
Effects of Potassium Nutrition on Corn (Zea mays L.) Physiology and Growth for Modeling


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Facility
2.2. Experimental Setup and Treatment
2.3. Trait Measurements
2.4. Data Analysis
2.5. Environmental Productivity Indices
3. Results
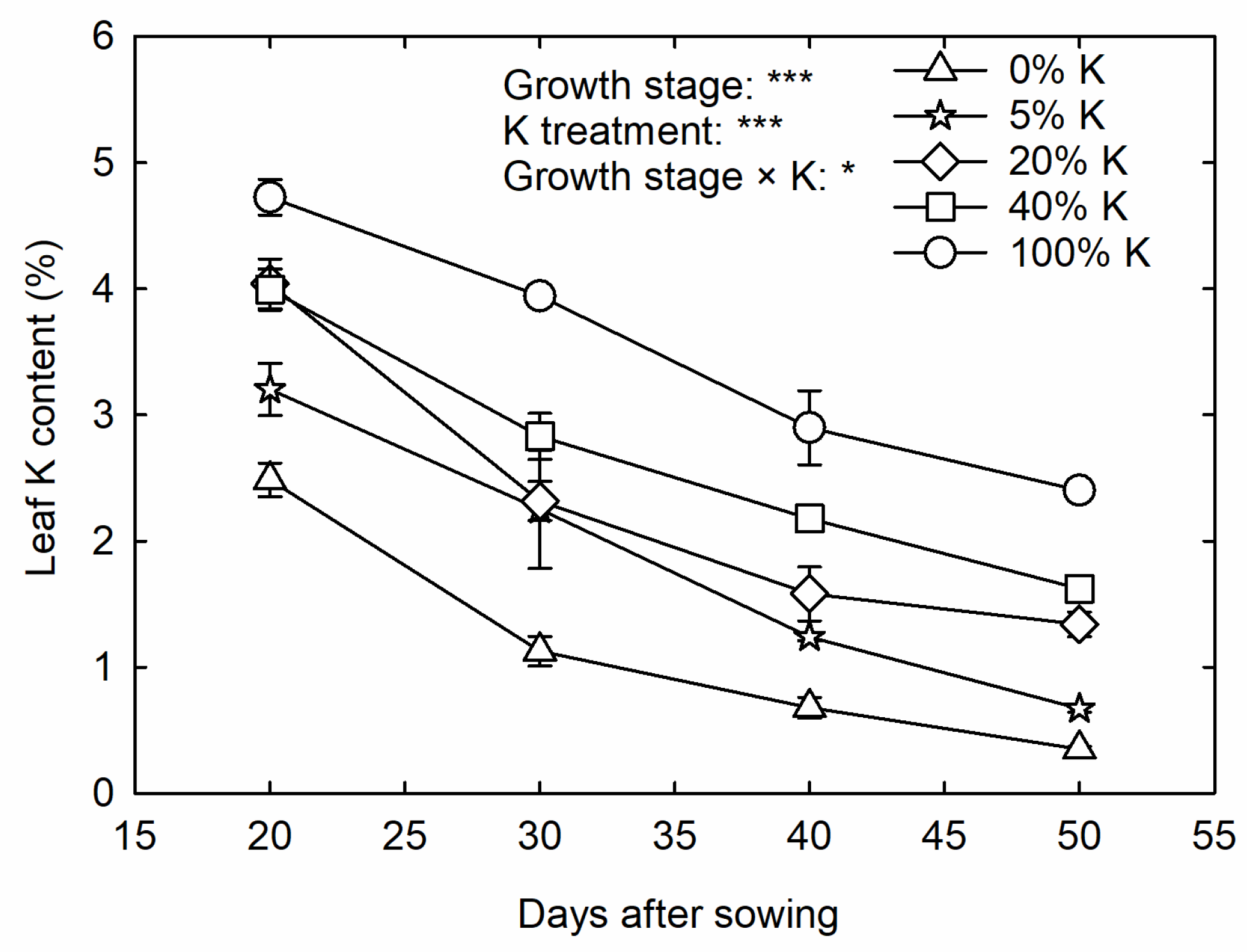
3.1. Growth Rate Influenced by K Deficiency
3.2. Effect of K Deficiency on Other Mineral Compositions
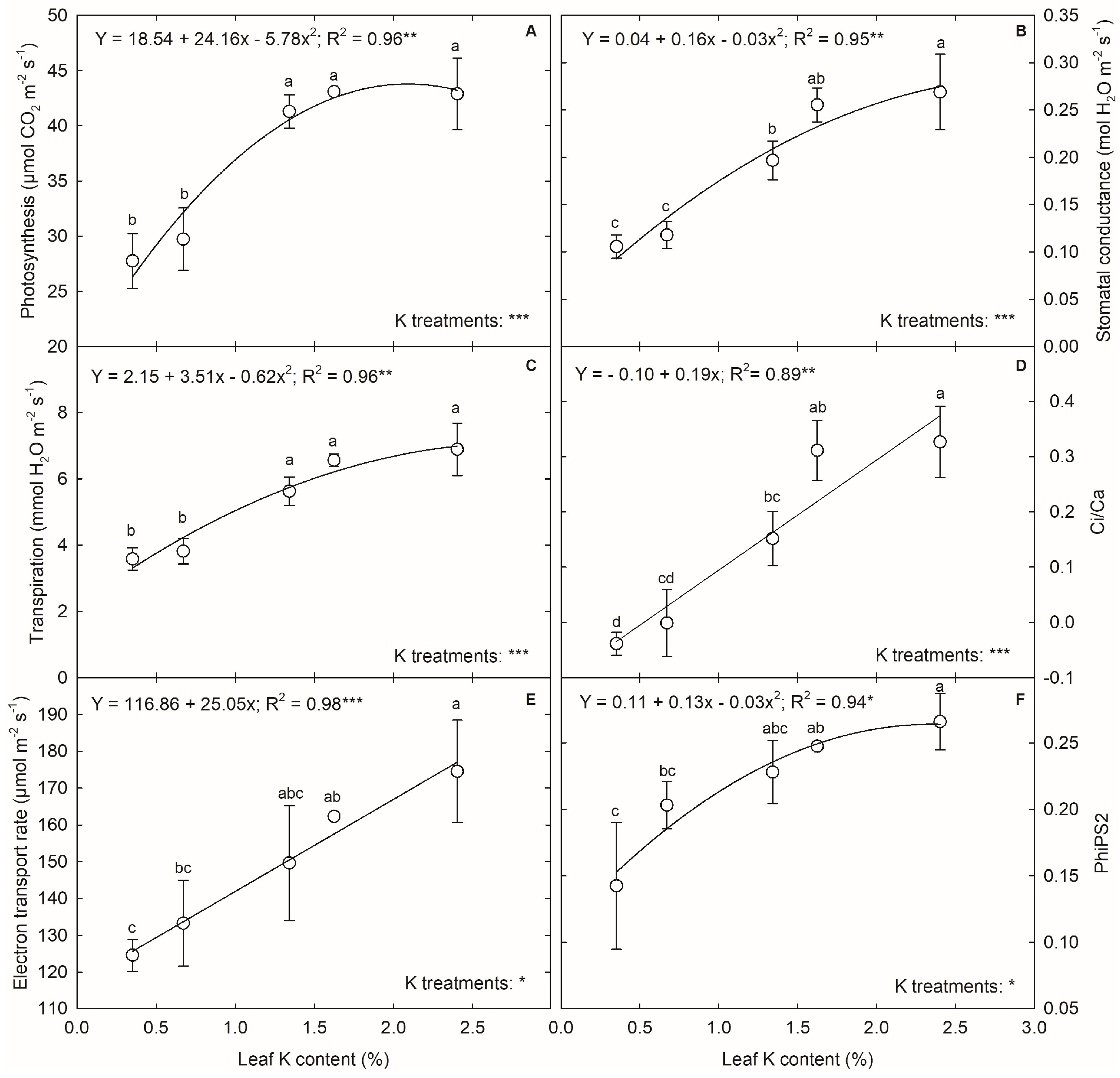
3.3. K Deficiency and Plant Physiology during the Tasseling Stage
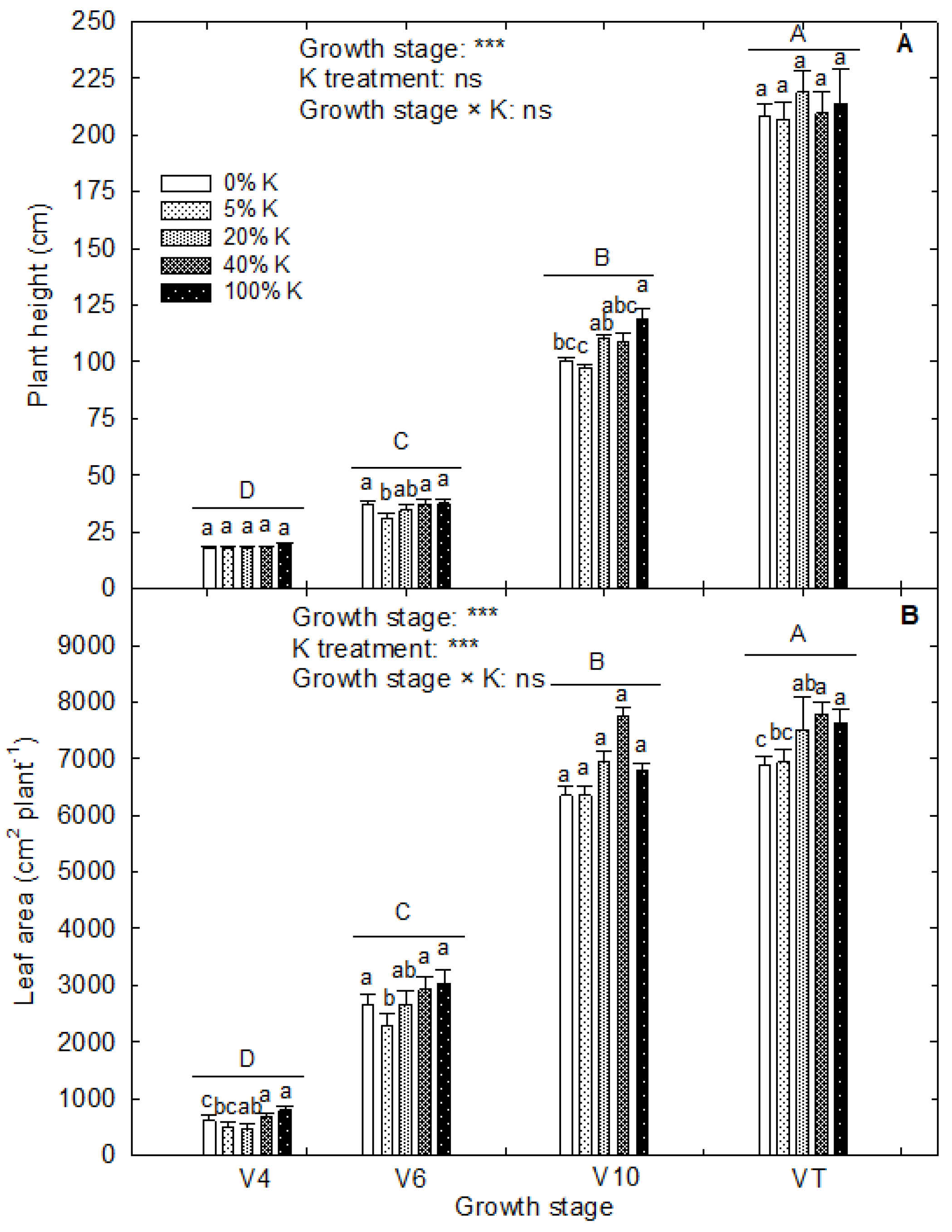
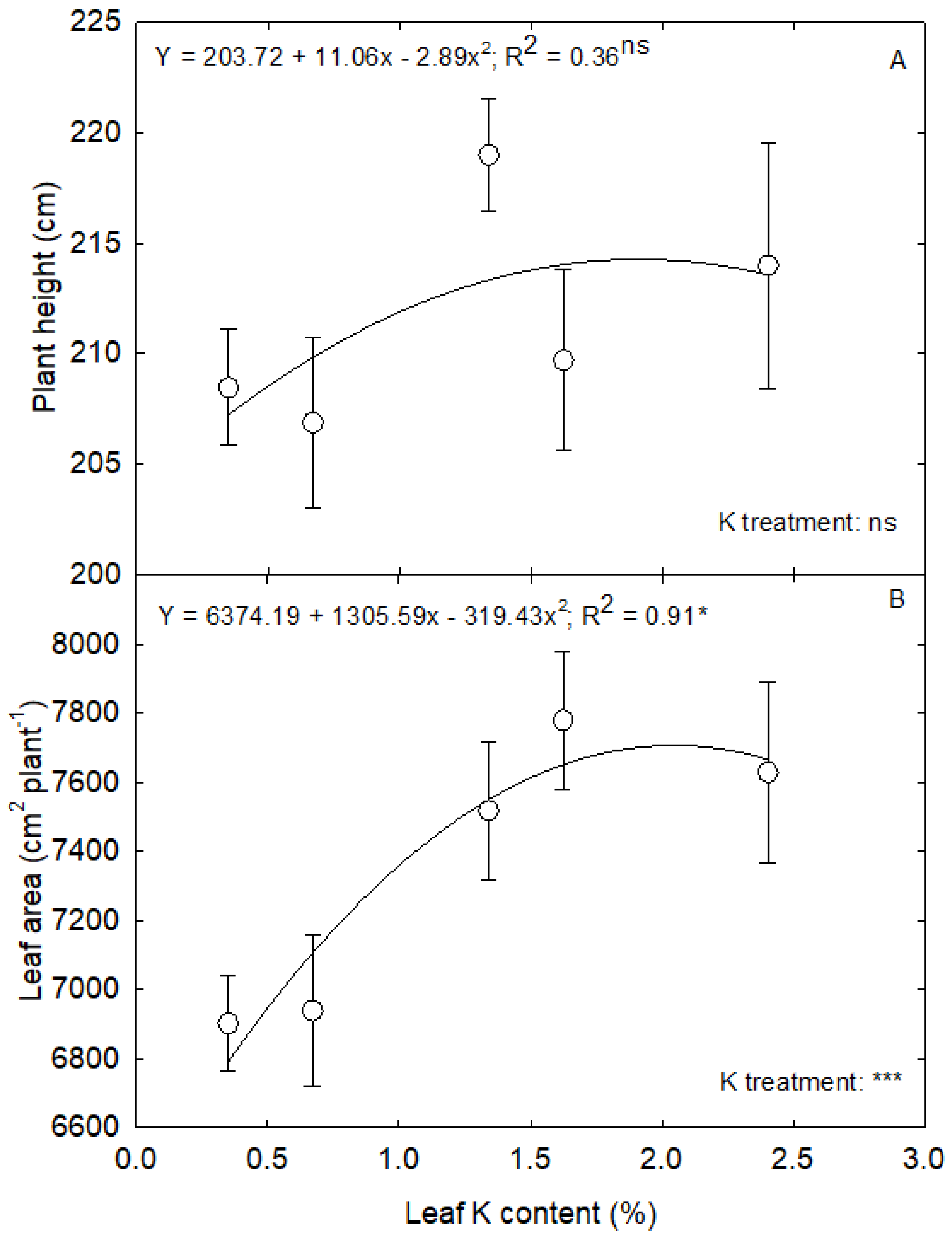
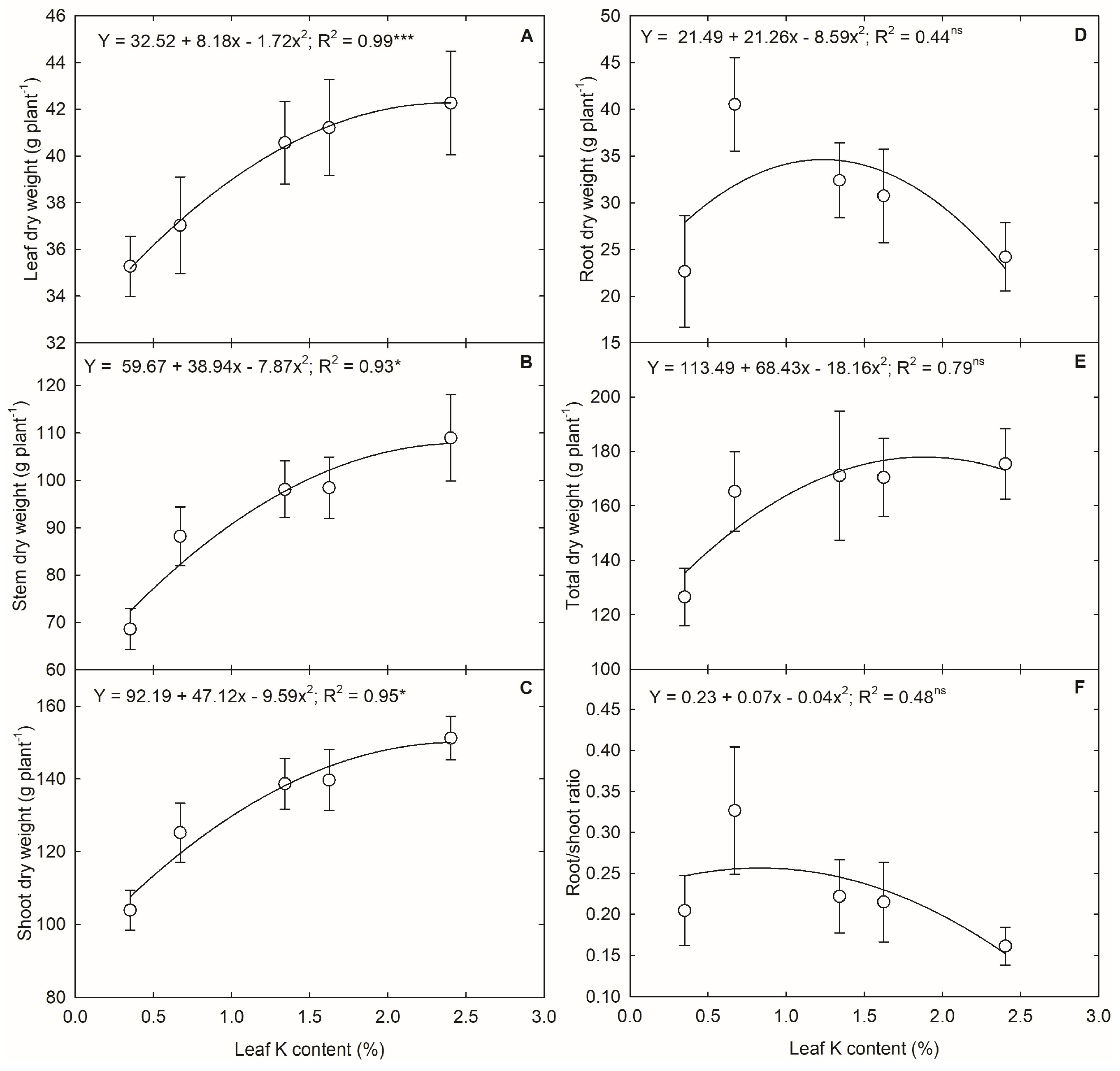
3.4. K Deficiency and Plant Growth and Biomass Accumulation
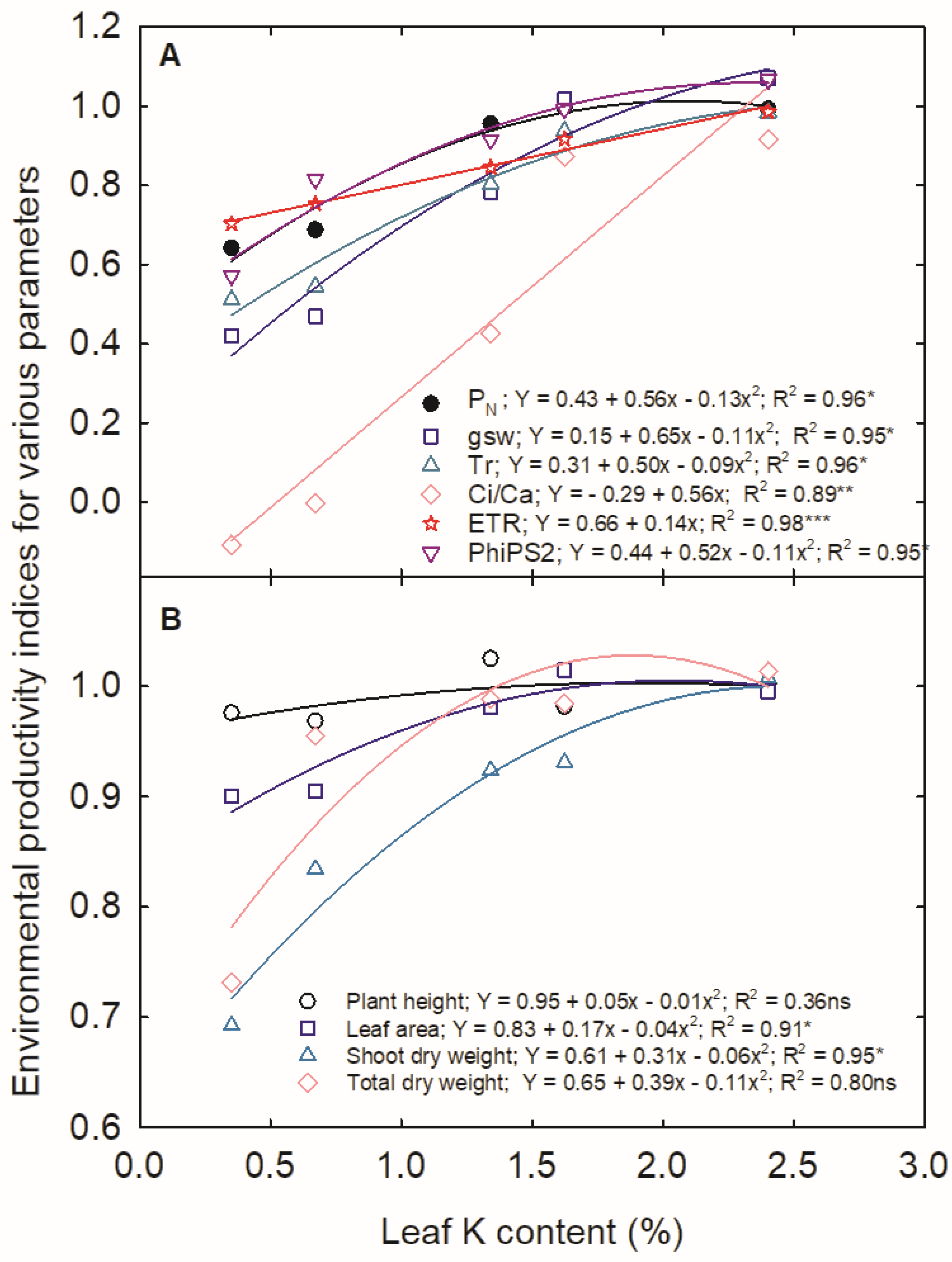
3.5. Environmental Productivity Index
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reynolds, M.; David, B.; Scott, C.C.; Robert, T.F.; Yann, M.; Diane, E.M.; Martin, A.J.P. Raising yield potential of wheat. i. overview of a consortium approach and breeding strategies. J. Exp. Bot. 2011, 62, 439–452. [Google Scholar] [CrossRef]
- Stewart, W.M.; Dibb, D.W.; Johnston, A.E.; Smyth, T.J. The contribution of commercial fertilizer nutrients to food production. Agron. J. 2005, 97, 1–6. [Google Scholar] [CrossRef]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture-status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Sparks, D.L. Potassium dynamics in soils. In Advances in Soil Science; Springer: New York, NY, USA, 1987; pp. 1–63. Available online: https://link.springer.com/book/10.1007/978-1-4612-4682-4 (accessed on 22 April 2024).
- Pingali, P.L. Green revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Crops and Livestock Products. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 23 April 2024).
- USDA-FAS. Corn Poduction in the U.S. 2014/15–2023/24. 2023, Statista. Available online: https://www.statista.com/statistics/686193/production-of-corn-us/ (accessed on 25 April 2024).
- USDA-NASS, Corn: Production by Year; Charts and Maps. 2023. Available online: https://www.nass.usda.gov/Charts_and_Maps/Field_Crops/cornprod.php (accessed on 26 April 2024).
- Oldham, L.; Jones, K.D. Nutrient Management Guidelines for Agronomic Crops Grown in Mississippi. 2023. Available online: http://extension.msstate.edu/publications/publications/nutrient-management-guidelines-for-agronomic-crops-grown-mississippi (accessed on 19 April 2024).
- Smil, V. Crop residues: Agriculture’s largest harvest. Bioscience 1999, 49, 299–308. [Google Scholar] [CrossRef]
- Dhillon, J.S.; Eickhoff, E.M.; Mullen, R.W.; Raun, W.R. World potassium use efficiency in cereal crops. Agron. J. 2019, 111, 889–896. [Google Scholar] [CrossRef]
- Hammond, H.D.; Epstein, E. Mineral nutrition in plants: Principles and perspectives. Bull. Torrey Bot. Club 1972, 99, 102. [Google Scholar] [CrossRef]
- Amtmann, A.; Stephanie, T.; Patrick, A. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant. 2008, 133, 682–691. [Google Scholar] [CrossRef]
- Andrés, Z.; Pérez-Hormaeche, J.; Leidi, E.O.; Schlücking, K.; Steinhorst, L.; McLachlan, D.H.; Schumacher, K.; Hetherington, A.M.; Kudla, J.; Cubero, B.; et al. Control of vacuolar dynamics and regulation of stomatal aperture by tonoplast potassium uptake. Proc. Natl. Acad. Sci. USA 2014, 111, E1806–E1814. [Google Scholar] [CrossRef]
- Faust, F.; Schubert, S. Protein synthesis is the most sensitive process when potassium is substituted by sodium in the nutrition of sugar beet (Beta vulgaris). Plant Physiol. Biochem. 2016, 107, 237–247. [Google Scholar] [CrossRef]
- Gerardeaux, E.; Jordan-Meille, L.; Constantin, J.; Pellerin, S.; Dingkuhn, M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ. Exp. Bot. 2010, 67, 451–459. [Google Scholar] [CrossRef]
- Hafsi, C.; Atia, A.; Lakhdar, A.; Debez, A.; Abdelly, C. Differential responses in potassium absorption and use efficiencies in the halophytes Catapodium rigidum and Hordeum maritimum to various potassium concentrations in the medium. Plant Prod. Sci. 2011, 14, 135–140. [Google Scholar] [CrossRef]
- Attia, H.; Rebah, F.; Ouhibi, C.; Saleh, M.A.; Althobaiti, A.T.; Alamer, K.H.; Nasri, M.B.; Lachaâl, M. Effect of potassium deficiency on physiological responses and anatomical structure of basil, Ocimum basilicum L. Biology 2022, 11, 1557. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Zhao, X.H.; Xia, L.; Jiang, C.J.; Wang, X.G.; Han, Y.; Wang, J.; Yu, H.Q. Effects of potassium deficiency on photosynthesis, chloroplast ultrastructure, ROS, and antioxidant activities in maize (Zea mays L.). J. Integr. Agric. 2019, 18, 395–406. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.A. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Marko, O.; Brdar, S.; Panic, M.; Lugonja, P.; Crnojevic, V. Soybean varieties portfolio optimisation based on yield prediction. Comp. Electron. Agric. 2016, 127, 467–474. [Google Scholar] [CrossRef]
- Shahhosseini, M.; Hu, G.; Huber, I.; Archontoulis, S.V. Coupling machine learning and crop modeling improves crop yield prediction in the US Corn Belt. Sci. Rep. 2021, 11, 1606. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Latham, J.; Licht, M.; Wang, L. A data-driven crop model for maize yield prediction. Commun. Biol. 2023, 6, 439. [Google Scholar] [CrossRef] [PubMed]
- Baker, C.H.; Horrocks, R.D. CORNMOD, a dynamic simulator of corn production. Agric. Sys. 1976, 1, 57–77. [Google Scholar] [CrossRef]
- Hewit, E.J. Sand and water culture methods used in the study of plant nutrition. Soil Sci. Soc. Am. J. 1953, 17, 301. [Google Scholar]
- Reddy, K.R.; Zhao, D. Interactive effects of elevated CO2 and potassium deficiency on photosynthesis, growth, and biomass partitioning of cotton. Field Crops Res. 2005, 94, 201–213. [Google Scholar] [CrossRef]
- Reddy, K.R.; Hodges, H.F.; Read, J.J.; Mckinion, J.M.; Baker, J.T.; Tarpley, L.; Reddy, V.R. Soil-plant-atmosphere-research (SPAR) facility: A tool for plant research and modeling. Biotronics 2001, 30, 27–50. [Google Scholar]
- McGeehan, S.L.; Naylor, D.V. Automated instrumental analysis of carbon and nitrogen in plant and soil samples. Commun. Soil Sci. Plant Anal. 1988, 19, 493–505. [Google Scholar] [CrossRef]
- Donohue, S.J.; Aho, D.W. Determination of P, K, Ca, Mg, Mn, Fe, Al, B, Cu, and Zn in plant tissue by inductively coupled plasma (ICP) emotion spectroscopy. In Plant Analysis Reference Procedures for the Southern Region of the United States; Plank, C.O., Ed.; Southern Cooperative Series Bulletin 368; Georgia Agricultural Experimental Station: Athens, GA, USA, 1992; pp. 37–40. [Google Scholar]
- Popat, R.; Banakara, K. Package Doebioresearch. 2022. Available online: https://CRAN.R-project.org/package=doebioresearch (accessed on 15 March 2024).
- Nobel, P.S. Productivity of Agave Deserti: Measurement by dry weight and monthly prediction using physiological responses to environmental parameters. Oecologia 1984, 64, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.R.; Kakani, V.G.; Hodges, H.F. Exploring the use of the environmental productivity index concept for crop production and modeling. In Response of Crops to Limited Water; American Society of Agronomy and Soil Science Society of America: Madison, WI, USA, 2015; pp. 387–410. [Google Scholar] [CrossRef]
- Byrant, C.J.; Jones, K.D. Potassium in Mississippi Soils. 2022. Available online: https://extension.msstate.edu/publications/potassium-mississippi-soils-0 (accessed on 26 April 2024).
- Broadley, M.R.; Escobar-gutiérrez, A.J.; Burns, A.; Burns, I.G. What are the effects of nitrogen deficiency on growth components of lettuce? New Phytol. 2000, 147, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Nleya, T.; Chungu, C.; Kleinjan, J. Corn growth and development. In iGrow Corn: Best Management Practices; Iowa State University: Ames, IA, USA, 2016; Available online: https://store.extension.iastate.edu/Product/Corn-Growth-and-Development (accessed on 20 April 2024).
- Ling, L.; Jiang, Y.; Meng, J.J.; Cai, L.M.; Cao, G.C. Phloem transport capacity of transgenic rice T1c-19 (Cry1C*) under several potassium fertilizer levels. PLoS ONE 2018, 13, e0195058. [Google Scholar] [CrossRef]
- Jordan-Meille, L.; Pellerin, S. Leaf area establishment of a maize (Zea mays L.) field crop under potassium deficiency. Plant Soil 2004, 265, 75–92. [Google Scholar] [CrossRef]
- Hu, W.; Zhao, W.; Yang, J.; Oosterhuis, D.M.; Loka, D.A.; Zhou, Z. Relationship between potassium fertilization and nitrogen metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll during the boll development stage. Plant Physiol. Biochem. 2016, 101, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; YLi, Y.; Hu, Q.; Yu, X.; Huang, M.; Zhao, J.; Dong, S.; Yuan, X.; Wen, Y. Potassium increases nitrogen and potassium utilization efficiency and yield in foxtail millet. Agronomy 2023, 13, 2200. [Google Scholar] [CrossRef]
- Niu, J.; Zhang, W.; Ru, S.; Chen, X.; Xiao, K.; Zhang, X.; Assaraf, M.; Imas, P.; Magen, H.; Zhang, F. Effects of potassium fertilization on winter wheat under different production practices in the North China plain. Field Crops Res. 2013, 140, 69–76. [Google Scholar] [CrossRef]
- Blevins, D.G.; Barnett, N.M.; Frost, W.B. Role of potassium and malate in nitrate uptake and translocation by wheat seedlings. Plant Physiol. 1978, 62, 784–788. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H.; Kirkby, E.A.; Cakmak, I. Effect of mineral nutritional status on shoot root partitioning of photoassimilates and cycling of mineral nutrients. J. Exp. Bot. 1996, 47, 1255–1263. [Google Scholar] [CrossRef]
- Talbott, L.D.; Zeiger, E. Central roles for potassium and sucrose in guard-cell osmoregulation. Plant Physiol. 1996, 111, 1051–1057. [Google Scholar] [CrossRef]
- Shabala, S. Regulation of potassium transport in leaves: From molecular to tissue level. Ann. Bot. 2003, 92, 627–634. [Google Scholar] [CrossRef]
- Lu, Z.; Lu, J.; Pan, Y.; Lu, P.; Li, X.; Cong, R.; Ren, T. Anatomical variation of mesophyll conductance under potassium deficiency has a vital role in determining leaf photosynthesis: Structural controls on Gm under K deficiency. Plant Cell Environ. 2016, 39, 2428–2439. [Google Scholar] [CrossRef]
- Tränkner, M.; Tavakol, E.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar] [CrossRef] [PubMed]
- Battie-Laclau, P.; Laclau, J.P.; Beri, C.; Mietton, L.; Muniz, M.R.A.; Arenque, B.C.; Piccolo, M.D.C.; Jordan-Meille, L.; Bouillet, J.P.; Nouvellon, Y. Photosynthetic and anatomical responses of eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell Environ. 2014, 37, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, R.; Koteyeva, N.; Voznesenskaya, E.; Evans, M.A.; Cousins, A.B.; Edwards, G.E. Coordination of leaf photosynthesis, transpiration, and structural traits in rice and wild relatives (Genus Oryza). Plant Physiol. 2013, 162, 1632–1651. [Google Scholar] [CrossRef]
- Lu, Z.; Ren, T.; Pan, Y.; Li, X.; Cong, R.; Lu, J. Differences on photosynthetic limitations between leaf margins and leaf centers under potassium deficiency for Brassica napus L. Sci. Rep. 2016, 6, 21725. [Google Scholar] [CrossRef]
- Oosterhuis, D.M.; Loka, D.A.; Raper, T.B. Potassium and stress alleviation: Physiological functions and management of cotton. J. Plant Nutr. Soil Sci. 2013, 176, 331–343. [Google Scholar] [CrossRef]
- Carroll, M.J.; Slaughter, L.H.; Krouse, J.M. Turgor potential and osmotic constituents of Kentucky bluegrass leaves supplied with four levels of potassium. Agron. J. 1994, 86, 1079–1083. [Google Scholar] [CrossRef]
- Martin, T.A.; Hinckley, T.M.; Meinzer, F.C.; Sprugel, D.G. Boundary layer conductance, leaf temperature and transpiration of Abies amabilis branches. Tree Physiol. 1999, 19, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Schuepp, P.H. Tansley Review No. 59. Leaf boundary layers. New Phytol. 1993, 125, 477–507. [Google Scholar] [CrossRef] [PubMed]
- Woodrow, I.E.; Ball, J.T.; Berry, J.A. Control of photosynthetic carbon-dioxide fixation by the boundary-layer, stomata and ribulose 1,5-biphosphate carboxylase oxygenase. Plant Cell Environ. 1990, 13, 339–347. [Google Scholar] [CrossRef]
- Havlin, J.L.; Beaton, J.D.; Tisdale, S.L.; Nelson, W.L. Soil Fertility and Fertilizers: An Introduction to Nutrient Management, 7th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2005. [Google Scholar]
- Oneill, S.D.; Spanswick, R.M. Characterization of native and reconstituted plasma membrane H+-ATPase from the plasma membrane of Beta vulgaris. J. Membr. Biol. 1984, 79, 245–256. [Google Scholar] [CrossRef]
- Takahashi, S.; Badger, M.R. Photoprotection in plants: A new light on photosystem II damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Kafkafi, U. The functions of plant K in overcoming environmental stress situations. In Proceedings of the 22nd Colloquium of the International Potash Institute, Soligorsk, USSR, 18–23 June 1990; International Potash Institute: Bern, Switzerland, 1990; pp. 81–93. [Google Scholar]
- Münch, E. Die stoffbewegungen in der pflanze (Gustav Fischer, jena). Curr. Opin. Plant Biol. 1930, 43, 36–42. [Google Scholar]
- Baker, R.F.; Leach, K.A.; Boyer, N.R.; Swyers, M.J.; Alfonso, Y.B.; Skopelitis, T.; Luo, A.; Sylvester, A.; Jackson, D.; Braun, D.M. Sucrose transporter ZmSut1 expression and localization uncover new insights into sucrose phloem loading. Plant Physiol. 2016, 172, 1876–1898. [Google Scholar] [CrossRef] [PubMed]
- Philippar, K.; Büchsenschütz, K.; Abshagen, M.; Fuchs, I.; Geiger, D.; Lacombe, B.; Hedrich, R. The K+ channel KZM1 mediates potassium uptake into the phloem and guard cells of the C4 grass Zea mays. J. Biol. Chem. 2003, 278, 16973–16981. [Google Scholar] [CrossRef]
- Atkins, C.A.; Smith, P.M.C. Translocation in legumes: Assimilates, nutrients, and signaling molecules1. Plant Physiol. 2007, 144, 550–561. [Google Scholar] [CrossRef]
- Chen, J.; Gabelman, W.H. Morphological and physiological characteristics of tomato roots associated with potassium-acquisition efficiency. Sci. Hortic. 2000, 83, 213–225. [Google Scholar] [CrossRef]
- Jones, C.A.; Kiniry, J.R. CERES-Corn: A Simulation of Corn Growth and Development; Texas A&M Univ. Press: College Station, TX, USA, 1986. [Google Scholar]
- Lizaso, J.I.; Batchelor, W.D.; Westgate, M.E. A leaf area model to simulate cultivar-specific expansion and senescence of corn leaves. Field Crops Res. 2003, 80, 1–17. [Google Scholar] [CrossRef]
- Yang, Y.; Timlin, D.J.; Fleisher, D.H.; Kim, S.H.; Quebedeaux, B.; Reddy, V.R. Simulating leaf area of corn plants at contrasting water status. Agric. For. Meteorol. 2009, 149, 1161–1167. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mineral (%) | Growth Stage | Potassium (K) | Growth Stage × K |
|---|---|---|---|
| Nitrogen | *** | * | ns |
| Phosphorus | *** | *** | * |
| Potassium | *** | *** | ns |
| Calcium | *** | *** | ns |
| Magnesium | *** | *** | *** |
| Sulfur | *** | *** | *** |
| Zinc | *** | ns | ns |
| Boron | *** | * | ** |
| Iron | *** | ns | ** |
| Manganese | *** | *** | *** |
| Copper | *** | *** | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thenveettil, N.; Reddy, K.N.; Reddy, K.R. Effects of Potassium Nutrition on Corn (Zea mays L.) Physiology and Growth for Modeling. Agriculture 2024, 14, 968. https://doi.org/10.3390/agriculture14070968
Thenveettil N, Reddy KN, Reddy KR. Effects of Potassium Nutrition on Corn (Zea mays L.) Physiology and Growth for Modeling. Agriculture. 2024; 14(7):968. https://doi.org/10.3390/agriculture14070968
Chicago/Turabian StyleThenveettil, Naflath, Krishna N. Reddy, and Kambham Raja Reddy. 2024. "Effects of Potassium Nutrition on Corn (Zea mays L.) Physiology and Growth for Modeling" Agriculture 14, no. 7: 968. https://doi.org/10.3390/agriculture14070968
APA StyleThenveettil, N., Reddy, K. N., & Reddy, K. R. (2024). Effects of Potassium Nutrition on Corn (Zea mays L.) Physiology and Growth for Modeling. Agriculture, 14(7), 968. https://doi.org/10.3390/agriculture14070968