
Impact of Salinity Changes on the Antioxidation of Juvenile Yellowfin Tuna (Thunnus albacares)

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish and Design
2.2. Analytical Method
2.3. Statistical Analysis
3. Results
3.1. Change of Gill State of Juvenile Yellowfin Tuna under Salinity Stress
3.2. Changes in Liver Status of Juvenile Yellowfin Tuna under Salinity Stress
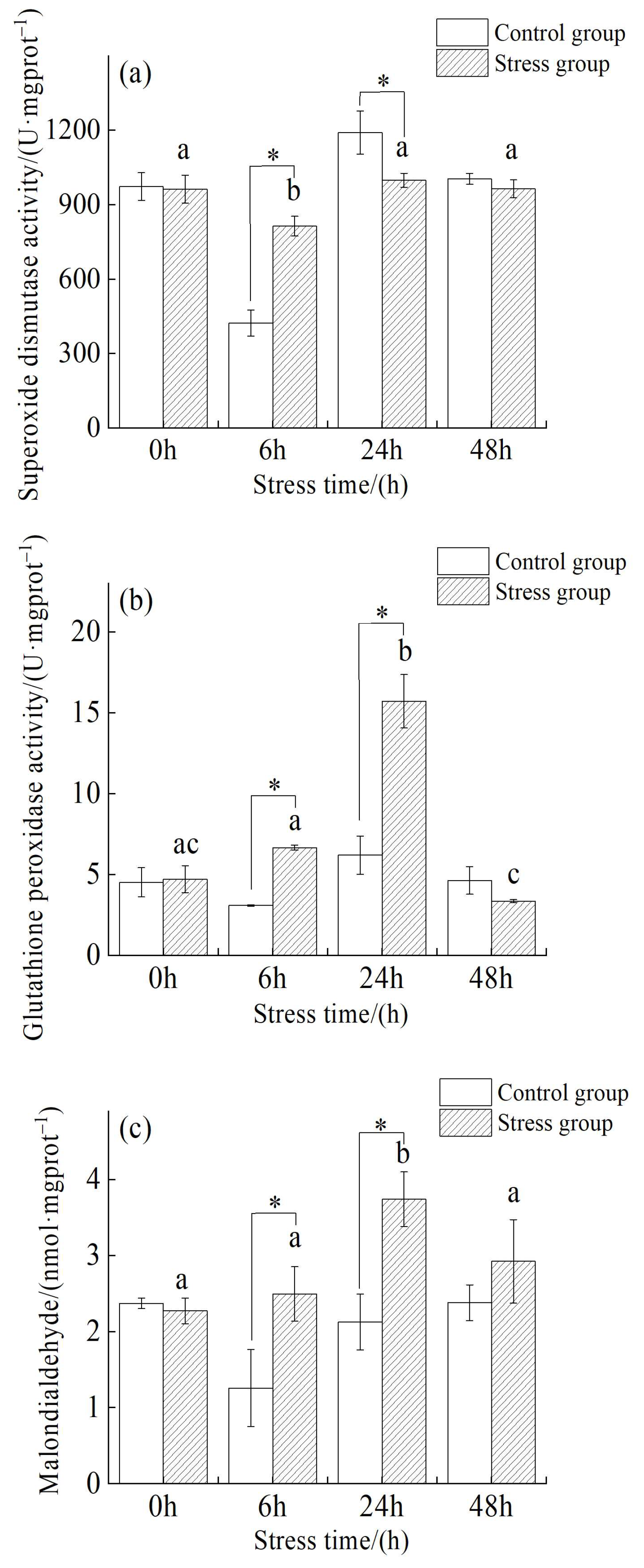
3.3. Changes in Red Muscle Status of Juvenile Yellowfin Tuna under Salinity Stress
3.4. Changes in White Muscle Status of Juvenile Yellowfin Tuna under Salinity Stress
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bisby, F.; Roskov, Y.; Culham, A.; Orrell, T.; Nicolson, D.; Paglinawan, L.; Bailly, N.; Appeltans, W.; Kirk, P.; Bourgoin, T.; et al. (Eds.) Species 2000 & ITIS Catalogue of Life, 2012 Annual Checklist; Reading: London, UK, 2012. [Google Scholar]
- Murua, H.; Rodriguez-Marin, E.; Neilson, J.D.; Farley, J.H.; Juan-Jordá, M.J. Fast versus slow growing tuna species: Age, growth, and implications for population dynamics and fisheries management. Rev. Fish Biol. Fish. 2017, 27, 733–773. [Google Scholar] [CrossRef]
- Zhipan, T.; Fei, W.; Siquan, T.; Qiuyun, M. Stock assessment for Atlantic yellowfin tuna based on extended surplus production model considering life history. Acta Oceanol. Sin. 2022, 41, 41–51. [Google Scholar]
- Weifeng, Z.; Huijuan, H.; Wei, F.; Shaofei, J. Impact of Abnormal Climatic Events on the CPUE of Yellowfin Tuna Fishing in the Central and Western Pacific. Sustainability 2022, 14, 1217. [Google Scholar]
- Benetti, D.D.; Partridge, G.J.; Stieglitz, J. Overview on status and technological advances in tuna aquaculture around the world. In Advances in Tuna Aquaculture; Benetti, D.D., Partridge, G.J., Buentello, A., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 1–19. [Google Scholar]
- Cobcroft, J. Addressing causes of early mortality in hatchery produced southern bluefin tuna larvae. Chapter 9: The effects of in-tank lighting on the early behaviour and performance of yellowtail kingfish larvae. Agric. Vet. Sci. 2014, 96232. [Google Scholar]
- Nguyen, K.Q.; Phan, H.T. Length-length, Length-weight, and Weight-weight Relationships of Yellowfin (Thunnus albacares) and Bigeye (Thunnus Obesus) Tuna Collected from the Commercial Handlines Fisheries in the South China Sea. Thalassas 2022, 38, 911–917. [Google Scholar] [CrossRef]
- Liu, H.; Fu, Z.; Zhou, S.; Hu, J.; Yang, R.; Yu, G.; Ma, Z. The Complete Mitochondrial Genome of Pennella sp. Parasitizing Thunnus alabacares. Front. Cell. Infect. Microbiol. 2022, 12, 945152. [Google Scholar] [CrossRef]
- Katz, S.L.; Syme, D.A.; Shadwick, R.E. Enhanced power in yellowfin tuna. Nature 2001, 410, 770–771. [Google Scholar] [CrossRef]
- Ern, R.; Huong DT, T.; Cong, N.V.; Bayley, M.; Wang, T. Effect of salinity on oxygen consumption in fishes: A review. J. Fish Biol. 2014, 84, 1210–1220. [Google Scholar] [CrossRef]
- Livingstone, D.R. Contaminant-stimulated Reactive Oxygen Species Production and Oxidative Damage in Aquatic Organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Wang, J.Q.; Lui, H.; Po, H.; Fan, L. Influence of salinity on food consumption, growth and energy conversion efficiency of common carp (Cyprinus carpio) fingerlings. Aquaculture 1997, 148, 115–124. [Google Scholar] [CrossRef]
- Choi, C.Y.; An, K.W.; An, M.I. Molecular characterization and mRNA expression of glutathione peroxidase and glutathione S-transferase during osmotic stress in olive flounder (Paralichthys olivaceus). Comp. Biochem. Physiol. Part A 2008, 149, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, W.-N.; Wang, A.-L.; Wang, J.-M.; Sun, R.-Y. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei (Boone, 1931) exposed to acute salinity changes. Aquaculture 2007, 265, 351–358. [Google Scholar] [CrossRef]
- Jiang, Z.; Huang, X.; Zhang, J. Dynamics of nonstructural carbohydrates in seagrass Thalassia hemprichii and its response to shading. Acta Oceanol. Sin. 2013, 32, 61–67. [Google Scholar] [CrossRef]
- Artetxe-Arratea, I.; Frailea, I.; Marsac, F.; Farley, J.H.; Rodriguez-Ezpeleta, N.; Davies, C.R.; Clear, N.P.; Grewed, P.; Murua, H. A review of the fisheries, life history and stock structure of tropical tuna (skipjack Katsuwonus pelamis, yellowfin Thunnus albacares and bigeye Thunnus obesus) in the Indian Ocean. Adv. Mar. Biol. 2020, 88, 39–89. [Google Scholar]
- Lushchak, V.I.; Bagnyukova, T.V.; Lushchak, O.V.; Storey, J.M.; Storey, K.B. Hypoxia and recovery perturb free radical processes and antioxidant potential in common carp (Cyprinus carpio) tissues. Int. J. Biochem. Cell Biol. 2005, 37, 1319–1330. [Google Scholar] [CrossRef]
- Canli, E.G.; Gülüzar Atli Eroglu, A.; Atli, G.; Canli, M. Effects of fish size on the response of antioxidant systems of Oreochromis niloticus following metal exposures. Fish Physiol. Biochem. 2014, 40, 1083–1091. [Google Scholar]
- Vasseur, P.; Rodius, F.; Doyen, P.; Rodius, F. Molecular cloning and expression study of pi-class glutathione S-transferase (pi-GST) and selenium-dependent glutathione peroxidase (Se-GSH-Px) transcripts in the freshwater bivalve Dreissena polymorpha. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2008, 147, 69–77. [Google Scholar]
- Sinha, A.K.; Abd Elgawad, H.; Zinta, G.; Dasan, A.F.; Rasoloniriana, R.; Asard, H.; Blust, R.; De Boeck, G. Nutritional Status as the Key Modulator of Antioxidant Responses Induced by High Environmental Ammonia and Salinity Stress in European Sea Bass (Dicentrarchus labrax). PLoS ONE 2015, 10, e0135091. [Google Scholar] [CrossRef] [Green Version]
- Tao, F.; Wei, Z.; Wan, H.; Rong, C. Effects of benzo(a)pyrene exposure on glutathione peroxidase activity in the liver of Boleophthalmus pectinirostris. J. Fish. Sci. China 2000, 7, 19–21. [Google Scholar]
- Pipe, R.K.; Porte, C.; Livingstone, D.R. Antioxidant Enzymes Associated with the Blood Cells and Hemolymph of the Mussel Mytilus edulis. Fish Shellfish. Immunol. 1993, 3, 221–233. [Google Scholar] [CrossRef]
- Klein, R.D.; Rosa, C.E.; Colares, E.P.; Robaldo, R.B.; Martinez, P.E.; Bianchini, A. Antioxidant defense system and oxidative status in Antarctic fishes: The sluggish rockcod Notothenia coriiceps versus the active marbled notothen Notothenia rossii. J. Therm. Biol. 2017, 68 Pt A, 119–127. [Google Scholar] [CrossRef]
- Gan, L.; Xu, Z.X.; Ma, J.J.; Xu, C.; Wang, X.D.; Chen, K.; Li, E.C. Effects of salinity on growth, body composition, muscle fatty acid composition, and antioxidant status of juvenile Nile tilapia Oreochromis niloticus (Linnaeus, 1758). J. Appl. Ichthyol. 2016, 32, 372–374. [Google Scholar] [CrossRef]
- Yokota, S.U. Regulation of the ion-transporting mitochondrion-rich cell during adaptation of teleost fishes to different salinities. Zool. Sci. 2001, 18, 1163–1174. [Google Scholar]
- Saoud, I.P.; Kreydiyyeh, S.; Chalfoun, A.; Fakih, M. Influence of salinity on survival, growth, plasma osmolality and gill Na+-K+-ATPase activity in the rabbitfish Siganus rivulatus. J. Exp. Mar. Biol. Ecol. 2007, 348, 183–190. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, X.; Dong, C.; Band, Z.; Li, L.; Yub, J.; Hu, Y.; Chen, C. Effects of ozone treatment on SOD activity and genes in postharvest cantaloupe. RSC Adv. 2020, 10, 17452–17460. [Google Scholar] [CrossRef] [PubMed]
- Hahn, O.; Drews, L.F.; An, N.; Tatsuta, T.; Gkioni, L.; Hendrich, O.; Zhang, Q.; Langer, T.; Pletcher, S.; Wakelam, M.J.O.; et al. A nutritional memory impairs survival, transcriptional and metabolic response to dietary restriction in old mice. Cold Spring Harb. Lab. 2019. [Google Scholar] [CrossRef]
- An, M.I.; Choi, C.Y. Activity of antioxidant enzymes and physiological responses in ark shell, Scapharca broughtonii, exposed to thermal and osmotic stress: Effects on hemolymph and biochemical parameters. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 155, 34–42. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant Defenses in Fish: Biotic and Abiotic Factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Zoysa, M.D.; Whang, I.; Lee, Y.; Lee, S.; Lee, J.S.; Lee, J. Transcriptional analysis of antioxidant and immune defense genes in disk abalone (Haliotis discus discus) during thermal, low-salinity and hypoxic stress. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 154, 387–395. [Google Scholar] [CrossRef]
- Wilhelm, D.; Giulivi, C.; Boveris, A. Antioxidant defenses in marine fish I.Teleosts. Comp. Biochem. Physiol. Part C Comp. Pharmacol. Toxicol. 1993, 106, 409–413. [Google Scholar]
- Vinagre, C.; Madeira, D.; Narciso, L.; Cabral, H.N.; Diniz, M. Effect of temperature on oxidative stress in fish: Lipid peroxidation and catalase activity in the muscle of juvenile seabass, Dicentrarchus labrax. Ecol. Indic. 2012, 23, 274–279. [Google Scholar] [CrossRef]
- Liao, Y.; Zhang, C.; Peng, S.; Gao, Q.; Shi, Z. Effects of salinity on activities of liver antioxidant enzymes and plasma lysozyme of Epinehelus moara. J. Shanghai Ocean. Univ. 2016, 25, 169–176. [Google Scholar]
- Bian, P.J.; Qiu, C.G.; Shan-Liang, X.U.; Lin, S. Effects of salinity on growth, activity of non-specific immune and antioxidant enzymes in obscure puffer Takifugu obscures. Acta Hydrobiol. Sin. 2014, 38, 108–114. [Google Scholar]
- Wang, R.P.; Dai, L.L.; Chen, Y.F. Effects of short-term temperature or salinity stress on feeding behavior and antioxidant of marine medaka(oryzias melastigma). Oceanol. Et Limnol. Sin. 2019, 10, 730853. [Google Scholar]
- Mak, Y.L.; Jia, J.W.; Chan, W.H.; Murphy, M.B.; Lam, J.C.; Chan, L.L.; Lam, P.K. Simultaneous quantification of Pacific ciguatoxins in fish blood using liquid chromatography-tandem mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 3331–3340. [Google Scholar] [CrossRef]
- Kalinina, E.V.; Chernov, N.N.; Novichkova, M.D. Role of Glutathione, Glutathione Transferase, and Glutaredoxin in Regulation of Redox-Dependent Processes. Biochemistry 2014, 79, 1562–1583. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.P. (Ed.) Encyclopedia of Fish Physiology: From Genome to Environment; Academic Press: London, UK, 2011. [Google Scholar]
- Xiaohong, L. Preliminary Study on the Effect of Water Cadmium Exposure on Liver Toxicity and Lipid Metabolism of Rare Gudgeon Crucian Carp; Southwest University: Chongqing, China, 2016. [Google Scholar]
- Zhao, F.; Zhuang, P.; Zhang, L.Z.; Huang, X.R.; Zhang, T.; Feng, G.P. Responses of antioxidases in different tissues of Acipenser schrenckii to increased salinity in water. Mar. Fish. Res. 2008, 29, 65–69. [Google Scholar]
- Yang, J.; Chen, G.; Huang, J.S.; Zhang, J.D.; Shi, G.; Tang, B.G.; Zhou, H. Effects of Temperature and Salinity on the Growth and Activities of Antioxidant Enzymes of Cobia (Rachycentron canadum) Juveniles. J. Guangdong Ocean. Univ. 2007, 4, 24–29. [Google Scholar]
- Davenport, J. Osmotic control in marine animals. Symp. Soc. Exp. Biol. 1985, 39, 207–244. [Google Scholar]
- Michael G, J.; Giacomo, Z. Nervous control of the gills. Acta Histochem. 2009, 111, 207–216. [Google Scholar]
- Ghanavatinasab, Y.; Salati, A.P.; Movahedinia, A.; Shahriari, A. Changes in Gill Antioxidant Status in Acanthopagrus sheim Exposed to Different Environmental Salinities. Iran. J. Sci. Technology. Trans. A Sci. 2018, 43, 1479–1483. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Y.; Gao, Q.; Shiming, P.E.N.G.; Zhaohong, S.H.I. Effect of low salinity stress on antioxidant function in liver of juvenile Nibea albiflora. South China Fish. Sci. 2015, 11, 59–64. [Google Scholar]
- Garcia, D.; Lima, D.; Silva, D.; de Almeida, E.A. Decreased malondialdehyde levels in fish (Astyanax altiparanae) exposed to diesel: Evidence of metabolism by aldehyde dehydrogenase in the liver and excretion in water. Ecotoxicol. Environ. Saf. 2020, 190, 110107. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, S.; Zhang, N.; Fu, Z.; Yu, G.; Ma, Z.; Zhao, L. Impact of Salinity Changes on the Antioxidation of Juvenile Yellowfin Tuna (Thunnus albacares). J. Mar. Sci. Eng. 2023, 11, 132. https://doi.org/10.3390/jmse11010132
Zhou S, Zhang N, Fu Z, Yu G, Ma Z, Zhao L. Impact of Salinity Changes on the Antioxidation of Juvenile Yellowfin Tuna (Thunnus albacares). Journal of Marine Science and Engineering. 2023; 11(1):132. https://doi.org/10.3390/jmse11010132
Chicago/Turabian StyleZhou, Shengjie, Ninglu Zhang, Zhengyi Fu, Gang Yu, Zhenhua Ma, and Lei Zhao. 2023. "Impact of Salinity Changes on the Antioxidation of Juvenile Yellowfin Tuna (Thunnus albacares)" Journal of Marine Science and Engineering 11, no. 1: 132. https://doi.org/10.3390/jmse11010132