
Identification of Ascorbate Oxidase Genes and Their Response to Cold Stress in Citrus sinensis

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Genome-Wide Identification of the AAO Gene Family in Citrus sinensis
2.2. Physicochemical Properties Analysis
2.3. Gene Structure and Conserved Motif Analysis
2.4. Phylogenetic Tree, Ka/Ks, and Collinearity Analysis
2.5. Prediction of Cis-Acting Elements and microRNAs Targeting CsAAO Genes
2.6. Expression Profiles of CsLACs in Various Tissues
2.7. Plant Materials and Low Temperature Treatments
2.8. RNA Extraction and Expressing Profile Analysis
3. Results
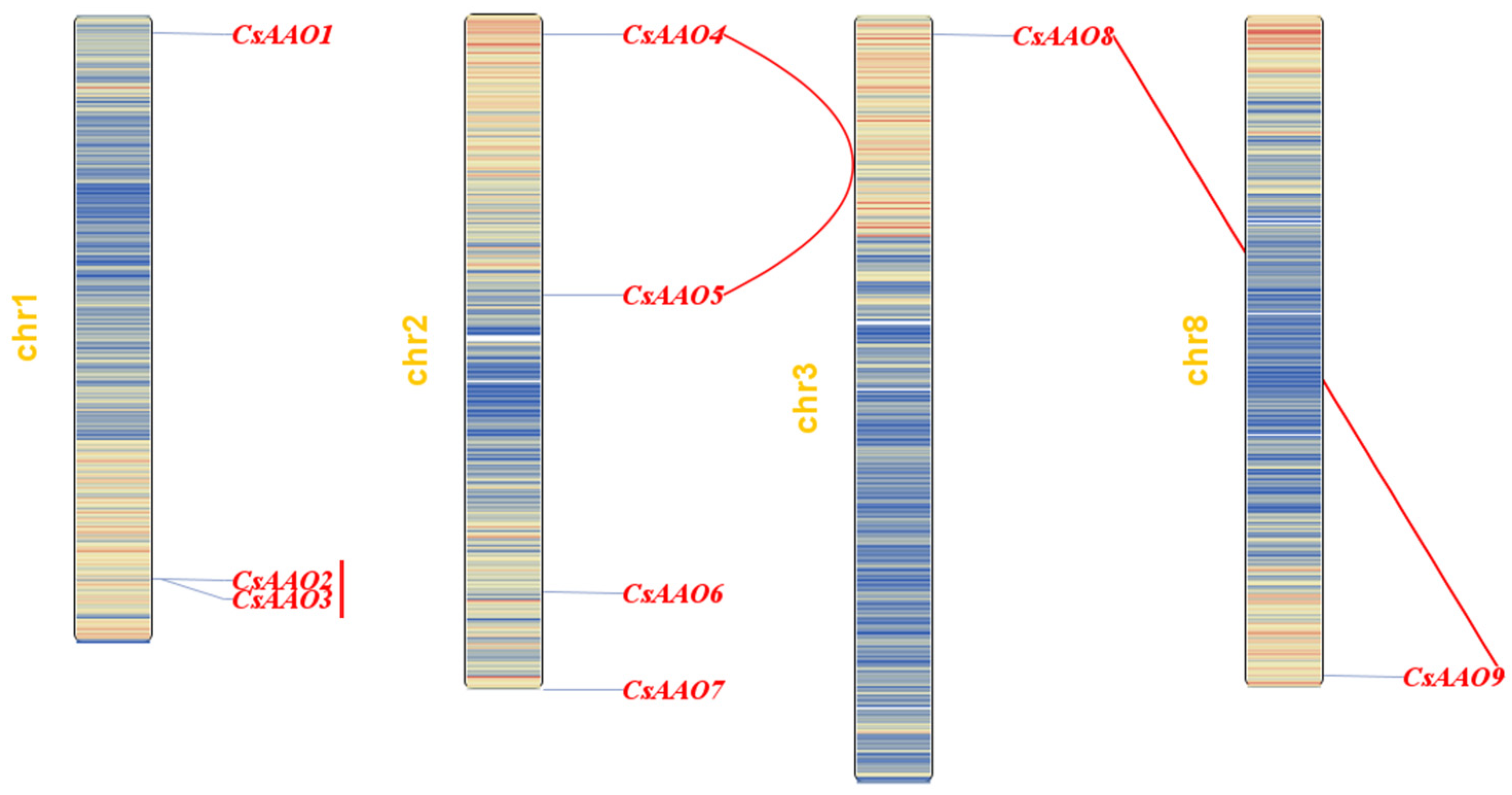
3.1. Identification, Physicochemical Properties, and Chromosomal Location of the CsAAO Family
3.2. Analysis of Conserved Motifs, Domain Composition, and Gene Structure of CsAAOs
3.3. Phylogenetic Analysis of AAO Proteins in Different Species
3.4. Gene Duplication and Genome Synteny Analysis of AAO Family Genes
3.5. Cis-Acting Element and MicroRNA Target Site Analysis of CsAAOs
3.6. Gene Expression Analysis of CsAAOs in Different Tissues
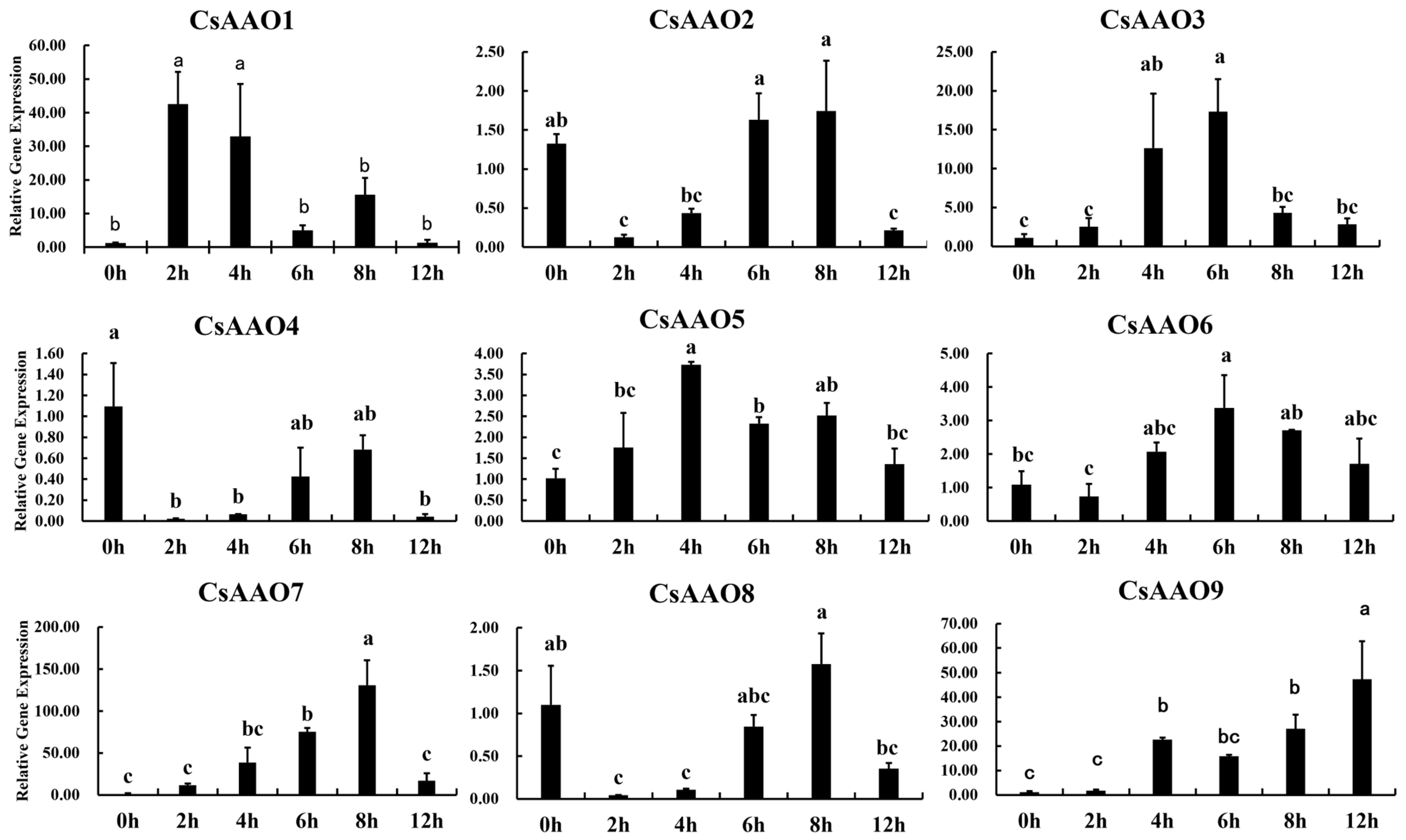
3.7. Analysis of CsAAOs Expression Patterns and Enzymatic Activities under Cold Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Stevens, R.; Truffault, V.; Baldet, P.; Gautier, H. Ascorbate oxidase in plant growth, development, and stress tolerance. In Ascorbic Acid in Plant Growth, Development and Stress Tolerance; Hossain, M.A., Munné-Bosch, S., Burritt, D.J., Diaz-Vivancos, P., Fujita, M., Lorence, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 273–295. [Google Scholar]
- Smirnoff, N. Ascorbic acid: Metabolism and functions of a multi-facetted molecule. Curr. Opin. Plant Biol. 2000, 3, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Mellidou, I.; Kanellis, A.K. Revisiting the role of ascorbate oxidase in plant systems. J. Exp. Bot. 2024, 75, 2740–2753. [Google Scholar] [CrossRef]
- Pignocchi, C.; Fletcher, J.M.; Wilkinson, J.E.; Barnes, J.D.; Foyer, C.H. The function of ascorbate oxidase in tobacco. Plant Physiol. 2003, 132, 1631–1641. [Google Scholar] [CrossRef] [PubMed]
- Sanmartin, M.; Drogoudi, P.D.; Lyons, T.; Pateraki, I.; Barnes, J.; Kanellis, A.K. Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta 2003, 216, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Chatzopoulou, F.; Sanmartin, M.; Mellidou, I.; Pateraki, I.; Koukounaras, A.; Tanou, G.; Kalamaki, M.S.; Veljovic-Jovanovic, S.; Antic, T.C.; Kostas, S.; et al. Silencing of ascorbate oxidase results in reduced growth, altered ascorbic acid levels and ripening pattern in melon fruit. Plant Physiol. Biochem. 2020, 156, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.X.; Liu, M.X.; Zhang, A.; Dai, Y.; Chen, W.Z.; Chen, F.; Wang, W.Y.; Shen, D.Y.; Telebanco-Yanoria, M.J.; Ren, B.; et al. Co-evolved plant and blast fungus ascorbate oxidases orchestrate the redox state of host apoplast to modulate rice immunity. Mol. Plant 2022, 15, 1347–1366. [Google Scholar] [CrossRef]
- Szent-Györgyi, A. On the function of hexuronic acid in the respiration of the cabbage leaf. J. Biol. Chem. 1931, 90, 385–393. [Google Scholar] [CrossRef]
- Ohkawa, J.; Okada, N.; Shinmyo, A.; Takano, M. Primary structure of cucumber (Cucumis sativus) ascorbate oxidase deduced from cDNA sequence: Homology with blue copper proteins and tissue-specific expression. Proc. Natl. Acad. Sci. USA 1989, 86, 1239–1243. [Google Scholar] [CrossRef]
- Esaka, M.; Hattori, T.; Fujisawa, K.; Sakajo, S.; Asahi, T. Molecular cloning and nucleotide sequence of full-length cDNA for ascorbate oxidase from cultured pumpkin cells. Eur. J. Biochem. 1990, 191, 537–541. [Google Scholar] [CrossRef]
- Kato, N.; Esaka, M. cDNA cloning and gene expression of ascorbate oxidase in tobacco. Plant Mol. Biol. 1996, 30, 833–837. [Google Scholar] [CrossRef]
- Diallinas, G.; Pateraki, I.; Sanmartin, M.; Scossa, A.; Stilianou, E.; Panopoulos, N.J.; Kanellis, A.K. Melon ascorbate oxidase: Cloning of a multigene family, induction during fruit development and repression by wounding. Plant Mol. Biol. 1997, 34, 759–770. [Google Scholar] [CrossRef] [PubMed]
- Batth, R.; Singh, K.; Kumari, S.; Mustafiz, A. Transcript profiling reveals the presence of abiotic stress and developmental stage specific ascorbate oxidase genes in plants. Front. Plant Sci. 2017, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Chen, L.H.; Wang, F.; Song, W.Y.; Cao, A.P.; Xie, S.Q.; Chen, X.F.; Jin, X.; Li, H.B. Genome-wide identification and expression analysis of the ascorbate oxidase gene family in Gossypium hirsutum reveals the critical role of GhAO1A in delaying dark-Induced leaf senescence. Int. J. Mol. Sci. 2019, 20, 6167. [Google Scholar] [CrossRef] [PubMed]
- Skorupa, M.; Szczepanek, J.; Yolcu, S.; Mazur, J.; Tretyn, A.; Tyburski, J. Characteristic of the ascorbate oxidase gene family in Beta vulgaris and analysis of the role of AAO in response to salinity and drought in Beet. Int. J. Mol. Sci. 2022, 23, 12773. [Google Scholar] [CrossRef]
- Zhu, M.; Liu, Q.; Liu, F.; Zheng, L.M.; Bing, J.; Zhou, Y.J.; Gao, F. Gene profiling of the ascorbate oxidase family genes under osmotic and cold stress reveals the role of AnAO5 in cold adaptation in Ammopiptanthus nanus. Plants 2023, 12, 677. [Google Scholar] [CrossRef]
- Sanmartin, M.; Pateraki, I.; Chatzopoulou, F.; Kanellis, A.K. Differential expression of the ascorbate oxidase multigene family during fruit development and in response to stress. Planta 2007, 225, 873–885. [Google Scholar] [CrossRef]
- Madhu; Kaur, A.; Singh, K.; Upadhyay, S.K. Ascorbate oxidases in bread wheat: Gene regulatory network, transcripts profiling, and interaction analyses provide insight into their role in plant development and stress response. Plant Growth Regul. 2024, 103, 209–224. [Google Scholar] [CrossRef]
- Abdelgawad, K.F.; El-Mogy, M.M.; Mohamed, M.I.A.; Garchery, C.; Stevens, R.G. Increasing ascorbic acid content and salinity tolerance of cherry tomato plants by suppressed expression of the ascorbate oxidase gene. Agronomy 2019, 9, 51. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Li, H.X.; Shu, W.B.; Zhang, C.J.; Zhang, W.; Ye, Z.B. Suppressed expression of ascorbate oxidase gene promotes ascorbic acid accumulation in tomato fruit. Plant Mol. Biol. Rep. 2011, 29, 638–645. [Google Scholar] [CrossRef]
- Fotopoulos, V.; Sanmartin, M.; Kanellis, A.K. Effect of ascorbate oxidase over-expression on ascorbate recycling gene expression in response to agents imposing oxidative stress. J. Exp. Bot. 2006, 57, 3933–3943. [Google Scholar] [CrossRef]
- Ahmed, R.; Kaldis, A.; Voloudakis, A. Silencing of a Nicotiana benthamiana ascorbate oxidase gene reveals its involvement in resistance against cucumber mosaic virus. Planta 2024, 259, 38. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.G.; Yang, R.X.; Yang, Z.R.; Yao, S.Z.; Zhao, S.S.; Wang, Y.; Li, P.C.; Song, X.W.; Jin, L.; Zhou, T.; et al. ROS accumulation and antiviral defence control by microRNA528 in rice. Nat. Plants 2017, 3, 16203. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; Pérez-Clemente, R.M.; Gómez-Cadenas, A.; López-Climent, M.F. Involvement of citrus shoots in response and tolerance to abiotic stress. Hortic. Adv. 2024, 2, 3. [Google Scholar] [CrossRef]
- Khan, M.; Dahro, B.; Wang, Y.; Wang, M.; Xiao, W.; Qu, J.; Zeng, Y.K.; Fang, T.; Xiao, P.; Xu, X.Y.; et al. The transcription factor ERF110 promotes cold tolerance by directly regulating sugar and sterol biosynthesis in citrus. Plant J. 2024, 119, 2385–2401. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Qu, J.; Wang, Y.; Fang, T.; Xiao, W.; Wang, Y.L.; Zhang, Y.; Khan, M.; Chen, Q.Y.; Xu, X.Y.; et al. Transcriptome and metabolome atlas reveals contributions of sphingosine and chlorogenic acid to cold tolerance in Citrus. Plant Physiol. 2024, 196, 634–650. [Google Scholar] [CrossRef]
- Xu, X.X.; Zhou, Y.P.; Wang, B.; Ding, L.; Wang, Y.; Luo, L.; Zhang, Y.L.; Kong, W.W. Genome-wide identification and characterization of laccase gene family in Citrus sinensis. Gene 2019, 689, 114–123. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zhang, Y.W.; Luo, L.; Lu, C.Y.; Kong, W.W.; Cheng, L.B.; Xu, X.Y.; Liu, J.H. Genome wide Identification of Respiratory burst oxidase homolog (Rboh) genes in Citrus sinensis and functional analysis of CsRbohD in cold tolerance. Int. J. Mol. Sci. 2022, 23, 648. [Google Scholar] [CrossRef]
- Xu, X.Y.; Zhang, Y.L.; Liang, M.G.; Kong, W.W.; Liu, J.H. The Citrus laccase gene CsLAC18 contributes to cold tolerance. Int. J. Mol. Sci. 2022, 23, 14509. [Google Scholar] [CrossRef]
- Chen, C.J.; Wu, Y.; Li, J.W.; Wang, X.; Zeng, Z.H.; Xu, J.; Liu, Y.L.; Feng, J.T.; Chen, H.; He, Y.H.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Yu, C.S.; Chen, Y.C.; Lu, C.H.; Hwang, J.K. Prediction of protein subcellular localization. Proteins Struct. Funct. Bioinf. 2006, 64, 643–651. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.B.; Zhuang, Z.H.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server (2017 release). Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-regulatory elements used to control gene expression in plants. Plant Cell Tissue Organ. Cult. 2016, 127, 269–287. [Google Scholar] [CrossRef]
- Li, S.J.; Castillo-Gonzalez, C.; Yu, B.; Zhang, X.R. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef]
- Tang, H.B.; Bowers, J.E.; Wang, X.Y.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Mohanty, B. Genomic architecture of promoters and transcriptional regulation of candidate genes in rice involved in tolerance to anaerobic germination. Curr. Plant Biol. 2022, 29, 100236. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Amino Acids Length | Molecular Weight (kDa) | Isoelectric Point (pI) | Instability Index (II) | Grand Average of Hydropathicity (GRAVY) | Subcellular Localization Predicted |
|---|---|---|---|---|---|---|---|
| CsAAO1 | Cs_ont_1g000870.1 | 586 | 66.16 | 8.78 | 40.14 | −0.266 | Extracell |
| CsAAO2 | Cs_ont_1g025500.1 | 539 | 60.29 | 9.17 | 34.78 | −0.199 | Extracell |
| CsAAO3 | Cs_ont_1g025510.1 | 541 | 60.48 | 9.15 | 39.82 | −0.164 | Extracell |
| CsAAO4 | Cs_ont_2g001690.1 | 543 | 61.47 | 9.73 | 43.86 | −0.259 | Extracell |
| CsAAO5 | Cs_ont_2g018990.1 | 538 | 59.52 | 8.09 | 38.34 | −0.227 | Extracell |
| CsAAO6 | Cs_ont_2g030090.1 | 580 | 64.72 | 7.37 | 32.56 | −0.318 | Extracell |
| CsAAO7 | Cs_ont_2g035780.1 | 554 | 62.19 | 8.27 | 32.50 | −0.252 | Extracell |
| CsAAO8 | Cs_ont_3g001440.1 | 542 | 60.21 | 9.27 | 30.76 | −0.227 | Extracell |
| CsAAO9 | Cs_ont_8g028600.1 | 545 | 60.91 | 9.32 | 37.26 | −0.234 | Extracell |
| Paralogous Genes | Ka | Ks | Ka/Ks | Selective Pressure |
|---|---|---|---|---|
| CsAAO2—CsAAO3 | 0.1689 | 1.6646 | 0.1015 | Purifying selection |
| CsAAO4—CsAAO5 | 0.1464 | 1.3449 | 0.1089 | Purifying selection |
| CsAAO8—CsAAO9 | 0.2440 | 1.4973 | 0.1629 | Purifying selection |
| Gene Name | Predicted miRNA Target Sites | miRNA Length | Expectation | Inhibition |
|---|---|---|---|---|
| CsAAO9 | csi-miR156h | 21 | 3.5 | Cleavage |
| CsAAO9 | csi-miR156i | 21 | 3.5 | Cleavage |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.; Miao, X.; Deng, N.; Liang, M.; Wang, L.; Jiang, L.; Zeng, S. Identification of Ascorbate Oxidase Genes and Their Response to Cold Stress in Citrus sinensis. Agriculture 2024, 14, 1643. https://doi.org/10.3390/agriculture14091643
Xu X, Miao X, Deng N, Liang M, Wang L, Jiang L, Zeng S. Identification of Ascorbate Oxidase Genes and Their Response to Cold Stress in Citrus sinensis. Agriculture. 2024; 14(9):1643. https://doi.org/10.3390/agriculture14091643
Chicago/Turabian StyleXu, Xiaoyong, Xingchen Miao, Naiyi Deng, Mengge Liang, Lun Wang, Lijuan Jiang, and Shaohua Zeng. 2024. "Identification of Ascorbate Oxidase Genes and Their Response to Cold Stress in Citrus sinensis" Agriculture 14, no. 9: 1643. https://doi.org/10.3390/agriculture14091643
APA StyleXu, X., Miao, X., Deng, N., Liang, M., Wang, L., Jiang, L., & Zeng, S. (2024). Identification of Ascorbate Oxidase Genes and Their Response to Cold Stress in Citrus sinensis. Agriculture, 14(9), 1643. https://doi.org/10.3390/agriculture14091643