
Environmental Factors Related to Climate Change Alter the Chemical Composition and Biological Activity of Lavandula viridis L’Hér Essential Oil

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Environmental Tests
2.2. Essential Oils
2.2.1. Extraction of EOs
2.2.2. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
2.2.3. Antioxidant Capacity
2.2.4. Enzymes’ (Tyrosinase and Cholinesterases) Inhibitory Capacity
2.3. Statistical Analysis
3. Results and Discussion
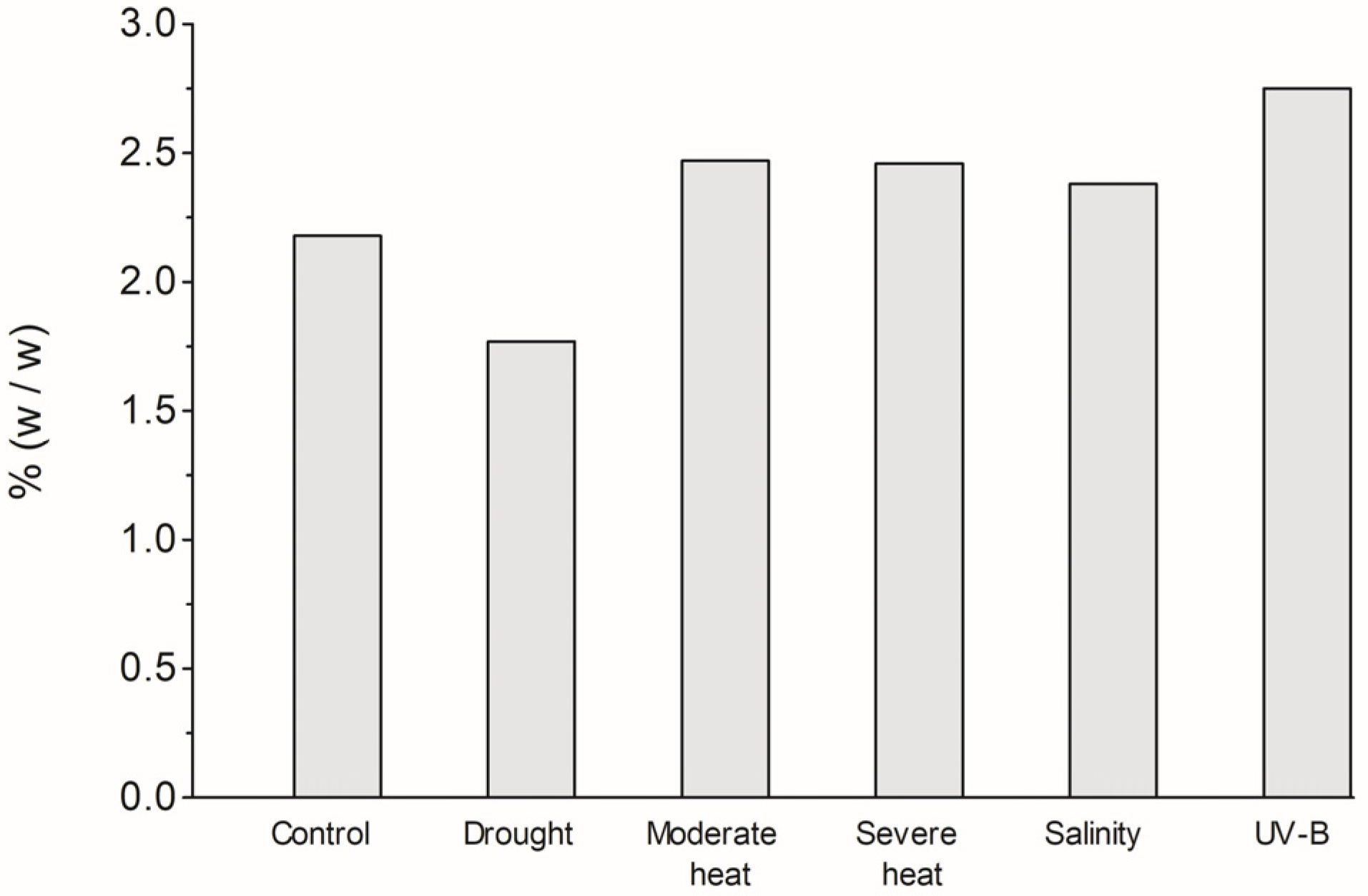
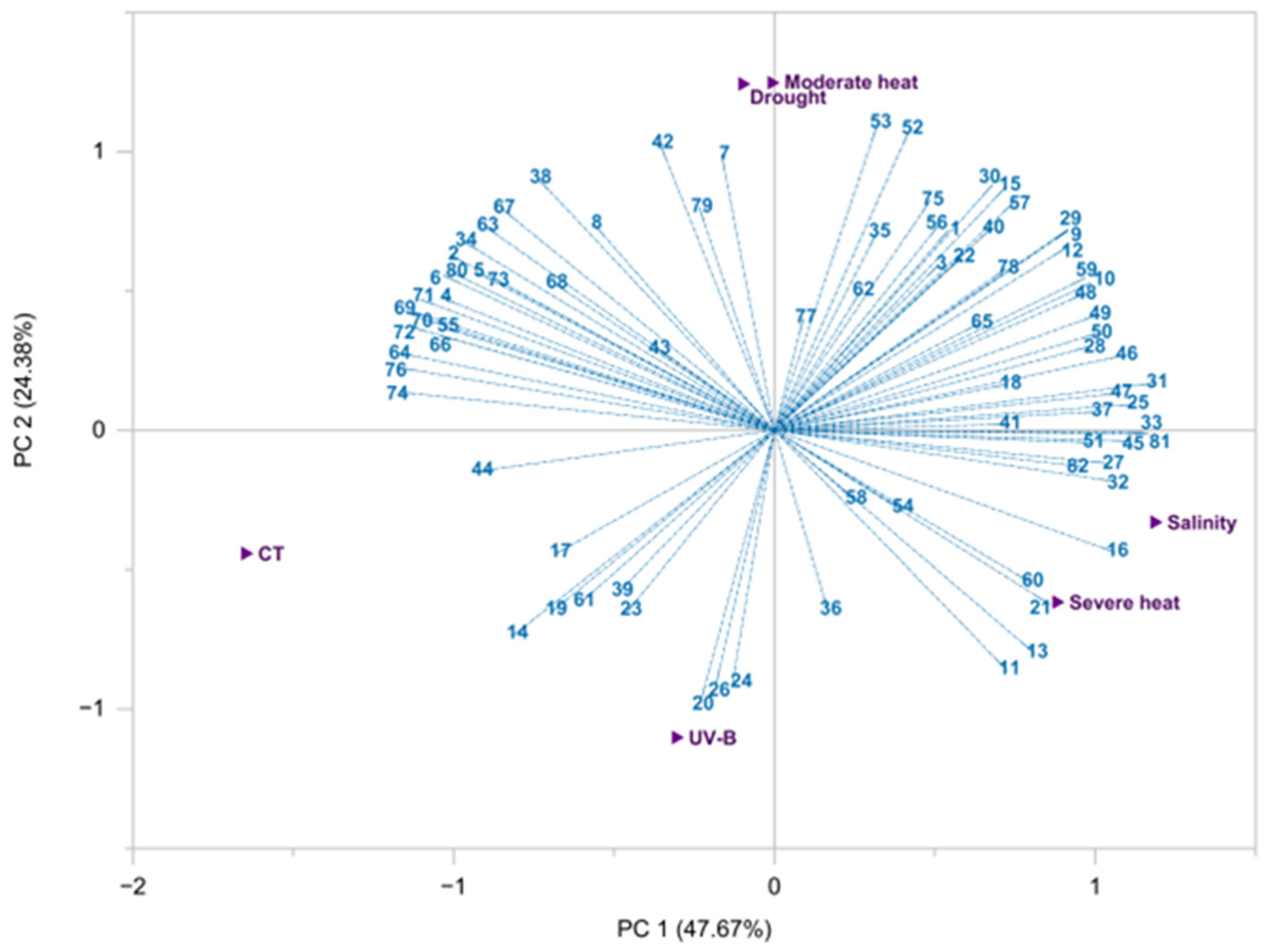
3.1. EO Extraction Yield and Chemical Profile
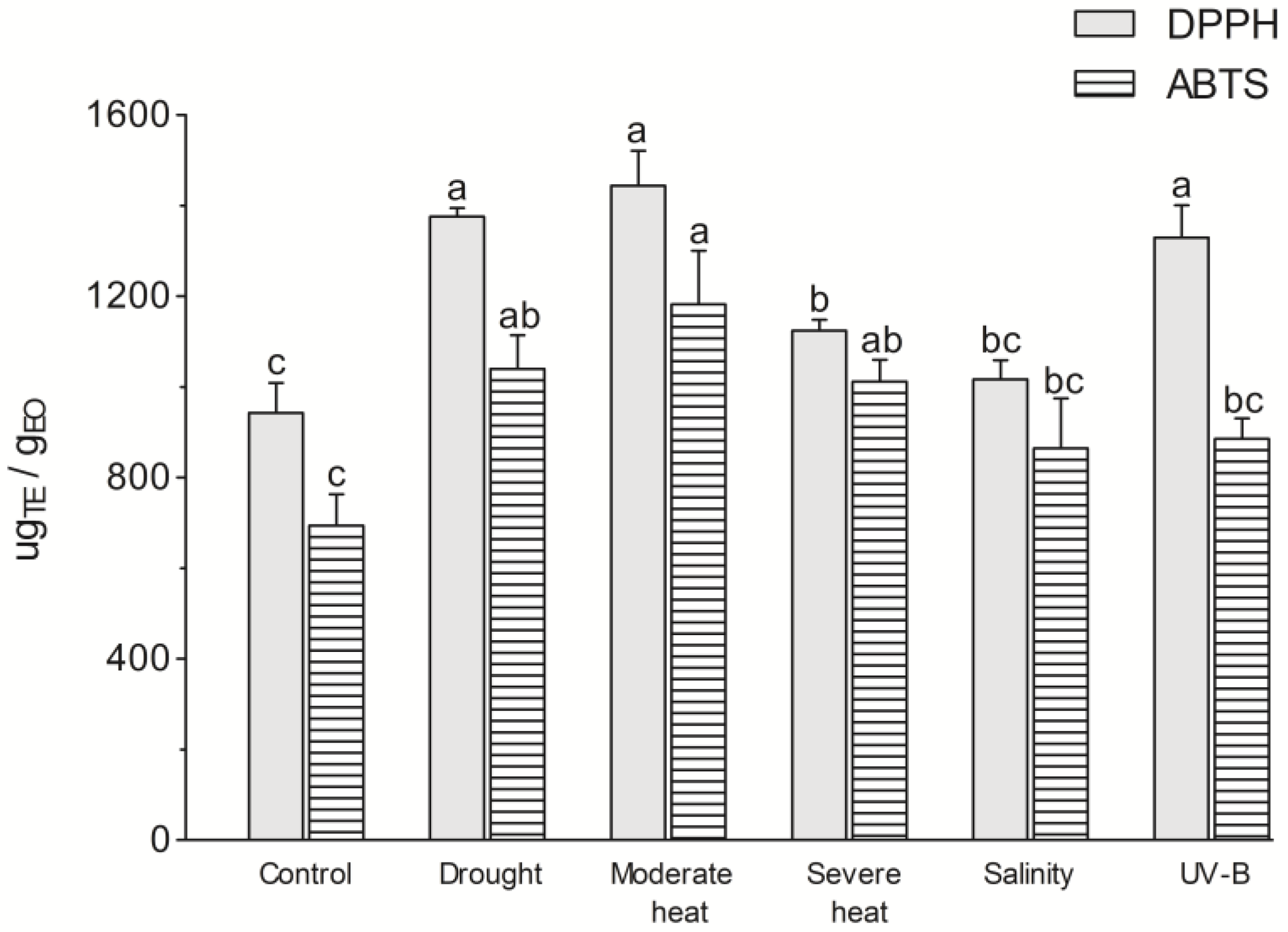
3.2. Antioxidant Potential of EOs
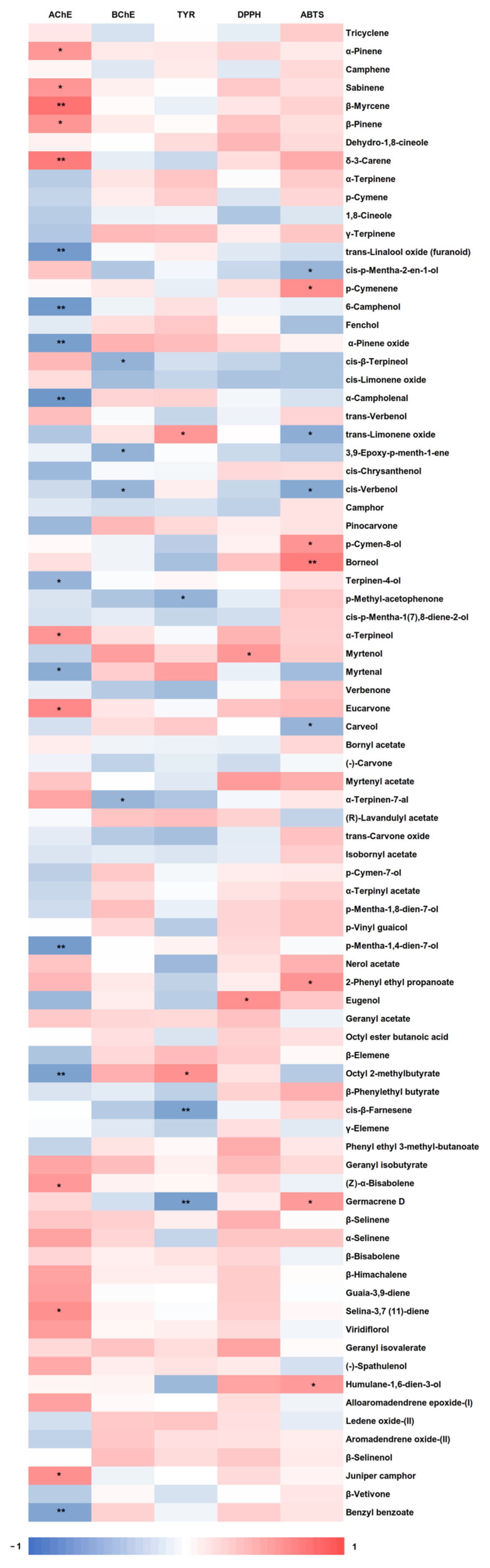
3.3. Enzyme Inhibitory Assays
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ballesteros, G.I.; Newsham, K.K.; Acuña-Rodríguez, I.S.; Atala, C.; Torres-Díaz, C.; Molina-Montenegro, M.A. Extreme Environments as Sources of Fungal Endophytes Mitigating Climate Change Impacts on Crops in Mediterranean-Type Ecosystems. Plants People Planet 2023, 6, 148–161. [Google Scholar] [CrossRef]
- Mansinhos, I.; Gonçalves, S.; Romano, A. How Climate Change-Related Abiotic Factors Affect the Production of Industrial Valuable Compounds in Lamiaceae Plant Species: A Review. Front. Plant Sci. 2024, 15, 1370810. [Google Scholar] [CrossRef]
- Verma, N.; Shukla, S. Impact of Various Factors Responsible for Fluctuation in Plant Secondary Metabolites. J. Appl. Res. Med. Aromat. Plants 2015, 2, 105–113. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.-T.; Moutinho-Pereira, J.; Correia, C.M. Drought Stress Effects and Olive Tree Acclimation under a Changing Climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Barut, M.; Tansı, L.S.; Karaman, S. Essential oil Composition of Lavender (Lavandula angustifolia Mill.) at Various Plantation Ages and Growth Stages in the Mediterranean Region. Turk. J. Agric.-Food Sci. Technol. 2022, 10, 746–753. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Teibo, J.O.; Wasef, L.; Shaheen, H.M.; Akomolafe, A.P.; Teibo, T.K.A.; Al-kuraishy, H.M.; Al-Garbeeb, A.I.; Alexiou, A.; Papadakis, M. A Review of the Bioactive Components and Pharmacological Properties of Lavandula Species. Naunyn-Schmiedebergs Arch. Pharmacol. 2023, 396, 877–900. [Google Scholar] [CrossRef] [PubMed]
- Mansinhos, I.; Gonçalves, S.; Rodríguez-Solana, R.; Ordóñez-Díaz, J.L.; Moreno-Rojas, J.M.; Romano, A. Impact of Temperature on Phenolic and Osmolyte Contents in In Vitro Cultures and Micropropagated Plants of Two Mediterranean Plant Species, Lavandula viridis and Thymus lotocephalus. Plants 2022, 11, 3516. [Google Scholar] [CrossRef] [PubMed]
- Bussotti, F.; Pollastrini, M. Opportunities and Threats of Mediterranean Evergreen Sclerophyllous Woody Species Subjected to Extreme Drought Events. Appl. Sci. 2020, 10, 8458. [Google Scholar] [CrossRef]
- Costa, P.; Sarmento, B.; Gonçalves, S.; Romano, A. Protective Effects of Lavandula viridis L’Hér Extracts and Rosmarinic Acid against H2O2-Induced Oxidative Damage in A172 Human Astrocyte Cell Line. Ind. Crops Prod. 2013, 50, 361–365. [Google Scholar] [CrossRef]
- Costa, P.; Gonçalves, S.; Valentão, P.; Andrade, P.B.; Romano, A. Accumulation of Phenolic Compounds in in Vitro Cultures and Wild Plants of Lavandula viridis L’Hér and Their Antioxidant and Anti-Cholinesterase Potential. Food Chem. Toxicol. 2013, 57, 69–74. [Google Scholar] [CrossRef]
- Costa, P.; Grosso, C.; Gonçalves, S.; Andrade, P.B.; Valentão, P.; Gabriela Bernardo-Gil, M.; Romano, A. Supercritical Fluid Extraction and Hydrodistillation for the Recovery of Bioactive Compounds from Lavandula viridis L’Hér. Food Chem. 2012, 135, 112–121. [Google Scholar] [CrossRef]
- Gonçalves, S.; Mansinhos, I.; Rodríguez-Solana, R.; Pereira-Caro, G.; Moreno-Rojas, J.M.; Romano, A. Impact of Metallic Nanoparticles on In Vitro Culture, Phenolic Profile and Biological Activity of Two Mediterranean Lamiaceae Species: Lavandula viridis L’Hér and Thymus lotocephalus G. López and R. Morales. Molecules 2021, 26, 6427. [Google Scholar] [CrossRef] [PubMed]
- Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Canhoto, J.; Vale-Silva, L.; Silva, M.J.; Pinto, E.; Salgueiro, L. Chemical Composition and Antifungal Activity of the Essential oils of Lavandula viridis L’Hér. J. Med. Microbiol. 2011, 60, 612–618. [Google Scholar] [CrossRef]
- Costa, P.; Grevenstuk, T.; Rosa da Costa, A.M.; Gonçalves, S.; Romano, A. Antioxidant and Anti-Cholinesterase Activities of Lavandula viridis L’Hér Extracts after in Vitro Gastrointestinal Digestion. Ind. Crops Prod. 2014, 55, 83–89. [Google Scholar] [CrossRef]
- Costa, P.; Medronho, B.; Gonçalves, S.; Romano, A. Cyclodextrins Enhance the Antioxidant Activity of Essential oils from Three Lamiaceae Species. Ind. Crops Prod. 2015, 70, 341–346. [Google Scholar] [CrossRef]
- Costa, S.; Cavadas, C.; Cavaleiro, C.; Salgueiro, L.; do Céu Sousa, M. In Vitro Susceptibility of Trypanosoma Brucei Brucei to Selected Essential oils and Their Major Components. Exp. Parasitol. 2018, 190, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, P.; Lima, A.S.; Vieira, P.; Dias, L.S.; Tinoco, M.T.; Barroso, J.G.; Pedro, L.G.; Figueiredo, A.C.; Mota, M. Nematicidal Activity of Essential oils and Volatiles Derived from Portuguese Aromatic Flora against the Pinewood Nematode, Bursaphelenchus Xylophilus. J. Nematol. 2010, 42, 8–16. [Google Scholar]
- Machado, M.; Martins, N.; Salgueiro, L.; Cavaleiro, C.; Sousa, M.C. Lavandula luisieri and Lavandula viridis Essential oils as Upcoming Anti-Protozoal Agents: A Key Focus on Leishmaniasis. Appl. Sci. 2019, 9, 3056. [Google Scholar] [CrossRef]
- Mateus, D.M.R.; Ferraz, E.; Perna, V.; Sales, P.; Hipólito-Correia, V. Essential oils and Extracts of Plants as Biocides against Microorganisms Isolated from the Ruins of the Roman City of Conímbriga (Portugal). Environ. Sci. Pollut. Res. 2024, 31, 40669–40677. [Google Scholar] [CrossRef]
- Zuzarte, M.; Francisco, V.; Neves, B.; Liberal, J.; Cavaleiro, C.; Canhoto, J.; Salgueiro, L.; Cruz, M.T. Lavandula viridis L’Hér Essential oil Inhibits the Inflammatory Response in Macrophages through Blockade of NF-KB Signaling Cascade. Front. Pharmacol. 2022, 12, 695911. [Google Scholar] [CrossRef]
- ISO 3515:2002; Oil of Lavender (Lavandula angustifolia Mill). ISO: Geneva, Switzerland, 2002.
- ISO 8902:2009; Oil of Lavandin Grosso (Lavandula angustifolia Mill. x Lavandula Latifolia Medik.), French Type. ISO: Geneva, Switzerland, 2009.
- Green Lavender Essential Oil. Available online: https://otefarm.eu/product/green-lavender-essential-oil/ (accessed on 21 June 2024).
- Green Lavender Essential Oil, 19.50 €. Available online: https://www.fontepenedo.com/en/essential-oils/lavandula-viridis-oil (accessed on 21 June 2024).
- Óleo Essencial de Rosmaninho Verde (Branco) SELVAGEM (Lavandula viridis). Available online: https://info714569.wixsite.com/btiquantum/product-page/óleo-essencial-de-lavandula-viridis-selvagem (accessed on 21 June 2024).
- Óleo Essencial Lavanda-Branca GT Portugal 5 mL. Available online: https://www.laszlo.com.br/oleo-essencial-lavanda-laszlo-lavanda-branca-gt-portugal-sku-7799-laszlo.html (accessed on 21 June 2024).
- Aceite Esencial de Lavanda Verde Silvestre y BIO. Available online: https://www.micosmeticacasera.es/aceite-esencial-de-lavanda-verde-bio/ (accessed on 21 June 2024).
- Nogueira, J.M.F.; Romano, A. Essential oils from Micropropagated Plants of Lavandula viridis. Phytochem. Anal. 2002, 13, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.-M.; Peng, J.-Q.; Chen, Y.; Tao, L.; Zhang, Y.-Y.; Fu, L.-Y.; Long, Q.-D.; Shen, X.-C. 1,8-Cineole: A Review of Source, Biological Activities, and Application. J. Asian Nat. Prod. Res. 2021, 23, 938–954. [Google Scholar] [CrossRef]
- Gonçalves, S.; Serra, H.; Nogueira, J.M.F.; Almeida, R.; Custódio, L.; Romano, A. Headspace-SPME of in Vitro Shoot-Cultures and Micropropagated Plants of Lavandula viridis. Biol. Plant. 2008, 52, 133–136. [Google Scholar] [CrossRef]
- Matos, F.; Miguel, M.G.; Duarte, J.; Venâncio, F.; Moiteiro, C.; Correia, A.I.D.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Antioxidant Capacity of the Essential oils from Lavandula luisieri, L. stoechas subsp. lusitanica, L. stoechas subsp. lusitanica x L. luisieri and L. viridis Grown in Algarve (Portugal). J. Essent. Oil Res. 2009, 21, 327–336. [Google Scholar] [CrossRef]
- Mansinhos, I.; Gonçalves, S.; Rodríguez-Solana, R.; Ordóñez-Díaz, J.L.; Moreno-Rojas, J.M.; Romano, A. Ultrasonic-Assisted Extraction and Natural Deep Eutectic Solvents Combination: A Green Strategy to Improve the Recovery of Phenolic Compounds from Lavandula pedunculata subsp. lusitanica (Chaytor) Franco. Antioxidants 2021, 10, 582. [Google Scholar] [CrossRef] [PubMed]
- Manukyan, A. Effects of PAR and UV-B Radiation on Herbal Yield, Bioactive Compounds and Their Antioxidant Capacity of Some Medicinal Plants Under Controlled Environmental Conditions. Photochem. Photobiol. 2013, 89, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Agrawal, S.B. Comparative Analysis of Essential oil Composition and Oil Containing Glands in Ocimum sanctum L. (Holy Basil) under Ambient and Supplemental Level of UV-B through Gas Chromatography–Mass Spectrometry and Scanning Electron Microscopy. Acta Physiol. Plant. 2011, 33, 1093–1101. [Google Scholar] [CrossRef]
- Manukyan, A. Secondary Metabolites and Their Antioxidant Capacity of Caucasian Endemic Thyme (Thymus Transcaucasicus Ronn.) as Affected by Environmental Stress. J. Appl. Res. Med. Aromat. Plants 2019, 13, 100209. [Google Scholar] [CrossRef]
- Akbari, G.A.; Soltani, E.; Binesh, S.; Amini, F. Cold Tolerance, Productivity and Phytochemical Diversity in Sweet Basil (Ocimum basilicum L.) Accessions. Ind. Crops Prod. 2018, 124, 677–684. [Google Scholar] [CrossRef]
- Tarchoune, I.; Baâtour, O.; Harrathi, J.; Cioni, P.L.; Lachaâl, M.; Flamini, G.; Ouerghi, Z. Essential oil and Volatile Emissions of Basil (Ocimum basilicum) Leaves Exposed to NaCl or Na2SO4 Salinity. J. Plant Nutr. Soil Sci. 2013, 176, 748–755. [Google Scholar] [CrossRef]
- Baatour, O.; Kaddour, R.; Aidi Wannes, W.; Lachaâl, M.; Marzouk, B. Salt Effects on the Growth, Mineral Nutrition, Essential oil Yield and Composition of Marjoram (Origanum majorana). Acta Physiol. Plant. 2010, 32, 45–51. [Google Scholar] [CrossRef]
- Bidgoli, D.R.; Azarnezhad, N.; Akhbari, M.; Ghorbani, M. Salinity Stress and PGPR Effects on Essential oil Changes in Rosmarinus officinalis L. Agric. Food Secur. 2019, 8, 2. [Google Scholar] [CrossRef]
- Said-Al Ahl, H.A.H.; Sabra, A.S.; Alataway, A.; Astatkie, T.; Mahmoud, A.A.; Bloem, E. Biomass Production and Essential oil Composition of Thymus Vulgaris in Response to Water Stress and Harvest Time. J. Essent. Oil Res. 2019, 31, 63–68. [Google Scholar] [CrossRef]
- Aprotosoaie, A.C.; Gille, E.; Trifan, A.; Luca, V.S.; Miron, A. Essential oils of Lavandula Genus: A Systematic Review of Their Chemistry. Phytochem. Rev. 2017, 16, 761–799. [Google Scholar] [CrossRef]
- Domingues, J.; Delgado, F.; Gonçalves, J.C.; Zuzarte, M.; Duarte, A.P. Mediterranean Lavenders from Section Stoechas: An Undervalued Source of Secondary Metabolites with Pharmacological Potential. Metabolites 2023, 13, 337. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Li, Q.; Niu, Y.; Zhou, X.; Liu, J.; Xu, Y.; Xu, Z. Odor-Active Compounds of Different Lavender Essential oils and Their Correlation with Sensory Attributes. Ind. Crops Prod. 2017, 108, 748–755. [Google Scholar] [CrossRef]
- Aćimović, M.; Lončar, B.; Pezo, M.; Stanković Jeremić, J.; Cvetković, M.; Rat, M.; Pezo, L. Volatile Compounds of Nepeta nuda L. from Rtanj Mountain (Serbia). Horticulturae 2022, 8, 85. [Google Scholar] [CrossRef]
- Aziz, E.E.; Al-Amier, H.; Craker, L.E. Influence of Salt Stress on Growth and Essential oil Production in Peppermint, Pennyroyal, and Apple Mint. J. Herbs Spices Med. Plants 2008, 14, 77–87. [Google Scholar] [CrossRef]
- Bahcesular, B.; Yildirim, E.D.; Karaçocuk, M.; Kulak, M.; Karaman, S. Seed Priming with Melatonin Effects on Growth, Essential oil Compounds and Antioxidant Activity of Basil (Ocimum basilicum L.) under Salinity Stress. Ind. Crops Prod. 2020, 146, 112165. [Google Scholar] [CrossRef]
- Elhindi, K.M.; Al-Suhaibani, N.A.; El-Din, A.F.S.; Yakout, S.M.; Al-Amri, S.M. Effect of Foliar-Applied Iron and Zinc on Growth Rate and Essential oil in Sweet Basil (Ocimum basilicum L.) under Saline Conditions. Progr. Nutr. 2016, 18, 288–298. [Google Scholar]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic Acid-Regulated Antioxidant Mechanisms and Gene Expression Enhance Rosemary Performance under Saline Conditions. Front. Physiol. 2017, 8, 716. [Google Scholar] [CrossRef]
- Tounekti, T.; Vadel, A.M.; Bedoui, A.; Khemira, H. NaCl Stress Affects Growth and Essential oil Composition in Rosemary (Rosmarinus officinalis L.). J. Hortic. Sci. Biotechnol. 2008, 83, 267–273. [Google Scholar] [CrossRef]
- Es-sbihi, F.Z.; Hazzoumi, Z.; Aasfar, A.; Amrani Joutei, K. Improving Salinity Tolerance in Salvia officinalis L. by Foliar Application of Salicylic Acid. Chem. Biol. Technol. Agric. 2021, 8, 25. [Google Scholar] [CrossRef]
- Masoudniaragh, A.; Oraei, M.; Gohari, G.; Akbari, A.; Faramarzi, A. Using Halloysite Nanotubes as Carrier for Proline to Alleviate Salt Stress Effects in Sweet Basil (Ocimum basilicum L.). Sci. Hortic. 2021, 285, 110202. [Google Scholar] [CrossRef]
- Taarit, B.M.; Msaada, K.; Hosni, K.; Hammami, M.; Kchouk, M.E.; Marzouk, B. Plant Growth, Essential oil Yield and Composition of Sage (Salvia officinalis L.) Fruits Cultivated under Salt Stress Conditions. Ind. Crops Prod. 2009, 30, 333–337. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V.; Niazi, A.; Moghadam, A. Effect of Salt Stress on Terpenoid Biosynthesis in Salvia Mirzayanii: From Gene to Metabolite. J. Hortic. Sci. Biotechnol. 2019, 94, 389–399. [Google Scholar] [CrossRef]
- Valifard, M.; Mohsenzadeh, S.; Kholdebarin, B.; Rowshan, V. Effects of Salt Stress on Volatile Compounds, Total Phenolic Content and Antioxidant Activities of Salvia Mirzayanii. S. Afr. J. Bot. 2014, 93, 92–97. [Google Scholar] [CrossRef]
- Kulak, M. Recurrent Drought Stress Effects on Essential oil Profile of Lamiaceae Plants: An Approach Regarding Stress Memory. Ind. Crops Prod. 2020, 154, 112695. [Google Scholar] [CrossRef]
- Ramezani, S.; Abbasi, A.; Sobhanverdi, S.; Shojaeiyan, A.; Ahmadi, N. The Effects of Water Deficit on the Expression of Monoterpene Synthases and Essential oils Composition in Salvia Ecotypes. Physiol. Mol. Biol. Plants 2020, 26, 2199–2207. [Google Scholar] [CrossRef] [PubMed]
- Chrysargyris, A.; Laoutari, S.; Litskas, V.D.; Stavrinides, M.C.; Tzortzakis, N. Effects of Water Stress on Lavender and Sage Biomass Production, Essential oil Composition and Biocidal Properties against Tetranychus Urticae (Koch). Sci. Hortic. 2016, 213, 96–103. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought Effects on Polyphenol Composition and Antioxidant Activities in Aerial Parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Llorens-Molina, J.A.; Vacas, S. Effect of Drought Stress on Essential oil Composition of Thymus vulgaris L. (Chemotype 1, 8-Cineole) from Wild Populations of Eastern Iberian Peninsula. J. Essent. Oil Res. 2017, 29, 145–155. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Samani, M.R.; Hashemi, M.; Zeinali, H. Salicylic Acid Affects Growth, Essential oil and Chemical Compositions of Thyme (Thymus Daenensis Celak.) under Reduced Irrigation. Plant Growth Regul. 2014, 72, 289–301. [Google Scholar] [CrossRef]
- Radwan, A.; Kleinwächter, M.; Selmar, D. Impact of Drought Stress on Specialised Metabolism: Biosynthesis and the Expression of Monoterpene Synthases in Sage (Salvia officinalis). Phytochemistry 2017, 141, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Kulak, M.; Gul, F.; Sekeroglu, N. Changes in Growth Parameter and Essential oil Composition of Sage (Salvia officinalis L.) Leaves in Response to Various Salt Stresses. Ind. Crops Prod. 2020, 145, 112078. [Google Scholar] [CrossRef]
- Chang, X.; Alderson, P.G.; Wright, C.J. Enhanced UV-B Radiation Alters Basil (Ocimum basilicum L.) Growth and Stimulates the Synthesis of Volatile oils. J. Hortic. For. 2009, 1, 27–31. Available online: https://academicjournals.org/journal/JHF/article-stat/58E7FAC4938 (accessed on 26 June 2024).
- El Kharraf, S.; Faleiro, M.L.; Abdellah, F.; El-Guendouz, S.; El Hadrami, E.M.; Miguel, M.G. Simultaneous Hydrodistillation-Steam Distillation of Rosmarinus Officinalis, Lavandula angustifolia and Citrus Aurantium from Morocco, Major Terpenes: Impact on Biological Activities. Molecules 2021, 26, 5452. [Google Scholar] [CrossRef]
- Eltayeb, L.M.H.; Yagi, S.; Mohamed, H.M.M.; Zengin, G.; Shariati, M.A.; Rebezov, M.; Uba, A.I.; Lorenzo, J.M. Essential oils Composition and Biological Activity of Chamaecyparis Obtusa, Chrysopogon Nigritanus and Lavandula Coronopifolia Grown Wild in Sudan. Molecules 2023, 28, 1005. [Google Scholar] [CrossRef]
- Carrasco, A.; Tomas, V.; Tudela, J.; Miguel, M.G. Comparative Study of GC-MS Characterization, Antioxidant Activity and Hyaluronidase Inhibition of Different Species of Lavandula and Thymus Essential oils. Flavour Fragr. J. 2016, 31, 57–69. [Google Scholar] [CrossRef]
- Gok, M.; Cicek, C.; Sari, S.; Bodur, E. Novel Activity of Human BChE: Lipid Hydrolysis. Biochimie 2023, 204, 127–135. [Google Scholar] [CrossRef]
- Pham, T.-N.; Cazier, E.A.; Gormally, E.; Lawrence, P. Valorization of Biomass Polyphenols as Potential Tyrosinase Inhibitors. Drug Discov. Today 2024, 29, 103843. [Google Scholar] [CrossRef] [PubMed]
- Vittorio, S.; Dank, C.; Ielo, L. Heterocyclic Compounds as Synthetic Tyrosinase Inhibitors: Recent Advances. Int. J. Mol. Sci. 2023, 24, 9097. [Google Scholar] [CrossRef]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Subhan, F.; Ahmed, J. Neuroprotective and Anti-Aging Potentials of Essential oils from Aromatic and Medicinal Plants. Front. Aging Neurosci. 2017, 9, 168. [Google Scholar] [CrossRef]
- Gonçalves, S.; Mansinhos, I.; Romano, A. Neuroprotective Compounds from Plant Sources and Their Modes of Action: An Update. In Plant-Derived Bioactives: Chemistry and Mode of Action; Swamy, M.K., Ed.; Springer: Singapore, 2020; pp. 417–440. ISBN 9789811523618. [Google Scholar]
- Chen, W.N.; Chin, K.W.; Tang, K.S.; Agatonovic-Kustrin, S.; Yeong, K.Y. Neuroprotective, Neurite Enhancing, and Cholinesterase Inhibitory Effects of Lamiaceae Family Essential oils in Alzheimer’s Disease Model. J. Herb. Med. 2023, 41, 100696. [Google Scholar] [CrossRef]
- Kharraf, S.E.; El-Guendouz, S.; Farah, A.; Mateus, M.C.; Hadrami, E.M.E.; Miguel, M.G. Impact of Fifteen Combinations of the Main Components of Rosemary, Lavender and Citrus Essential oils on in Vitro Biological Activities. S. Afr. J. Bot. 2023, 156, 162–168. [Google Scholar] [CrossRef]
- El Omari, N.; Balahbib, A.; Bakrim, S.; Benali, T.; Ullah, R.; Alotaibi, A.; Naceiri El Mrabti, H.; Goh, B.H.; Ong, S.-K.; Ming, L.C.; et al. Fenchone and Camphor: Main Natural Compounds from Lavandula stoechas L., Expediting Multiple in Vitro Biological Activities. Heliyon 2023, 9, e21222. [Google Scholar] [CrossRef]
- Nwanade, C.F.; Wang, M.; Li, H.; Masoudi, A.; Yu, Z.; Liu, J. Individual and Synergistic Toxicity of Cinnamon Essential oil Constituents against Haemaphysalis Longicornis (Acari: Ixodidae) and Their Potential Effects on Non-Target Organisms. Ind. Crops Prod. 2022, 178, 114614. [Google Scholar] [CrossRef]
- Dragomanova, S.; Lazarova, M.; Munkuev, A.; Suslov, E.; Volcho, K.; Salakhutdinov, N.; Bibi, A.; Reynisson, J.; Tzvetanova, E.; Alexandrova, A.; et al. New Myrtenal-Adamantane Conjugates Alleviate Alzheimer’s-Type Dementia in Rat Model. Molecules 2022, 27, 5456. [Google Scholar] [CrossRef] [PubMed]
- Prabu, S.; Jing, D.; Chandran, V.; Mathew, P. Insecticidal Activity of Origanum majorana L. Essential oil as Anti-Cholinergic. Agent. Entomol. Res. 2020, 50, 402–413. [Google Scholar] [CrossRef]
- Tran-Trung, H.; Thang, T.D.; Nguyen, T.H.D.; Vu, D.C.; Tuan, N.H.; Ha, N.X.; Chen, T.V.; Oanh, H.T.; Giang, N.T.T.; Thuy, P.T. Essential oils From the Trunks and Leaves of Paramignya Scandens (Griff.) Craib From Vietnam: Phytochemical Composition, In Vitro α-Amylase and Tyrosinase Inhibitory Activities and In Silico Molecular Docking Studies. Nat. Prod. Commun. 2023, 18, 1934578X231222383. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Class | RI * | RI Literature | Composition (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | Drought | Moderate Heat | Severe Heat | Salinity | UV-B | |||||
| 1 | Tricyclene | MH | 938 | 938 | 0.41 ± 0.01 b | 0.47 ± 0.03 ab | 0.46 ± 0.01 ab | 0.49 ± 0.02 a | 0.44 ± 0.02 ab | 0.34 ± 0.01 c |
| 2 | α-Pinene | MH | 948 | 947 | 5.99 ± 0.11 a | 5.25 ± 0.21 b | 5.53 ± 0.09 ab | 3.57 ± 0.18 cd | 3.14 ± 0.17 d | 4.09 ± 0.30 c |
| 3 | Camphene | MH | 967 | 967 | 3.61 ± 0.09 a | 3.72 ± 0.16 a | 3.85 ± 0.04 a | 3.93 ± 0.21 a | 3.68 ± 0.23 a | 3.34 ± 0.26 a |
| 4 | Sabinene | MH | 986 | 985 | 0.81 ± 0.01 a | 0.67 ± 0.02 b | 0.77 ± 0.00 a | 0.53 ± 0.02 c | 0.41 ± 0.02 d | 0.61 ± 0.03 b |
| 5 | Myrcene | MH | 992 | 992 | 0.28 ± 0.00 a | 0.19 ± 0.01 c | 0.25 ± 0.00 b | 0.13 ± 0.00 d | 0.05 ± 0.00 f | 0.11 ± 0.00 |
| 6 | β-Pinene | MH | 995 | 995 | 2.08 ± 0.03 a | 1.76 ± 0.02 b | 1.96 ± 0.02 a | 1.20 ± 0.04 d | 0.95 ± 0.03 e | 1.48 ± 0.08 c |
| 7 | Dehydro-1,8-cineole | OM | 1003 | 1004 | 0.51 ± 0.00 bc | 0.68 ± 0.00 a | 0.55 ± 0.03 b | 0.47 ± 0.01 c | 0.49 ± 0.02 bc | 0.47 ± 0.01 c |
| 8 | δ-3-Carene | MH | 1024 | 1022 | 0.74 ± 0.01 b | 0.66 ± 0.01 c | 0.84 ± 0.01 a | 0.60 ± 0.02 d | 0.16 ± 0.01 f | 0.23 ± 0.01 e |
| 9 | α-Terpinene | MH | 1031 | 1030 | 0.44 ± 0.01 c | 0.58 ± 0.03 ab | 0.57 ± 0.01 abc | 0.53 ± 0.04 abc | 0.61 ± 0.06 a | 0.47 ± 0.05 bc |
| 10 | p-Cymene | MH | 1044 | 1042 | 0.72 ± 0.05 a | 0.89 ± 0.05 a | 0.87 ± 0.01 a | 0.87 ± 0.06 a | 0.97 ± 0.13 a | 0.72 ± 0.03 a |
| 11 | 1,8-Cineole | OM | 1050 | 1050 | 24.25 ± 0.03 c | 21.56 ± 0.26 d | 24.55 ± 0.01 c | 28.98 ± 0.88 a | 28.37 ± 0.74 ab | 26.88 ± 0.76 b |
| 12 | γ-Terpinene | MH | 1071 | 1071 | 0.61 ± 0.01 c | 0.71 ± 0.02 ab | 0.75 ± 0.01 a | 0.68 ± 0.02 abc | 0.76 ± 0.04 a | 0.65 ± 0.03 bc |
| 13 | trans-Linalool oxide | OM | 1084 | 1084 | 0.48 ± 0.00 b | 0.51 ± 0.03 b | 0.52 ± 0.01 b | 0.69 ± 0.04 a | 0.75 ± 0.05 a | 0.75 ± 0.05 a |
| 14 | cis-p Mentha-2-en-1-ol | OM | 1086 | 1085 | 0.75 ± 0.03 a | 0.42 ± 0.01 c | 0.28 ± 0.03 d | 0.54 ± 0.01 b | 0.34 ± 0.00 d | 0.53 ± 0.03 b |
| 15 | p-Cymenene | MH | 1093 | 1093 | 0.05 ± 0.00 c | 0.08 ± 0.00 b | 0.09 ± 0.00 a | 0.08 ± 0.00 b | 0.08 ± 0.00 b | 0.05 ± 0.00 c |
| 16 | 6-Camphenol | OM | 1100 | 1097 | 0.92 ± 0.01 b | 1.01 ± 0.02 ab | 0.98 ± 0.02 ab | 1.07 ± 0.04 a | 1.10 ± 0.05 a | 1.06 ± 0.05 a |
| 17 | Fenchol | OM | 1104 | 1105 | 2.94 ± 0.18 ab | 2.43 ± 0.20 b | 1.67 ± 0.15 c | 1.22 ± 0.05 c | 2.40 ± 0.19 b | 3.36 ± 0.15 a |
| 18 | α-Pinene oxide | OM | 1106 | 1106 | 0.20 ± 0.01 c | 0.31 ± 0.00 b | 0.29 ± 0.00 b | 0.24 ± 0.00 c | 0.35 ± 0.02 a | 0.32 ± 0.02 ab |
| 19 | cis-β-Terpineol | OM | 1118 | 1121 | 0.26 ± 0.00 a | 0.19 ± 0.00 d | 0.14 ± 0.00 f | 0.23 ± 0.01 b | 0.16 ± 0.00 e | 0.21 ± 0.00 c |
| 20 | cis-Limonene oxide | OM | 1134 | 1134 | 0.31 ± 0.01 a | 0.24 ± 0.01 bc | 0.21 ± 0.01 c | 0.32 ± 0.02 a | 0.26 ± 0.01 bc | 0.28 ± 0.01 ab |
| 21 | α-Campholenal | OM | 1146 | 1136 | 1.00 ± 0.04 b | 1.03 ± 0.04 b | 1.07 ± 0.02 ab | 1.12 ± 0.03 ab | 1.21 ± 0.05 a | 1.18 ± 0.05 a |
| 22 | trans-Verbenol | OM | 1153 | 1153 | 0.11 ± 0.01 a | 0.11 ± 0.01 a | 0.12 ± 0.00 a | 0.12 ± 0.00 a | 0.12 ± 0.01 a | 0.09 ± 0.02 a |
| 23 | trans-Limonene oxide | OM | 1155 | 1151 | 0.12 ± 0.02 a | 0.11 ± 0.01 a | 0.10 ± 0.00 a | 0.10 ± 0.00 a | 0.12 ± 0.01 a | 0.13 ± 0.01 a |
| 24 | 3,9-Epoxy-p-menth-1-ene | OM | 1159 | 1178 | 0.09 ± 0.02 a | 0.06 ± 0.00 a | 0.05 ± 0.00 a | 0.11 ± 0.03 a | 0.06 ± 0.01 a | 0.09 ± 0.03 a |
| 25 | cis-Chrysanthenol | OM | 1166 | 1168 | 1.17 ± 0.02 b | 1.40 ± 0.01 a | 1.36 ± 0.09 a | 1.47 ± 0.01 a | 1.43 ± 0.08 a | 1.36 ± 0.02 a |
| 26 | cis-Verbenol | OM | 1170 | 1170 | 4.27 ± 0.10 a | 4.10 ± 0.11 a | 3.45 ± 0.03 b | 4.27 ± 0.09 a | 4.05 ± 0.08 a | 4.35 ± 0.13 a |
| 27 | Camphor | OM | 1176 | 1174 | 10.74 ± 0.10 d | 11.28 ± 0.16 c | 11.61 ± 0.06 c | 13.21 ± 0.03 a | 12.52 ± 0.12 b | 10.86 ± 0.10 d |
| 28 | Pinocarvone | OM | 1189 | 1181 | 3.41 ± 0.00 d | 4.24 ± 0.08 b | 4.21 ± 0.05 b | 3.96 ± 0.10 c | 4.74 ± 0.06 a | 4.06 ± 0.08 bc |
| 29 | p-Cymen-8-ol | OM | 1195 | 1194 | 0.17 ± 0.04 b | 0.31 ± 0.03 a | 0.29 ± 0.02 ab | 0.30 ± 0.04 a | 0.30 ± 0.03 ab | 0.18 ± 0.02 bc |
| 30 | Borneol | OM | 1199 | 1199 | 2.31 ± 0.05 c | 2.75 ± 0.19 a | 2.76 ± 0.01 a | 2.70 ± 0.12 ab | 2.55 ± 0.06 abc | 2.36 ± 0.05 bc |
| 31 | Terpinen-4-ol | OM | 1201 | 1200 | 1.29 ± 0.04 c | 1.52 ± 0.07 ab | 1.56 ± 0.03 ab | 1.63 ± 0.06 ab | 1.65 ± 0.01 a | 1.47 ± 0.03 b |
| 32 | p-Methyl-acetophenone | Others | 1207 | 1199 | 0.32 ± 0.04 c | 0.39 ± 0.02 cb | 0.38 ± 0.03 bc | 0.50 ± 0.01 a | 0.43 ± 0.03 ab | 0.38 ± 0.03 bc |
| 33 | cis-p-Mentha-1(7),8-diene-2-ol | OM | 1213 | 1206 | 0.26 ± 0.05 b | 0.32 ± 0.01 ab | 0.31 ± 0.00 ab | 0.38 ± 0.02 a | 0.38 ± 0.00 a | 0.29 ± 0.02 b |
| 34 | α-Terpineol | OM | 1215 | 1213 | 1.79 ± 0.01 a | 1.69 ± 0.05 a | 1.78 ± 0.02 a | 1.18 ± 0.04 c | 1.05 ± 0.03 c | 1.41 ± 0.05 b |
| 35 | Myrtenol | OM | 1220 | 1224 | 1.08 ± 0.02 b | 1.24 ± 0.03 a | 1.25 ± 0.02 a | 1.08 ± 0.02 b | 1.21 ± 0.04 a | 1.20 ± 0.00 a |
| 36 | Myrtenal | OM | 1223 | 1223 | 1.01 ± 0.03 ab | 1.02 ± 0.05 ab | 0.96 ± 0.01 b | 0.98 ± 0.03 ab | 1.09 ± 0.03 a | 1.09 ± 0.02 a |
| 37 | Verbenone | OM | 1236 | 1236 | 2.28 ± 0.02 c | 2.79 ± 0.07 ab | 2.58± 0.05 bc | 3.14 ± 0.20 a | 2.80 ± 0.13 ab | 2.48 ± 0.11 bc |
| 38 | Eucarvone | OM | 1244 | 1245 | 0.14 ± 0.01 a | 0.14 ± 0.00 a | 0.15 ± 0.00 a | 0.08 ± 0.01 b | 0.06 ± 0.00 c | 0.08 ± 0.01 bc |
| 39 | Carveol | OM | 1250 | 1252 | 0.31 ± 0.00 ab | 0.28 ± 0.01 v | 0.23 ± 0.00 c | 0.22 ± 0.02 c | 0.30 ± 0.01 ab | 0.34 ± 0.02 a |
| 40 | Bornyl acetate | Others | 1254 | 1255 | 0.22 ± 0.00 ab | 0.25 ± 0.01 a | 0.24 ± 0.00 ab | 0.25 ± 0.02 ab | 0.25 ± 0.02 ab | 0.20 ± 0.01 b |
| 41 | (-)-Carvone | OM | 1266 | 1265 | 0.54 ± 0.02 a | 0.60 ± 0.02 a | 0.52 ± 0.04 a | 0.58 ± 0.02 a | 0.60 ± 0.03 a | 0.53 ± 0.04 a |
| 42 | Myrtenyl acetate | Others | 1272 | 1284 | 0.24 ± 0.00 bc | 0.29 ± 0.01 a | 0.26 ± 0.00 ab | 0.22 ± 0.02 bc | 0.21 ± 0.01 c | 0.22 ± 0.01 bc |
| 43 | α-Terpinen-7-al | OM | 1277 | 1276 | 0.19 ± 0.01 ab | 0.20 ± 0.00 a | 0.15 ± 0.01 b | 0.19 ± 0.02 ab | 0.15 ± 0.01 b | 0.15 ± 0.01 b |
| 44 | (R)-Lavandulyl acetate | Others | 1281 | 1282 | 0.66 ± 0.00 a | 0.60 ± 0.02 b | 0.57 ± 0.01 bc | 0.45 ± 0.01 d | 0.54 ± 0.00 c | 0.69 ± 0.00 a |
| 45 | trans-Carvone oxide | OM | 1295 | 1280 | 0.14 ± 0.01 a | 0.18 ± 0.01 a | 0.16 ± 0.03 a | 0.20 ± 0.01 a | 0.19 ± 0.02 a | 0.16 ± 0.01 a |
| 46 | Isobornyl acetate | Others | 1304 | 1304 | 0.44 ± 0.00 d | 0.53 ± 0.00 bc | 0.51 ± 0.01 c | 0.58 ± 0.02 a | 0.57 ± 0.01 ab | 0.45 ± 0.01 d |
| 47 | p-Cymen-7-ol | OM | 1308 | 1308 | 0.21 ± 0.00 b | 0.26 ± 0.01 ab | 0.26 ± 0.01 ab | 0.26 ± 0.02 ab | 0.30 ± 0.02 a | 0.26 ± 0.03 ab |
| 48 | α-Terpinyl acetate | Others | 1313 | 1314 | 0.14 ± 0.00 b | 0.18 ± 0.01 a | 0.17 ± 0.00 ab | 0.17 ± 0.01 ab | 0.19 ± 0.01 a | 0.16 ± 0.01 ab |
| 49 | p-Mentha-1,8-dien-7-ol | OM | 1319 | 1313 | 0.12 ± 0.00 b | 0.14 ± 0.01 ab | 0.14 ± 0.00 ab | 0.13 ± 0.01 ab | 0.15 ± 0.00 a | 0.13 ± 0.01 ab |
| 50 | p-Vinyl guaicol | Others | 1328 | 1328 | 0.09 ± 0.00 a | 0.15 ± 0.05 a | 0.17 ± 0.00 a | 0.15 ± 0.06 a | 0.17 ± 0.06 a | 0.15 ± 0.07 a |
| 51 | p-Mentha-1,4-dien-7-ol | OM | 1345 | 1333 | 0.27 ± 0.01 c | 0.34 ± 0.00 ab | 0.32 ± 0.01 b | 0.33 ± 0.00 ab | 0.36 ± 0.02 a | 0.34 ± 0.00 ab |
| 52 | Nerol acetate | Others | 1359 | 1359 | 0.12 ± 0.03 a | 0.15 ± 0.02 a | 0.15 ± 0.02 a | 0.14 ± 0.06 a | 0.13 ± 0.05 a | 0.12 ± 0.03 a |
| 53 | 2-Phenyl ethyl propanoate | Others | 1363 | 1356 | 0.15 ± 0.01 a | 0.17 ± 0.01 a | 0.18 ± 0.01 a | 0.15 ± 0.02 a | 0.16 ± 0.01 a | 0.13 ± 0.01 a |
| 54 | Eugenol | OM | 1369 | 1369 | 0.36 ± 0.01 c | 0.41 ± 0.00 b | 0.43 ± 0.00 b | 0.43 ± 0.02 b | 0.41 ± 0.01 b | 0.49 ± 0.01 a |
| 55 | Geranyl acetate | Others | 1377 | 1377 | 1.51 ± 0.02 a | 1.46 ± 0.02 ab | 1.27 ± 0.01 c | 0.82 ± 0.05 e | 1.00 ± 0.03 d | 1.34 ± 0.05 bc |
| 56 | Octyl ester butanoic acid | Others | 1387 | 1386 | 0.10 ± 0.00 a | 0.14 ± 0.02 a | 0.13 ± 0.00 a | 0.10 ± 0.02 a | 0.14 ± 0.06 a | 0.12 ± 0.04 a |
| 57 | β-Elemene | SH | 1414 | 1416 | 0.08 ± 0.00 a | 0.13 ± 0.03 a | 0.12 ± 0.04 a | 0.11 ± 0.03 a | 0.11 ± 0.03 a | 0.10 ± 0.01 a |
| 58 | Octyl 2-methylbutyrate | Others | 1433 | 1436 | 0.32 ± 0.05 bc | 0.38 ± 0.02 ab | 0.33 ± 0.01 bc | 0.29 ± 0.02 c | 0.43 ± 0.01 a | 0.44 ± 0.01 a |
| 59 | β-Phenylethyl butyrate | Others | 1455 | 1457 | 0.29 ± 0.00 b | 0.37 ± 0.02 a | 0.34 ± 0.01 ab | 0.35 ± 0.00 ab | 0.36 ± 0.02 ab | 0.33 ± 0.03 ab |
| 60 | cis-β-Farnesene | SH | 1459 | 1459 | 0.10 ± 0.00 b | 0.10 ± 0.01 ab | 0.10 ± 0.00 b | 0.12 ± 0.01 a | 0.11 ± 0.00 ab | 0.10 ± 0.01 ab |
| 61 | γ-Elemene | SH | 1485 | 1482 | 0.13 ± 0.02 a | 0.13 ± 0.01 a | 0.10 ± 0.01 a | 0.12 ± 0.01 a | 0.11 ± 0.02 a | 0.16 ± 0.02 a |
| 62 | Phenyl ethyl 3-methyl-butanoate | Others | 1504 | 1508 | 1.50 ± 0.04 b | 2.14 ± 0.18 a | 1.72 ± 0.02 ab | 1.52 ± 0.09 b | 1.88 ± 0.16 ab | 1.88 ±0.14 ab |
| 63 | Geranyl isobutyrate | Others | 1509 | 1511 | 0.31 ± 0.01 ab | 0.30 ± 0.00 ab | 0.31 ± 0.00 a | 0.19 ± 0.03 d | 0.22 ± 0.02 cd | 0.26 ± 0.01 bc |
| 64 | (Z)-α-Bisabolene | SH | 1514 | 1509 | 0.77 ± 0.02 a | 0.57 ± 0.01 b | 0.51 ± 0.01 b | 0.26 ± 0.04 c | 0.22 ± 0.05 c | 0.48 ± 0.06 b |
| 65 | Germacrene D | SH | 1520 | 1519 | 0.15 ± 0.02 a | 0.17 ± 0.01 a | 0.18 ± 0.03 a | 0.18 ± 0.02 a | 0.16 ± 0.03 a | 0.16 ± 0.03 a |
| 66 | β-Selinene | SH | 1531 | 1531 | 1.38 ± 0.01 a | 1.34 ± 0.02 a | 1.28 ± 0.01 a | 0.99 ± 0.06 b | 1.06 ± 0.04 b | 1.32 ± 0.08 a |
| 67 | α-Selinene | SH | 1537 | 1534 | 0.51 ± 0.02 a | 0.51 ± 0.02 a | 0.53 ± 0.01 a | 0.45 ± 0.06 a | 0.45 ± 0.06 a | 0.48 ± 0.05 a |
| 68 | β-Bisabolene | SH | 1557 | 1547 | 0.23 ± 0.01 ab | 0.25 ± 0.01 a | 0.20 ± 0.00 bc | 0.17 ± 0.01 c | 0.20 ± 0.01 bc | 0.21 ± 0.02 bc |
| 69 | β-Himachalene | SH | 1563 | 1550 | 0.26 ± 0.00 a | 0.22 ± 0.00 b | 0.21 ± 0.00 b | 0.12 ± 0.00 d | 0.10 ± 0.01 e | 0.18 ± 0.00 c |
| 70 | Guaia-3,9-diene | SH | 1567 | 1556 | 0.80 ± 0.01 a | 0.70 ± 0.02 a | 0.54 ± 0.01 b | 0.30 ± 0.04 c | 0.22 ± 0.04 c | 0.51 ± 0.04 b |
| 71 | Selina-3,7 (11)-diene | SH | 1578 | 1562 | 0.59 ± 0.01 a | 0.50 ± 0.20 ab | 0.46 ± 0.01 bc | 0.26 ± 0.03 d | 0.22 ± 0.03 d | 0.38 ± 0.04 c |
| 72 | Viridiflorol | OS | 1585 | 1584 | 2.42 ± 0.03 a | 2.05 ± 0.05 b | 1.73 ± 0.01 c | 1.06 ± 0.05 d | 1.01 ± 0.02 d | 1.63 ± 0.08 c |
| 73 | Geranyl isovalerate | OS | 1601 | 1600 | 0.74 ± 0.01 a | 0.77 ± 0.00 a | 0.73 ± 0.01 a | 0.57 ± 0.03 b | 0.63 ± 0.04 b | 0.73 ± 0.01 a |
| 74 | (-)-Spathulenol | OS | 1604 | 1619 | 0.25 ± 0.03 a | 0.22 ± 0.01 ab | 0.20 ± 0.00 ab | 0.17 ± 0.01 b | 0.18 ± 0.00 b | 0.21 ±0.01 ab |
| 75 | Humulane-1,6-dien-3-ol | OS | 1623 | 1619 | 0.15 ± 0.00 a | 0.18 ± 0.01 a | 0.18 ± 0.00 a | 0.17 ± 0.01 a | 0.16 ± 0.02 a | 0.17 ± 0.02 a |
| 76 | Alloaromadendrene epoxide-(I) | OS | 1657 | 1646 | 0.94 ± 0.02 a | 0.77 ± 0.02 b | 0.70 ± 0.04 b | 0.49 ± 0.02 c | 0.47 ± 0.06 c | 0.71 ± 0.08 b |
| 77 | Ledene oxide-(II) | OS | 1660 | 1678 | 0.91 ± 0.00 ab | 1.11 ± 0.05 a | 0.93 ± 0.01 ab | 0.79 ± 0.04 b | 1.08 ± 0.12 a | 1.00 ± 0.03 a |
| 78 | Aromadendrene oxide-(II) | OS | 1682 | 1678 | 0.46 ± 0.02 c | 0.62 ± 0.00 a | 0.55 ± 0.02 ab | 0.50 ± 0.03 c | 0.63 ± 0.06 a | 0.52 ± 0.03 ab |
| 79 | β-selinenol | OS | 1705 | 1691 | 0.85 ± 0.00 ab | 0.96 ± 0.02 a | 0.89 ± 0.04 ab | 0.74 ± 0.05 b | 0.88 ± 0.07 ab | 0.85 ± 0.07 ab |
| 80 | Juniper camphor | OS | 1743 | 1741 | 0.80 ± 0.00 a | 0.75 ± 0.02 a | 0.59 ± 0.02 b | 0.39 ± 0.01 c | 0.28 ± 0.02 d | 0.45 ± 0.02 c |
| 81 | β-Vetivone | OS | 1795 | 1808 | 0.61 ± 0.01 b | 0.70 ± 0.00 ab | 0.70 ± 0.01 ab | 0.72 ± 0.02 ab | 0.77 ± 0.08 a | 0.70 ± 0.06 ab |
| 82 | Benzyl benzoate | Others | 1808 | 1789 | 0.72 ± 0.00 b | 0.88 ± 0.01 a | 0.92 ± 0.00 a | 0.91 ± 0.03 a | 1.01 ± 0.06 a | 0.99 ± 0.05 a |
| Grouped compounds | ||||||||||
| Monoterpene hydrocarbons (MH) | 15.73 ± 0.18 a | 14.98± 0.54 a | 15.93 ± 0.17 a | 12.61 ± 0.61 b | 11.24 ± 0.72 b | 12.08 ± 0.78 b | ||||
| Oxygenated monoterpenes (OM) | 64.02 ± 0.05 c | 63.90 ± 0.02 c | 64.99 ± 0.01 c | 71.93 ± 0.39 a | 72.05 ± 0.55 a | 69.00 ± 0.50 b | ||||
| Sesquiterpene hydrocarbons (SH) | 4.99 ± 0.06 a | 4.61 ± 0.12 ab | 4.22 ± 0.03 b | 3.08 ± 0.25 c | 2.96 ± 0.24 c | 4.08 ± 0.33 b | ||||
| Oxygenated sesquiterpenes (OS) | 8.12 ± 0.07 a | 8.11 ± 0.14 a | 7.20 ± 0.10 ab | 5.59 ± 0.28 d | 6.09 ± 0.49 cd | 6.97 ± 0.42 bc | ||||
| Others | 7.13 ± 0.10 ab | 8.40 ± 0.30 a | 7.65 ± 0.09 ab | 6.79 ± 0.46 b | 7.69 ± 0.52 ab | 7.86 ± 0.52 ab | ||||
| Treatment | AChE Inhibition (IC50, μg/mL) | BChE Inhibition (IC50, μg/mL) | Tyr Inhibition (IC50, μg/mL) |
|---|---|---|---|
| Control | 781.33 ± 31.70 a | 4479.75 ± 94.99 ab | 6303.13 ± 952.03 a |
| Drought | 515.56 ± 117.51 bc | 3919.30 ± 182.36 b | 5991.08 ± 665.98 ab |
| Moderate heat | 626.07 ± 8.53 ab | 6027.41 ± 825.47 a | 5478.96 ± 994.46 ab |
| Severe heat | 566.21 ± 96.06 abc | 2250.22 ± 205.42 c | 3551.01 ± 315.98 b |
| Salinity | 406.56 ± 53.43 bc | 5660.39 ± 505.35 a | 7332.91 ± 682.69 a |
| UV-B | 338.80 ± 9.11 c | 4799.53 ± 293.32 ab | 5612.41 ± 712.80 ab |
| Kojic acid | 6.88 ± 0.77 | ||
| Galantamine | 10.05 ± 0.03 | 153.13 ± 8.82 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansinhos, I.; Gonçalves, S.; Rodríguez-Solana, R.; Moreno-Rojas, J.M.; Romano, A. Environmental Factors Related to Climate Change Alter the Chemical Composition and Biological Activity of Lavandula viridis L’Hér Essential Oil. Agriculture 2024, 14, 1067. https://doi.org/10.3390/agriculture14071067
Mansinhos I, Gonçalves S, Rodríguez-Solana R, Moreno-Rojas JM, Romano A. Environmental Factors Related to Climate Change Alter the Chemical Composition and Biological Activity of Lavandula viridis L’Hér Essential Oil. Agriculture. 2024; 14(7):1067. https://doi.org/10.3390/agriculture14071067
Chicago/Turabian StyleMansinhos, Inês, Sandra Gonçalves, Raquel Rodríguez-Solana, José Manuel Moreno-Rojas, and Anabela Romano. 2024. "Environmental Factors Related to Climate Change Alter the Chemical Composition and Biological Activity of Lavandula viridis L’Hér Essential Oil" Agriculture 14, no. 7: 1067. https://doi.org/10.3390/agriculture14071067
APA StyleMansinhos, I., Gonçalves, S., Rodríguez-Solana, R., Moreno-Rojas, J. M., & Romano, A. (2024). Environmental Factors Related to Climate Change Alter the Chemical Composition and Biological Activity of Lavandula viridis L’Hér Essential Oil. Agriculture, 14(7), 1067. https://doi.org/10.3390/agriculture14071067