680–687. [Google Scholar] [CrossRef]
- Prencipe, S.; Siciliano, I.; Gatti, C.; Garibaldi, A.; Gullino, M.L.; Botta, R.; Spadaro, D. Several species of Penicillium isolated from chestnut flour processing are pathogenic on fresh chestnuts and produce mycotoxins. Food Microbiol. 2018, 76, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Ciegler, A. Fungi that produce mycotoxins: Conditions and occurrence. Mycopathologia 1978, 65, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.A.I.; de Felice, D.V.; Ianiri, G.; Pinedo-Villa, C.; De Curtis, F.; Castoria, R. Two rapid assays for screening of patulin biodegradation. Int. J. Environ. Sci. Technol. 2014, 11, 1387–1398. [Google Scholar] [CrossRef]
- Coton, M.; Dantigny, P. Mycotoxin migration in moldy foods. Curr. Opin. Food Sci. 2019, 29, 88–93. [Google Scholar] [CrossRef]
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer: New York, NY, USA, 2009; ISBN 978-1-4899-8409-8. [Google Scholar]
- Tziros, G.T.; Diamandis, S. Sclerotinia pseudotuberosa as the cause of black rot of chestnuts in Greece. J. Plant Pathol. 2018, 100, 131. [Google Scholar] [CrossRef]
- Shuttleworth, L.A.; Guest, D.I.; Liew, E.C.Y. Fungal Planet Description Sheet 107: Gnomoniopsis smithogilvyi. Persoonia 2012, 28, 142–143. [Google Scholar]
- Lewis, A.; Gorton, C.; Rees, H.; Webber, J.; Pérez-Sierra, A. First report of Gnomoniopsis smithogilvyi causing lesions and cankers of sweet chestnut in the United Kingdom. New Dis. Rep. 2017, 35, 20. [Google Scholar] [CrossRef]
- Coleine, C.; Stajich, J.E.; Pombubpa, N.; Zucconi, L.; Onofri, S.; Selbmann, L. Sampling strategies to assess microbial diversity of Antarctic cryptoendolithic communities. Polar Biol. 2020, 43, 225–235. [Google Scholar] [CrossRef]
- Khater, M.; Escosura-Muñiz, A.; Merkoçi, A. Biosensors for plant pathogen detection. Biosens. Bioelectron. 2017, 93, 72–86. [Google Scholar] [CrossRef]
- Ventura-Aguilar, R.I.; Bautista-Baños, S.; Mendoza-Acevedo, S.; Bosquez-Molina, E. Nanomaterials for designing biosensors to detect fungi and bacteria related to food safety of agricultural products. Postharvest Biol. Technol. 2023, 195, 112116. [Google Scholar] [CrossRef]
- Li, C.; Krewer, G.W.; Ji, P.; Scherm, H.; Kays, S.J. Gas sensor array for blueberry fruit disease detection and classification. Postharvest Biol. Technol. 2010, 55, 144–149. [Google Scholar] [CrossRef]
- Cinquanta, L.; Albanese, D.; De Curtis, F.; Malvano, F.; Crescitelli, A.; Di Matteo, M. Rapid assessment of gray mold (Botrytis cinerea) infection in grapes with a biosensor system. Am. J. Enol. Vitic. 2015, 66, 502–508. [Google Scholar] [CrossRef]
- Jia, W.; Liang, G.; Tian, H.; Sun, J.; Wan, C. Electronic Nose-Based Technique for Rapid Detection and Recognition of Moldy Apples. Sensors 2019, 19, 1526. [Google Scholar] [CrossRef]
- Chalupowicz, D.; Veltman, B.; Droby, S.; Eltzov, E. Evaluating the use of biosensors for monitoring of Penicillium digitatum infection in citrus fruit. Sens. Actuators B Chem. 2020, 311, 127896. [Google Scholar] [CrossRef]
- Yang, X.-H.; Li, X.-M.; Zhu, D.-H.; Zeng, Y.; Zhao, L.-Q. The diversity and dynamics of fungi in Dryocosmus kuriphilus community. Insects 2021, 12, 426. [Google Scholar] [CrossRef]
- Kačániová, M.; Sudzinová, J.; Kádasi-Horáková, M.; Valšíková, M.; Kráčmar, S. The determination of microscopic fungi from chestnut (Castanea sativa Mill.) Fruits, leaves, crust and pollen. Acta Univ. Agric. Silvic. Mendel. Brun. 2010, 58, 73–78. [Google Scholar] [CrossRef]
- Muñoz-Adalia, E.J.; Rodríguez, D.; Casado, M.; Diez, J.; Fernández, M. Fungal community of necrotic and healthy galls in chestnut trees colonized by Dryocosmus kuriphilus (Hymenoptera, Cynipidae). iForest 2019, 12, 411–417. [Google Scholar] [CrossRef]
- Aragona, M.; Haegi, A.; Valente, M.T.; Riccioni, L.; Orzali, L.; Vitale, S.; Luongo, L.; Infantino, A. New-Generation Sequencing Technology in Diagnosis of Fungal Plant Pathogens: A Dream Comes True? J. Fungi 2022, 8, 737. [Google Scholar] [CrossRef] [PubMed]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. 2022. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 20 July 2022).
- Jiang, N.; Yang, Q.; Fan, X.-L.; Tian, C.-M. Identification of six Cytospora species on Chinese chestnut in China. MycoKeys 2020, 62, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Scott, J.A.; Wong, B.; Summerbell, R.C.; Untereiner, W.A. A survey of Penicillium brevicompactum and P. bialowiezense from indoor environments, with commentary on the taxonomy of the P. brevicompactum group. Botany 2008, 86, 732–741. [Google Scholar] [CrossRef]
- Pitt, J.; Hocking, A. Fungi and Food Spoilage; Blackie Academic and Professional: London, UK, 2009. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Cheewangkoon, R.; Carnegie, A.J.; Burgess, T.I.; Summerell, B.A.; Groenewald, J.Z. Foliar pathogens of eucalypts. Stud. Mycol. 2019, 94, 125–298. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Samuels, G.J.; Ismaiel, A.; Voglmayr, H. Disentangling the Trichoderma viridescens complex. Pers. Mol. Phylogeny Evol. Fungi 2013, 31, 112–146. [Google Scholar] [CrossRef]
- Possamai, G. Podridão da Castanha em Trás-os-Montes: Caracterização Morfológica, Ecofisiológica e Molecular do Agente Causal Gnomoniopsis smithogilvyi. Master’s Dissertation, Instituto Politécnico de Bragança, Bragança, Portugal, 2020. Available online: http://hdl.handle.net/10198/23171 (accessed on 22 March 2022).









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).