

Sulfate Nutrition Modulates the Oxidative Response against Short-Term Al3+-Toxicity Stress in Lolium perenne cv. Jumbo Shoot Tissues
 , and
, and
Abstract
:
1. Introduction
2. Material and Methods
2.1. Experimental Growth Conditions and Plant Leaves Tissue Collection
2.2. Aluminum, Calcium, and Magnesium Estimation in Plant Leaves via Atomic Absorption Spectroscopy (AAS)
2.3. Sulfur Estimation in L. perenne Leaves via Gravimetry
2.4. Lipid Peroxidation Measurements
2.5. Ascorbic Acid and Dehydroascorbic Acid Quantitation
2.6. Quantitation of Oxidized (GSSG) and Reduced (GSH) Glutathione in Leaves of L. perenne
2.7. Protein Extraction from L. perenne Leaf Tissue and Calculation of ROS Enzymatic Activities
2.8. Total RNA Extraction from Leaves of Lolium perenne and qRT-PCR Assays
2.9. Statistical Analysis
3. Results
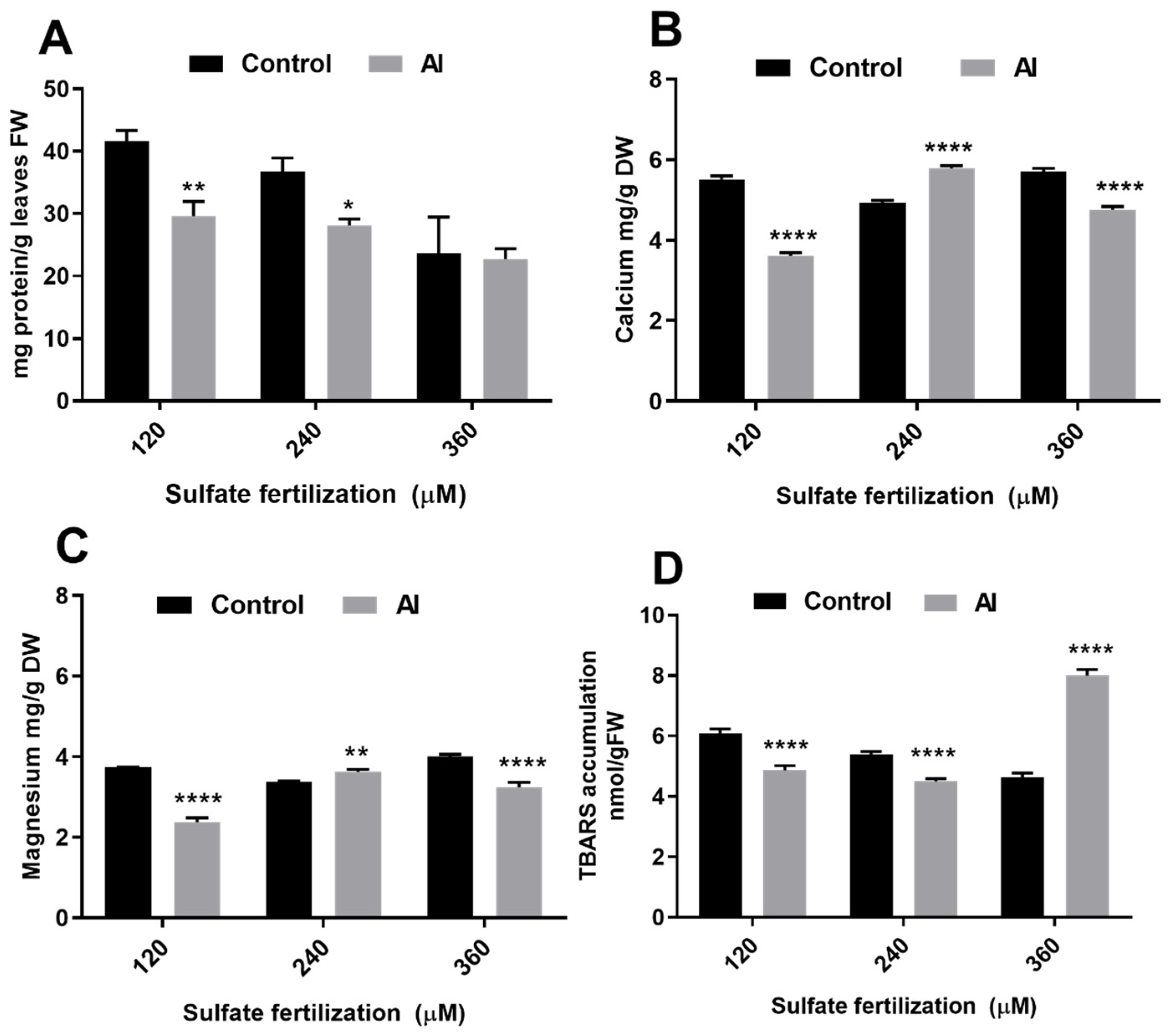
3.1. Sulfate Supplementation Modulates Total Proteins, Calcium, Magnesium and TBARs Concentration in Leaves of Plants Subjected to Al3+ Toxicity
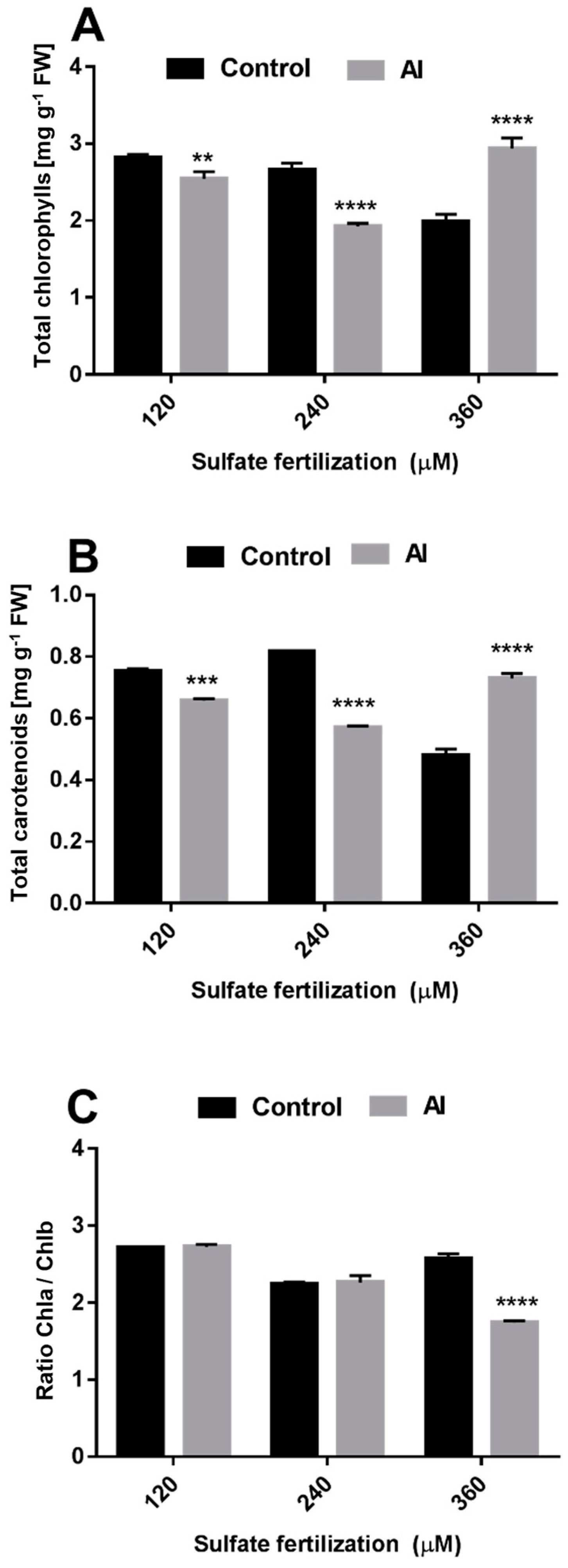
3.2. Sulfate Supplementation Modulated Photosynthetic Response in Leaves of Plants Subjected to Al3+ Toxicity
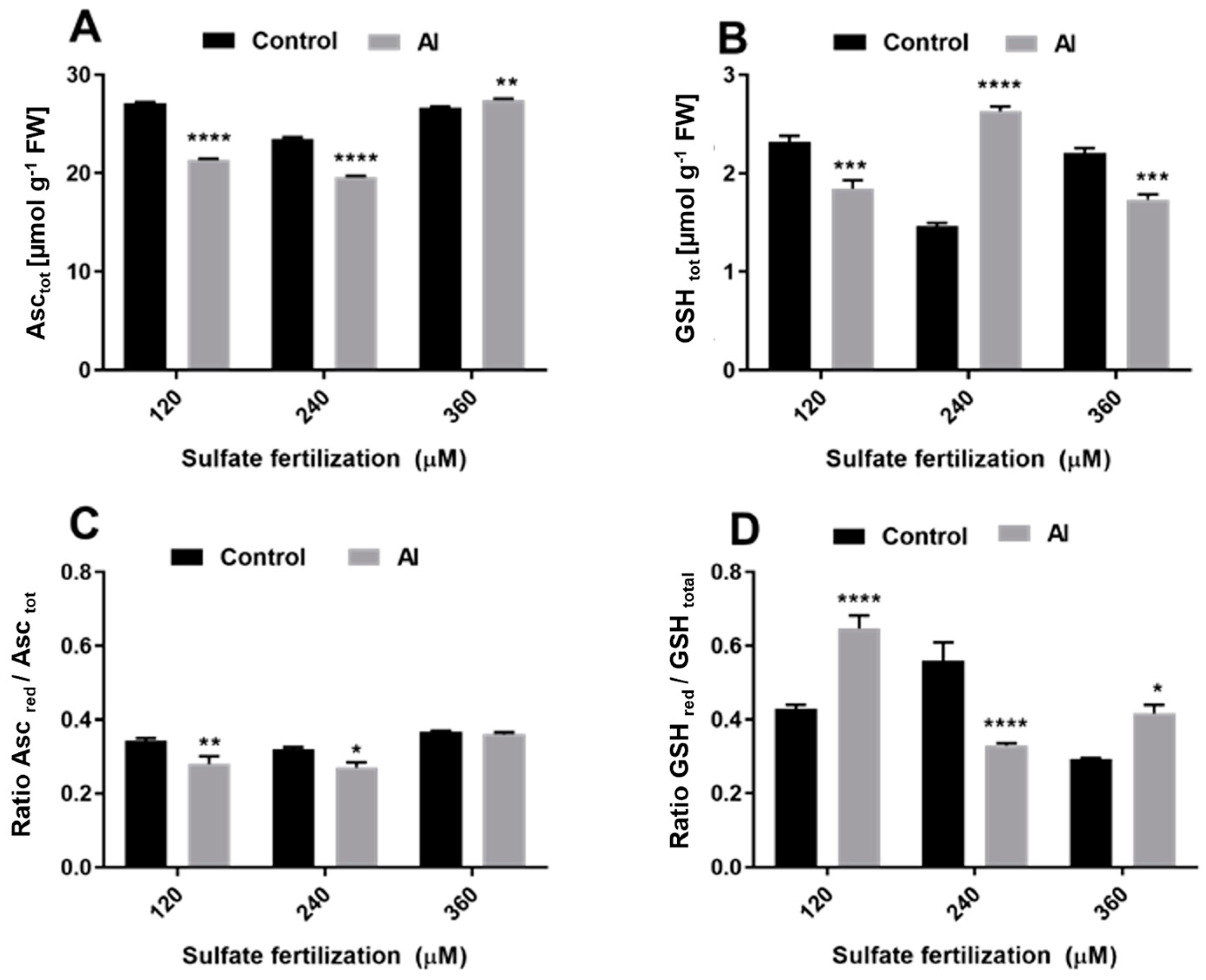
3.3. Sulfate Supply Modulates Antioxidant Metabolites in Shoots of L. perenne cv. Jumbo Exposed to Al3+ Toxicity
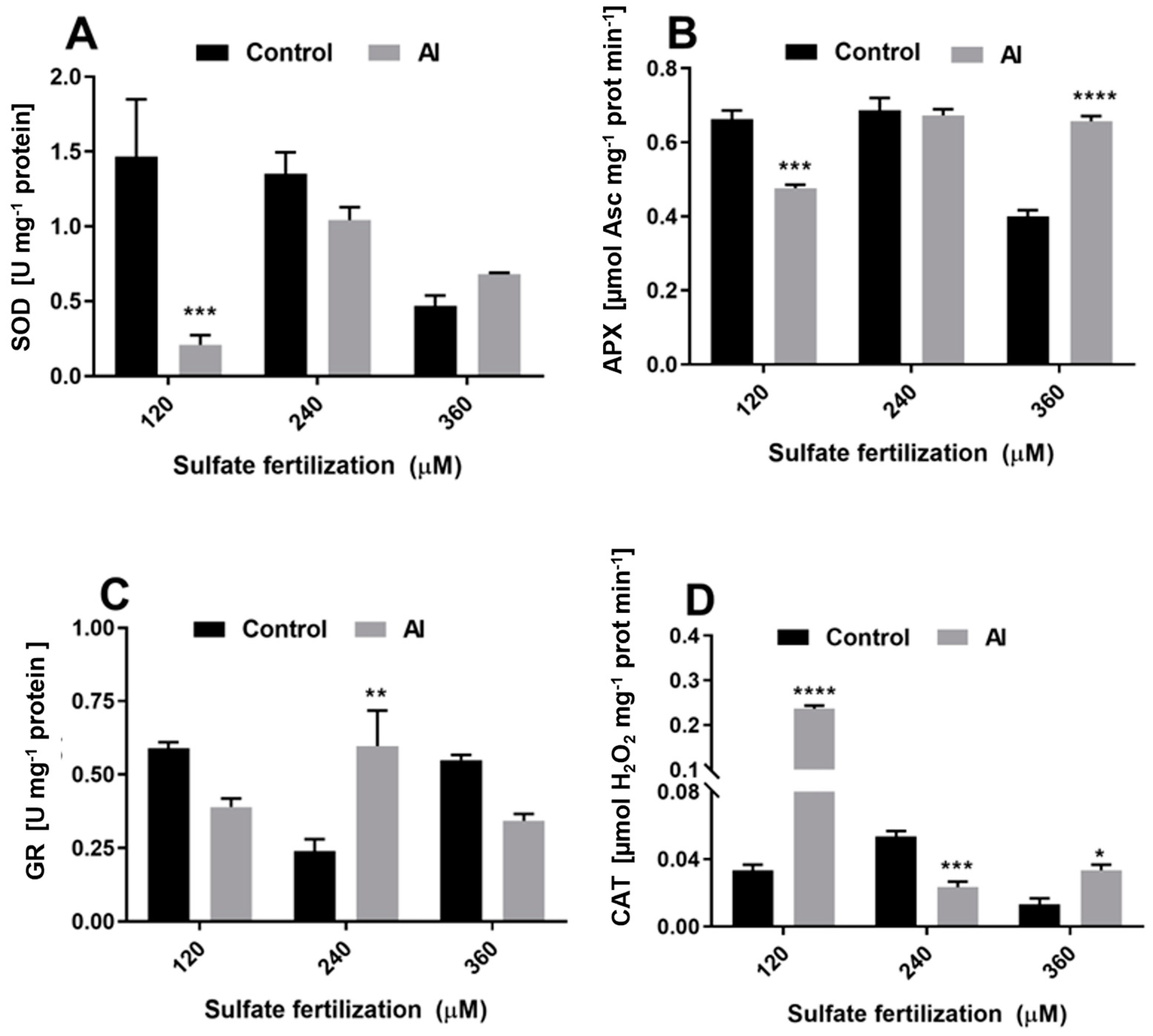
3.4. Sulfate Supplementation Modulates Enzyme Activity and Gene Expression of ROS Scavenging Enzymes in Shoots of L. perenne cv. Jumbo Exposed to Al3+ Toxicity
4. Discussion
4.1. Adequate Sulfate Supply Recovered Membrane Damage, and Mineral Uptake; but Not Total Protein Amount after Al3+ Toxicity Exposure
4.2. Adequate Sulfate Supplementation Was Able to Maintain Photosynthetic Parameters in Leaves of L. perennial cv. Jumbo Exposed to Al3+ Toxicity
4.3. Sulfate Supplementation Activates ROS Defense Mechanism in Leaves of L. perennial cv. Jumbo Exposed to Al3+ Toxicity
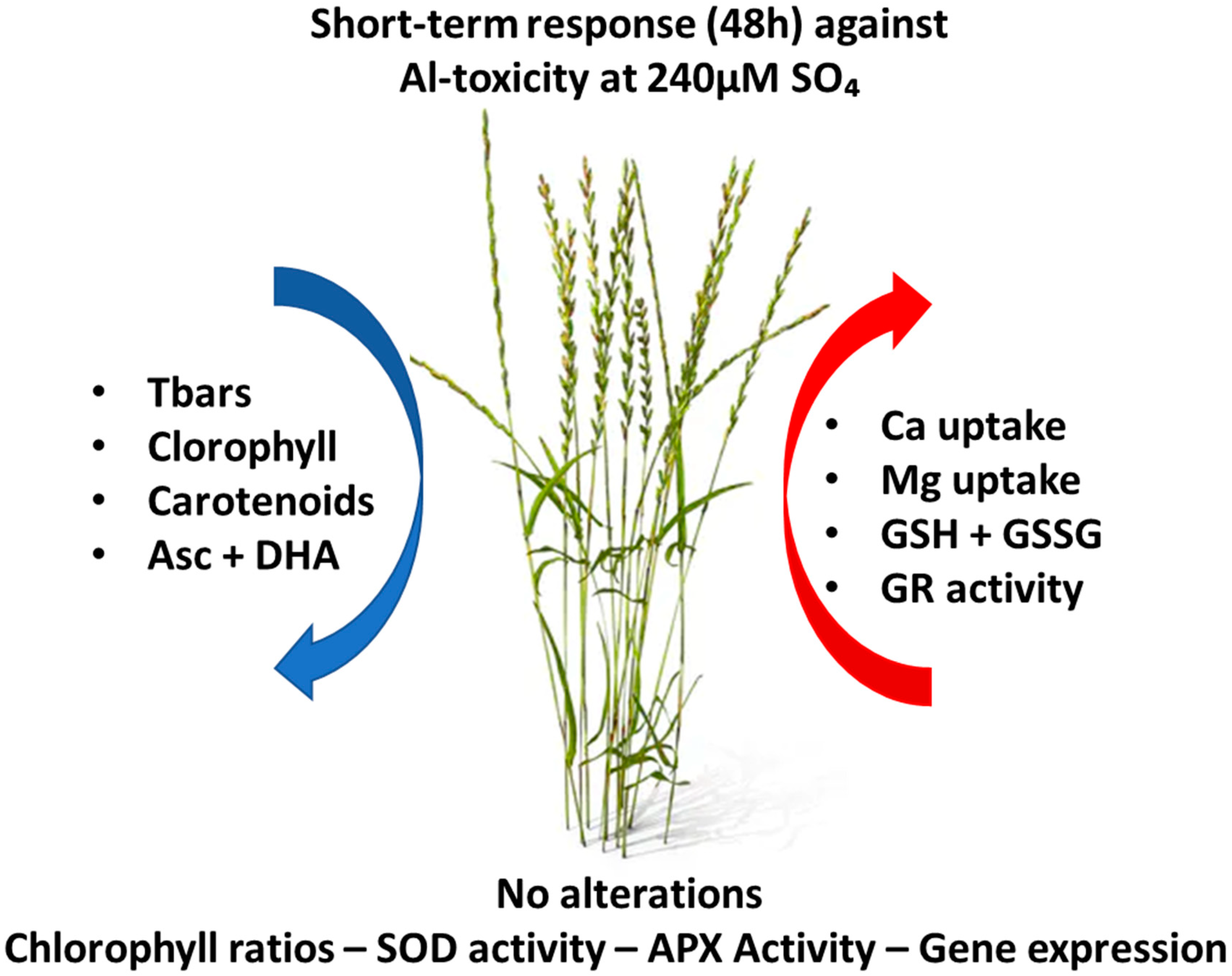
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Uexküll, H.R.; Mutert, E. Global Extent, Development and Economic Impact of Acid Soils. Plant Soil 1995, 171, 1–15. [Google Scholar] [CrossRef]
- Raza, S.; Zamanian, K.; Ullah, S.; Kuzyakov, Y.; Virto, I.; Zhou, J. Inorganic Carbon Losses by Soil Acidification Jeopardize Global Efforts on Carbon Sequestration and Climate Change Mitigation. J. Clean. Prod. 2021, 315, 128036. [Google Scholar] [CrossRef]
- Nachtergaele, F. Soil Taxonomy—A Basic System of Soil Classification for Making and Interpreting Soil Surveys. Geoderma 2001, 99, 336–337. [Google Scholar] [CrossRef]
- Aguilera, S.M.; Borie, G.; Peirano, P.; Galindo, G. Organic Matter in Volcanic Soils in Chile: Chemical and Biochemical Characterization. Commun. Soil Sci. Plant Anal. 1997, 28, 899–912. [Google Scholar] [CrossRef]
- Shetty, R.; Vidya, C.S.N.; Prakash, N.B.; Lux, A.; Vaculík, M. Aluminum Toxicity in Plants and Its Possible Mitigation in Acid Soils by Biochar: A Review. Sci. Total Environ. 2021, 765, 142744. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y. Aluminum Toxicity in Plant Cells: Mechanisms of Cell Death and Inhibition of Cell Elongation. Soil Sci. Plant Nutr. 2019, 65, 41–55. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Gunsé, B.; Corrales, I.; Barceló, J. A Glance into Aluminum Toxicity and Resistance in Plants. Sci. Total Environ. 2008, 400, 356–368. [Google Scholar] [CrossRef]
- Matsumoto, H. Cell Biology of Aluminum Toxicity Tolerance in Higher Plants. Int. Rev. Cytol. 2000, 200, 1–46. [Google Scholar]
- Llugany, M.; Poschenrieder, C.; Barceló, J. Monitoring of Aluminium-induced Inhibition of Root Elongation in Four Maize Cultivars Differing in Tolerance to Aluminium and Proton Toxicity. Physiol. Plant. 1995, 93, 265–271. [Google Scholar] [CrossRef]
- Silva, I.R.; Smyth, T.J.; Moxley, D.F.; Carter, T.E.; Allen, N.S.; Rufty, T.W. Aluminum Accumulation at Nuclei of Cells in the Root Tip. Fluorescence Detection Using Lumogallion and Confocal Laser Scanning Microscopy. Plant Physiol. 2000, 123, 543–552. [Google Scholar] [CrossRef]
- He, H.; Li, Y.; He, L.F. Aluminum Toxicity and Tolerance in Solanaceae Plants. S. Afr. J. Bot. 2019, 123, 23–29. [Google Scholar] [CrossRef]
- Cartes, P.; McManus, M.; Wulff-Zottele, C.; Leung, S.; Gutiérrez-Moraga, A.; Mora, M. de la L. Differential Superoxide Dismutase Expression in Ryegrass Cultivars in Response to Short Term Aluminium Stress. Plant Soil 2012, 350, 353–363. [Google Scholar] [CrossRef]
- Frantzios, G.; Galatis, B.; Apostolakos, P. Aluminium Effects on Microtubule Organization in Dividing Root-Tip Cells of Triticum Turgidum. II. Cytokinetic Cells. J. Plant Res. 2001, 114, 157–170. [Google Scholar] [CrossRef]
- Horst, W.J.; Wang, Y.; Eticha, D. The Role of the Root Apoplast in Aluminium-Induced Inhibition of Root Elongation and in Aluminium Resistance of Plants: A Review. Ann. Bot. 2010, 106, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Mano, J.; Wang, S.; Tsuji, W.; Tanaka, K. The Involvement of Lipid Peroxide-Derived Aldehydes in Aluminum Toxicity of Tobacco Roots. Plant Physiol. 2010, 152, 1406–1417. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Kobayashi, Y.; Matsumoto, H. Lipid Peroxidation Is an Early Symptom Triggered by Aluminum, but Not the Primary Cause of Elongation Inhibition in Pea Roots. Plant Physiol. 2001, 125, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kobayashi, Y.; Devi, S.R.; Rikiishi, S.; Matsumoto, H. Aluminum Toxicity Is Associated with Mitochondrial Dysfunction and the Production of Reactive Oxygen Species in Plant Cells. Plant Physiol. 2002, 128, 63–72. [Google Scholar] [CrossRef]
- Wulff-Zottele, C.; Hesse, H.; Fisahn, J.; Bromke, M.; Vera-Villalobos, H.; Li, Y.; Frenzel, F.; Giavalisco, P.; Ribera-Fonseca, A.; Zunino, L.; et al. Sulphate Fertilization Ameliorates Long-Term Aluminum Toxicity Symptoms in Perennial Ryegrass (Lolium perenne). Plant Physiol. Biochem. 2014, 83, 88–99. [Google Scholar] [CrossRef]
- Reyes-Díaz, M.; Inostroza-Blancheteau, C.; Millaleo, R.; Cruces, E.; Wulff-Zottele, C.; Alberdi, M.; de la Luz Mora, M. Long-Term Aluminum Exposure Effects on Physiological and Biochemical Features of Highbush Blueberry Cultivars. J. Am. Soc. Hortic. Sci. 2010, 135, 212–222. [Google Scholar] [CrossRef]
- Reyes-Diaz, M.; Alberdi, M.; Mora, M.D.L.L. Short-Term Aluminum Stress Differentially Affects the Photochemical Efficiency of Photosystem II in Highbush Blueberry Genotypes. J. Am. Soc. Hortic. Sci. 2009, 134, 14–21. [Google Scholar] [CrossRef]
- Cheng, X.; Fang, T.; Zhao, E.; Zheng, B.; Huang, B.; An, Y.; Zhou, P. Protective Roles of Salicylic Acid in Maintaining Integrity and Functions of Photosynthetic Photosystems for Alfalfa (Medicago sativa L.) Tolerance to Aluminum Toxicity. Plant Physiol. Biochem. 2020, 155, 570–578. [Google Scholar] [CrossRef]
- Phukunkamkaew, S.; Tisarum, R.; Pipatsitee, P.; Samphumphuang, T.; Maksup, S.; Cha-um, S. Morpho-Physiological Responses of Indica Rice (Oryza sativa Sub. indica) to Aluminum Toxicity at Seedling Stage. Environ. Sci. Pollut. Res. 2021, 28, 29321–29331. [Google Scholar] [CrossRef]
- Ofoe, R.; Thomas, R.H.; Asiedu, S.K.; Wang-Pruski, G.; Fofana, B.; Abbey, L. Aluminum in Plant: Benefits, Toxicity and Tolerance Mechanisms. Front. Plant Sci. 2023, 13, 1085998. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Li, Q.; Qi, Y.P.; Yang, L.T.; Ye, X.; Chen, H.H.; Chen, L.S. Sulfur-Mediated-Alleviation of Aluminum-Toxicity in Citrus Grandis Seedlings. Int. J. Mol. Sci. 2017, 18, 2570. [Google Scholar] [CrossRef] [PubMed]
- Ritchey, K.D.; Snuffer, J.D. Limestone, Gypsum, and Magnesium Oxide Influence Restoration of an Abandoned Appalachian Pasture. Agron. J. 2002, 94, 830–839. [Google Scholar] [CrossRef]
- Demanet-Fillipi, R. Enmiendas Calcareas. Plant Lechero Watt´s. Chile. Bulletin N°2. April 2017. Available online: https://www.watts.cl/docs/default-source/charlas-a-productores/enmienda-calcarea.pdf?sfvrsn=4 (accessed on 30 October 2023).
- Meriño-Gergichevich, C.; Alberdi, M.; Ivanov, A.G.; Reyes-Díaz, M. Al3+-Ca2+ Interaction in Plants Growing in Acid Soils: Al-Phytotoxicity Response to Calcareous Amendments. J. Soil Sci. Plant Nutr. 2010, 10, 217–243. [Google Scholar]
- Toma, M.; Sumner, M.E.; Weeks, G.; Saigusa, M. Long-term Effects of Gypsum on Crop Yield and Subsoil Chemical Properties. Soil Sci. Soc. Am. J. 1999, 63, 891–895. [Google Scholar] [CrossRef]
- Takahashi, T.; Ikeda, Y.; Nakamura, H.; Nanzyo, M. Efficiency of Gypsum Application to Acid Andosols Estimated Using Aluminum Release Rates and Plant Root Growth. Soil Sci. Plant Nutr. 2006, 52, 584–592. [Google Scholar] [CrossRef]
- Kinraide, T.B. Identity of the Rhizotoxic Aluminium Species. Plant Soil 1991, 134, 167–178. [Google Scholar] [CrossRef]
- Mora, M.L.; Cartes, P.; Demanet, R.; Cornforth, I.S. Effects of Lime and Gypsum on Pasture Growth and Composition on an Acid Andisol in Chile, South America. Commun. Soil Sci. Plant Anal. 2002, 33, 2069–2081. [Google Scholar] [CrossRef]
- Vera-Villalobos, H.; Lunario-Delgado, L.; Pérez-Retamal, D.; Román, D.; Leiva, J.C.; Zamorano, P.; Mercado-Seguel, A.; Gálvez, A.S.; Benito, C.; Wulff-Zottele, C. Sulfate Nutrition Improves Short-Term Al3+-Stress Tolerance in Roots of Lolium perenne L. Plant Physiol. Biochem. 2020, 148, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Kopriva, S.; Giordano, M.; Saito, K.; Hell, R. Sulfur Assimilation in Photosynthetic Organisms: Molecular Functions and Regulations of Transporters and Assimilatory Enzymes. Annu. Rev. Plant Biol. 2011, 62, 157–184. [Google Scholar] [CrossRef]
- Saito, K. Sulfur Assimilatory Metabolism. The Long and Smelling Road. Plant Physiol. 2004, 136, 2443–2450. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.Q.; Zhang, J.H.; Sun, L.M.; Zhu, L.F.; Abliz, B.; Hu, W.J.; Zhong, C.; Bai, Z.G.; Sajid, H.; Cao, X.C.; et al. Hydrogen Sulfide Alleviates Aluminum Toxicity via Decreasing Apoplast and Symplast al Contents in Rice. Front. Plant Sci. 2018, 9, 294. [Google Scholar] [CrossRef]
- Alarcón-Poblete, E.; Inostroza-Blancheteau, C.; Alberdi, M.; Rengel, Z.; Reyes-Díaz, M. Molecular Regulation of Aluminum Resistance and Sulfur Nutrition during Root Growth. Planta 2018, 247, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.J.; Foy, C.D. Mechanisms of Aluminum Tolerance in Triticum aestivum L. (Wheat). I. Differential PH Induced by Winter Cultivars in Nutrient Solutions. Am. J. Bot. 1985, 72, 695–701. [Google Scholar] [CrossRef]
- Shaff, J.E.; Schultz, B.A.; Craft, E.J.; Clark, R.T.; Kochian, L.V. GEOCHEM-EZ: A Chemical Speciation Program with Greater Power and Flexibility. Plant Soil 2010, 330, 207–214. [Google Scholar] [CrossRef]
- Román-Silva, D.A.; Rivera, L.; Morales, T.; Ávila, J.; Cortés, P. Determination of Trace Elements in Environmental and Biological Samples Using Improved Sample Introduction in Flame Atomic Absorption Spectrometry (HHPN-AAS; HHPN-FF-AAS). Int. J. Environ. Anal. Chem. 2003, 83, 327–341. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified Thiobarbituric Acid Assay for Measuring Lipid Oxidation in Sugar-Rich Plant Tissue Extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Gillespie, K.M.; Ainsworth, E.A. Measurement of Reduced, Oxidized and Total Ascorbate Content in Plants. Nat. Protoc. 2007, 2, 871–874. [Google Scholar] [CrossRef]
- Rahman, I.; Kode, A.; Biswas, S.K. Assay for Quantitative Determination of Glutathione and Glutathione Disulfide Levels Using Enzymatic Recycling Method. Nat. Protoc. 2007, 1, 3159–3165. [Google Scholar] [CrossRef] [PubMed]
- Donahue, J.L.; Okpodu, C.M.; Cramer, C.L.; Grabau, E.A.; Alscher, R.G. Responses of Antioxidants to Paraquat in Pea Leaves: Relationships to Resistance. Plant Physiol. 1997, 113, 249–257. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide Is Scavenged by Ascorbate-Specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Ambuko, J.; Carmel Zanol, G.; Sekozawa, Y.; Sugaya, S.; Gemma, H. Reactive Oxygen Species (ROS) Scavenging in Hot Air Preconditioning Mediated Alleviation of Chilling Injury in Banana Fruits. J. Agric. Sci. 2012, 5, 319. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Ruijter, J.M.; Benes, V.; van den Hoff, M.J.B. Web-Based LinRegPCR: Application for the Visualization and Analysis of (RT)-QPCR Amplification and Melting Data. BMC Bioinform. 2021, 22, 398. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]
- Chen, W.W.; Tang, L.; Wang, J.Y.; Zhu, H.H.; Jin, J.F.; Yang, J.L.; Fan, W. Research Advances in the Mutual Mechanisms Regulating Response of Plant Roots to Phosphate Deficiency and Aluminum Toxsicity. Int. J. Mol. Sci. 2022, 23, 1137. [Google Scholar] [CrossRef]
- Correia, S.; Matos, M.; Ferreira, V.; Martins, N.; Gonçalves, S.; Romano, A.; Pinto-Carnide, O. Molecular Instability Induced by Aluminum Stress in Plantago Species. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2014, 770, 105–111. [Google Scholar] [CrossRef]
- Yang, M.; Tan, L.; Xu, Y.; Zhao, Y.; Cheng, F.; Ye, S.; Jiang, W. Effect of Low PH and Aluminum Toxicity on the Photosynthetic Characteristics of Different Fast-Growing Eucalyptus Vegetatively Propagated Clones. PLoS ONE 2015, 10, e0130963. [Google Scholar] [CrossRef]
- Lyčka, M.; Barták, M.; Helia, O.; Kopriva, S.; Moravcová, D.; Hájek, J.; Fojt, L.; Čmelík, R.; Fajkus, J.; Fojtová, M. Sulfate supplementation affects nutrient and photosynthetic status of Arabidopsis thaliana and Nicotiana tabacum differently under prolonged exposure to cadmium. J. Hazard. Mater. 2023, 445, 130527. [Google Scholar] [CrossRef]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2022, 9, 2030082. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Duan, R.X.; Hu, W.J.; Zhang, N.N.; Lin, X.Y.; Zhang, J.H.; Zheng, H.L. Unravelling Calcium-Alleviated Aluminium Toxicity in Arabidopsis Thaliana: Insights into Regulatory Mechanisms Using Proteomics. J. Proteom. 2019, 199, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Mariano, E.D.; Keltjens, W.G. Long-Term Effects of Aluminum Exposure on Nutrient Uptake by Maize Genotypes Differing in Aluminum Resistance. J. Plant Nutr. 2005, 28, 323–333. [Google Scholar] [CrossRef]
- Danyaei, A.; Hassanpour, S.; Baghaee, M.A.; Dabbagh, M.; Babarabie, M. The Effect of Sulfur-Containing Humic Acid on Yield and Nutrient Uptake in Olive Fruit. Open J. Ecol. 2017, 7, 279–288. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Castellarín, J.M.; Miralles, D.J.; Pedrol, H.M. Sulfur Fertilization Improves Nitrogen Use Efficiency in Wheat by Increasing Nitrogen Uptake. Field Crops Res. 2009, 113, 170–177. [Google Scholar] [CrossRef]
- Shafiq, B.A.; Nawaz, F.; Majeed, S.; Aurangzaib, M.; Al Mamun, A.; Ahsan, M.; Ahmad, K.S.; Shehzad, M.A.; Ali, M.; Hashim, S.; et al. Sulfate-Based Fertilizers Regulate Nutrient Uptake, Potosynthetic Gas Exchange, and Enzymatic Antioxidants to Increase Sunflower Growth and Yield Under Drought Stress. J. Soil Sci. Plant Nutr. 2021, 21, 2229–2241. [Google Scholar] [CrossRef]
- Mariño Macana, Y.A.; de Toledo, F.H.S.F.; de Vicente Ferraz, A.; de Moraes Gonçalves, J.L.; Díaz López, S.M.; Barrero Cubillos, C.A.; Sierra Rojas, Á.N.; Bolognani, H.A. Soil Fertility and Fine Root Distribution after Gypsum Application in Eucalyptus Plantations with Different Tolerance to Water Deficit. New For. 2020, 51, 1039–1054. [Google Scholar] [CrossRef]
- Hussain, B.; Ma, Y.; Li, J.; Gao, J.; Ullah, A.; Tahir, N. Cadmium in Rice Is Affected by Fertilizer-Borne Chloride and Sulfate Anions: Long-Term Field Versus Pot Experiments. Processes 2022, 10, 1253. [Google Scholar] [CrossRef]
- Yan, S.; Yang, J.; Si, Y.; Tang, X.; Ma, Y.; Ye, W. Arsenic and cadmium bioavailability to rice (Oryza sativa L.) plant in paddy soil: Influence of sulfate application. Chemosphere 2022, 307, 135641. [Google Scholar] [CrossRef]
- Nagayama, T.; Nakamura, A.; Yamaji, N.; Satoh, S.; Furukawa, J.; Iwai, H. Changes in the Distribution of Pectin in Root Border Cells Under Aluminum Stress. Front. Plant Sci. 2019, 10, 1216. [Google Scholar] [CrossRef]
- Yan, L.; Riaz, M.; Liu, J.; Liu, Y.; Zeng, Y.; Jiang, C. Boron Reduces Aluminum Deposition in Alkali-Soluble Pectin and Cytoplasm to Release Aluminum Toxicity. J. Hazard. Mater. 2021, 401, 123388. [Google Scholar] [CrossRef]
- Jaskowiak, J.; Kwasniewska, J.; Milewska-Hendel, A.; Kurczynska, E.U.; Szurman-Zubrzycka, M.; Szarejko, I. Aluminum Alters the Histology and Pectin Cell Wall Composition of Barley Roots. Int. J. Mol. Sci. 2019, 20, 3039. [Google Scholar] [CrossRef]
- Sato, M.; Nagano, M.; J In, S.; Miyagi, A.; Yamaguchi, M.; Kawai-Yamada, M.; Ishikawa, T. Plant-Unique Cis/Trans Isomerism of Long-Chain Base Unsaturation Is Selectively Required for Aluminum Tolerance Resulting from Glucosylceramide-Dependent Plasma Membrane Fluidity. Plants 2020, 9, 19. [Google Scholar] [CrossRef]
- Kochian, L.V. Cellular Mechanisms of Aluminum Toxicity and Resistance in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 237–260. [Google Scholar] [CrossRef]
- Tanoi, K.; Kobayashi, N.I. Leaf Senescence by Magnesium Deficiency. Plants 2015, 4, 756–772. [Google Scholar] [CrossRef]
- Filipovic, M.R.; Jovanović, V.M. More than Just an Intermediate: Hydrogen Sulfide Signalling in Plants. J. Exp. Bot. 2017, 68, 4733–4736. [Google Scholar] [CrossRef]
- Silva, G.S.; Gavassi, M.A.; Nogueira, M.A.; Habermann, G. Aluminum Prevents Stomatal Conductance from Responding to Vapor Pressure Deficit in Citrus limonia. Environ. Exp. Bot. 2018, 155, 662–671. [Google Scholar] [CrossRef]
- Kasajima, I. Difference in Oxidative Stress Tolerance between Rice Cultivars Estimated with Chlorophyll Fluorescence Analysis. BMC Res. Notes 2017, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Maina, J.N.; Wang, Q. Seasonal Response of Chlorophyll a/b Ratio to Stress in a Typical Desert Species: Haloxylon Ammodendron. Arid. Land Res. Manag. 2015, 29, 321–334. [Google Scholar] [CrossRef]
- Mathot, M.; Thélier-Huché, L.; Lambert, R. Sulphur and Nitrogen Content as Sulphur Deficiency Indicator for Grasses. Eur. J. Agron. 2009, 30, 172–176. [Google Scholar] [CrossRef]
- Howarth, J.R.; Parmar, S.; Jones, J.; Shepherd, C.E.; Corol, D.I.; Galster, A.M.; Hawkins, N.D.; Miller, S.J.; Baker, J.M.; Verrier, P.J.; et al. Co-Ordinated Expression of Amino Acid Metabolism in Response to N and S Deficiency during Wheat Grain Filling. J. Exp. Bot. 2008, 59, 3675–3689. [Google Scholar] [CrossRef]
- Alam, A.; Tariq, M.; Ali, J.; Adnan, M.; Fahad, S.; Ahmad, M.; Romman, M.; Saleem, M.H.; Okla, M.K.; Ahmad, S.; et al. Co-application of Phosphorus and Sulphur Improve Yield, Quality, and Nutrients Uptake in Nicotiana tabaccum L. Philipp. Agric. Sci. 2022, 105, 61–68. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative Stress, Antioxidants and Stress Tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive Oxygen Gene Network of Plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Lima-Melo, Y.; Alencar, V.T.C.B.; Lobo, A.K.M.; Sousa, R.H.V.; Tikkanen, M.; Aro, E.M.; Silveira, J.A.G.; Gollan, P.J. Photoinhibition of Photosystem i Provides Oxidative Protection during Imbalanced Photosynthetic Electron Transport in Arabidopsis Thaliana. Front. Plant Sci. 2019, 10, 916. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic Stress and Reactive Oxygen Species: Generation, Signaling, and Defense Mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- Moustaka, J.; Ouzounidou, G.; Sperdouli, I.; Moustakas, M. Photosystem II Is More Sensitive than Photosystem I to Al3+ Induced Phytotoxicity. Materials 2018, 11, 1772. [Google Scholar] [CrossRef]
- Zhu, D.B.; Hu, K.D.; Guo, X.K.; Liu, Y.; Hu, L.Y.; Li, Y.H.; Wang, S.H.; Zhang, H. Sulfur Dioxide Enhances Endogenous Hydrogen Sulfide Accumulation and Alleviates Oxidative Stress Induced by Aluminum Stress in Germinating Wheat Seeds. Oxidative Med. Cell. Longev. 2015, 2015, 612363. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Type | Sequence (5′->3′) | Tm (°C) | Product Size (pb) |
|---|---|---|---|---|
| 26s proteosome subunit (26S) | Forward | TGCTTAGTTCCCCTAAGATAGTGA | 54 | 142 |
| Reverse | CTGAGACCAAACACGATTTCA | |||
| RuBisCO (RuBisCO) | Forward | TCATATCGAGCCTGTTGCTG | 60 | 144 |
| Reverse | AGAGCACGTAGGGCTTTGAA | |||
| Chlorophyll A binding protein (ChlAbp) | Forward | GCGCTCGGCCTATAATCTAC | 60 | 170 |
| Reverse | AACACACCACCCGAAAGAAG | |||
| Ferredoxin (Fered) | Forward | CGTCATCGAGACCCACAAGG | 58 | 93 |
| Reverse | GGTGGCGATGCCCATGTA | |||
| Copper–zinc superoxide dismutase (Cu/Zn-SOD) | Forward | CTCTTGCTAAGCTCATGTCC | 57 | 49 |
| Reverse | TCCTGGACTTCATGGCTTC | |||
| Iron superoxide dismu-tase (Fe-SOD) | Forward | GAAGCATCAGCAGGATCAC | 58 | 70 |
| Reverse | GTTAACTACCGAGAGTTTCCTC | |||
| Ascorbate peroxidase (APX) | Forward | GCTCTATCTGGTGGCCACAG | 57 | 82 |
| Reverse | TCAGAGGATCACGGGTCCAT | |||
| Glutathione reductase (GR) | Forward | TATCCGTCGTGGGTCTCAGT | 57 | 84 |
| Reverse | GACAGACTTCTCCTGCCGTT |
| Sulfate Nutrition (µM) | RuBisCO | ChlAbp | Fered |
|---|---|---|---|
| 120 | 7.54 * ± 2.20 | 11.49 * ± 3.96 | 2.24 * ± 0.40 |
| 240 | 1.84 ± 1.09 | 1.17 ± 0.55 | 1.12 ± 0.15 |
| 360 | 1.54 ± 0.24 | 1.30 ± 0.20 | 0.26 * ± 0.02 |
| Sulfate Nutrition (µM) | Fe-SOD | Cu/Zn SOD | APX | GR |
|---|---|---|---|---|
| 120 | 1.69 * ± 0.26 | 0.86 ± 0.15 | 1.63 * ± 0.34 | 0.93 ± 0.16 |
| 240 | 1.27 ± 0.13 | 1.11 ± 0.11 | 0.98 ± 0.03 | 1.01 ± 0.03 |
| 360 | 0.47 * ± 0.12 | 0.60 ± 0.12 | 0.47 * ± 0.07 | 0.80 ± 0.07 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vera-Villalobos, H.; Lunario-Delgado, L.; Gálvez, A.S.; Román-Silva, D.; Mercado-Seguel, A.; Wulff-Zottele, C. Sulfate Nutrition Modulates the Oxidative Response against Short-Term Al3+-Toxicity Stress in Lolium perenne cv. Jumbo Shoot Tissues. Agriculture 2024, 14, 1506. https://doi.org/10.3390/agriculture14091506
Vera-Villalobos H, Lunario-Delgado L, Gálvez AS, Román-Silva D, Mercado-Seguel A, Wulff-Zottele C. Sulfate Nutrition Modulates the Oxidative Response against Short-Term Al3+-Toxicity Stress in Lolium perenne cv. Jumbo Shoot Tissues. Agriculture. 2024; 14(9):1506. https://doi.org/10.3390/agriculture14091506
Chicago/Turabian StyleVera-Villalobos, Hernan, Lizzeth Lunario-Delgado, Anita S. Gálvez, Domingo Román-Silva, Ana Mercado-Seguel, and Cristián Wulff-Zottele. 2024. "Sulfate Nutrition Modulates the Oxidative Response against Short-Term Al3+-Toxicity Stress in Lolium perenne cv. Jumbo Shoot Tissues" Agriculture 14, no. 9: 1506. https://doi.org/10.3390/agriculture14091506