Abstract
Marine dissolved organic matter (DOM) is a key component of the global carbon cycle. While marine DOM properties are usually characterized by chemical approaches, an effort to integrate its variations in an ecological perspective is needed. Heterotrophic prokaryotes being the main consumers of marine DOM, targeting their responses to varying DOM properties provides an effect-based characterization of DOM properties. Dilution experiments of natural marine prokaryotic assemblages were used to assess the spatio-temporal heterogeneity of DOM properties in the coastal environment through its potential to stimulate heterotrophic prokaryotic growth. Reduced top-down pressures allowed to repeatedly evidence a stronger bottom-up stimulation potential of heterotrophic prokaryotes’ net growth with DOM collected in harbours under strong human and continental influence compared to DOM collected in more open coastal sites. Shedding the light on high spatial heterogeneity of DOM properties that corresponded only partly to dissolved organic carbon concentrations, this experimental approach represents a simple and reproducible method to improve our understanding of the marine DOM cycle.
1. Introduction
Marine dissolved organic matter (DOM) represents a dynamic component in the interactions between geosphere, hydrosphere and biosphere and, as such, has the potential to influence the global carbon cycle [1]. Heterotrophic prokaryotes (HP) are considered as the major consumers and mineralizers of DOM in the ocean, thus initiating the microbial loop [2]. Forty PgC are recycled each year by the microbial loop which demonstrates that HP represent a very dynamic compartment in global biogeochemical cycles. Prokaryotic DOM uptake induces different processes, assimilation and mineralization of DOM being two essential processes in the marine DOM cycle [3]. Their relative contributions are influenced by DOM properties [4].
In coastal areas, the ecosystem is exposed to rapid changes in environmental conditions and human pressures. Numerous studies have also demonstrated that primary production by phytoplankton is stimulated in coastal areas feeding the base of the heterotrophic food web [5,6]. Interestingly, Coclet et al. [7,8] observed the selection of prokaryotic taxa adapted to a greater availability of nutritive resources in the most enclosed and chemically contaminated sites of an urbanized Mediterranean bay, suggesting intriguing trophic responses of HP to coastal human influences. Considering the human footprint on the Mediterranean Sea shore line [9], a better evaluation of human influence on marine DOM and its cycling appears of particular importance. For that purpose, it is required to get a simple and reliable method to evaluate the effect of spatio-temporal variations of DOM properties on the heterotrophic growth in the highly variable coastal area.
Several chemical and biological methods exist to assess the effect of DOM properties on heterotrophic microorganisms. Biological methods allow measuring bacterial production and bacterial growth, using the incorporation of radiolabelled tracers into stable macromolecules, in particular DNA, RNA, proteins or phospholipids. The main drawbacks of these techniques are (i) they require special equipment for the use of radioactive or stable isotopes and (ii) they involve the use of several conversion factors that are not easy to estimate accurately [10]. Among the chemical methods, the measurement of dissolved organic carbon (DOC) consumption by heterotrophic microorganisms is the most widely used, but it has several drawbacks, particularly for its application to the ocean environment [11]. Indeed, the low concentrations of DOC observed in the ocean, especially in oligotrophic area, require long incubation times of several days to observe a significant consumption, even with highly precise equipment.
In this study, we proposed a simple, rapid and reproducible method to compare the effect of DOM properties on the growth of heterotrophic prokaryotes from different water samples. The method is based on the potential of the pool of dissolved substances to stimulate HP growth in the absence of significant top-down pressure. It consists of exposing natural microbial communities to various DOM pools in dilution experiments (also referred to as seawater culture) [12] with short-term growth monitoring by flow cytometry. The method requires very few manipulations and is based on ecological responses, with the use of natural complex communities. It was used for a first assessment of the spatial heterogeneity of dissolved substances in the coastal north-west Mediterranean Sea. Based on previous studies, we hypothesized that DOM of the enclosed harbours within the strongly urbanized and land-connected Toulon Bay could sustain more efficiently the heterotrophic base of the marine planktonic food web. We repeatedly observed a stimulation of HP growth with the water of the harbour compared to more open coastal areas. This difference in biological responses only partly corresponded to DOC variations, proving the usefulness of this easy-to-use method to complement classical chemical characterization of DOM.
2. Materials and Methods
Two different water bodies were considered on the French coast of the NW Mediterranean Sea: (i) enclosed waters from a large harbour within the strongly urbanized and land-connected Toulon Bay (further referred to as H), and (ii) open coastal waters under little anthropogenic and continental influences sampled at the mouth or outside from the bay (furthered referred to as O). Toulon Bay was chosen because of its numerous human activities, resulting in chemical contaminations gradients and associated HP abundance and diversity gradients previously observed in the water column on a restricted geographical scale [7,13,14]. Both areas were sampled at three distinct dates (March, July and November 2020) to presumably cover different seasonal variations in water composition, in terms of both DOM pool and prokaryotic community composition. Although the comparison between an enclosed urbanized station and a more open to the sea station was always preserved, the COVID-19 outbreak did not allow us to access the same harbour or to use boats for sampling off the coast during the different lockdowns decided in France. This forced us to choose different sampling stations at the different sampling dates (Figure 1). Since the aim of our study was not to compare sites but rather to use contrasted DOM pools to validate the usefulness of the methods, we believe that sampling different stations does not alter our conclusions. The different H stations are located in enclosed docs of Toulon harbour area and are, therefore, subject to strong anthropogenic pressures and influence of freshwater loadings. All three O stations were all under weak anthropogenic influence, either being located off the coast (ONOV, OMAR) in a rather open area or along the coast (OJUL), without any major human activity in their close vicinity and more exposed to sea currents than H stations. Station OJUL was located farther in the east to access a rocky shoreline, with very few human frequentation, within a marine-protected area (Port-Cros National Park).

Figure 1.
Location of the sampling stations. H: Harbour area under high anthropogenic and continental influences. O: open area under low anthropogenic and continental influences. Mar, Jul and Nov: sampling campaigns of March, July and November, respectively.
To avoid any contamination during the sampling and the experiments, all the material used was previously washed three times with Milli-Q water, then with 10% (v/v) hydrochloric acid (HCl, Analytical Grade, Fisher Scientific, Leicester, UK) and finally rinsed three times with Milli-Q water. The material was then conditioned with the target sample. At each station and each date, a unique sample of 2 L of surface water (−1 m) was collected with a van Dorn horizontal sampler and transferred in fluorinated ethylene propylene bottle (FEP, Nalgene, Fisher Scientific, Leicester, UK). Samples were brought back in the lab in a cool box and processed within half a day in order to avoid any major change in dissolved substances properties or prokaryotic communities.
The initial quantity of DOM was estimated at each site and for each sampling date by measuring DOC concentrations. For that, 24 mL samples filtered through a 0.2 µm PES syringe filter were collected in previously conditioned glass tubes and stored at 4 °C. DOC concentrations were determined by high temperature catalytic oxidation using a Shimadzu TOC-VCSH carbon analyser with an accuracy of 1.2 µM. Prior to oxidation, samples were acidified with 2 M high purity HCl and purged for 3 min with O2 to remove inorganic carbon. In order to obtain satisfactory analytical precision (±1%), up to 5 repeated injections were performed. Every 8 samples, the system blank was measured with Milli-Q water and the operation of the instrument was verified by comparing data with DOC Consensus Reference Material (SUPER-05) [15,16].
For each water sample, a 0.2 µm mixed cellulose esters membrane (Whatman, 47 mm) was rinsed with 100 mL of 10% (v/v) hydrochloric acid (HCl, Analytical Grade, Fisher Scientific, Leicester, UK) and 1 L of Milli-Q water to avoid any significant contamination of the filtrate. Then, 500 mL of seawater was filtered through to isolate the dissolved substances’ pool in the filtrate. Similarly, a glass fibre filter (GF/C, Whatman, 1.2 µm, 47 mm) was calcinated (450 °C, 6 h) before being rinsed with Milli-Q water and the targeted sample. Then, 100 mL of raw seawater was filtered to isolate the free-living HP community in the filtrate while removing most of the potential grazers. Dilution experiments were then conducted by exposing each isolated HP community to each isolated dissolved substance pool, resulting in 4 experimental conditions at each sampling date (Figure 2). Because of a conservation issue after the July sampling campaign, we were unable to use the community from H in the incubation experiments. At this date, only the data with the community from O could be acquired.
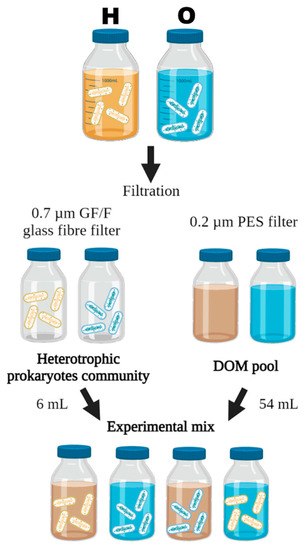
Figure 2.
Schematic representation of the experimental design. At each sampling date, two seawater samples were taken from two contrasted stations. The heterotrophic prokaryotic communities and the pool of dissolved substances were isolated by filtration through 0.7 µm and 0.2 µm filters, respectively. Experimental mixtures were then made by exposing each community to each pool of dissolved substances, in triplicates. H and O represent raw waters from the harbours and the open sites, respectively.
The different experimental conditions were performed in triplicate. For each replicate, 54 mL of isolated dissolved substances (<0.2 µm) was inoculated with 6 mL of free-living HP (<1.2 µm) for 48 h at 20 °C in a 60 mL FEP bottle (Figure 2). These incubations were carried out under conditions promoting HP growth (removal of most of the grazers, reduced competition for resources and viral pressure by dilution) and inhibiting any fresh DOM production by phytoplankton production (darkness). Diluting the prokaryotes allowed us to expose them to a large majority of dissolved substances originating from another water sample, without manipulating them too much. This dilution also maximized the analytical precision of the growth estimations while establishing more homogeneous competition and growth conditions between the different incubations.
To measure bacterial growth, 1 mL of subsamples was taken from each bottle at the initial and final time of incubation, fixed with 0.25% glutaraldehyde (final concentration) and frozen at −80 °C until further analysis. After thawing, samples were stained with SYBR Green (1× final concentration) for 15 min in darkness and heterotrophic prokaryotes were counted with an Accuri C6 flow cytometer (BD Biosciences, Becton, NJ, USA). Cells considered as HP were smaller than 2.0 µm Fluoresbrite® YG Microspheres (Polysciences Inc., Eppelheim, Germany), exhibited low complexity (low SSC), emitted green fluorescence and no red fluorescence [17]. Data were acquired using BD Accuri CFlow Plus software and HP abundances were expressed as numbers of cells per mL (cell·mL−1). HP abundances were used to calculate net bacterial growth (cell·mL−1·h−1) by subtracting initial abundances from final abundances and dividing by incubation time (48 h).
3. Results and Discussion
In situ variability showed that DOM quantitative properties varied from a spatial but also temporal point of view. The DOC concentration varied between 65.8 and 112.8 µM (Figure 3). On the basis of the sampling campaigns of March and July, DOC concentration tended to significantly increase in the harbour when compared to the open area (p-value < 0.05). Human activities could also contribute to higher DOC concentrations in the harbour area through substantial anthropogenic DOC (ADOC) inputs [18,19]. Although the organic contamination of the water column has not been documented yet in Toulon Bay, the strong organic contamination recorded in the sediments [20,21] and the contamination of the water by sediment resuspension [22,23] do not let any doubt about its occurrence. However, in November, no significant difference in DOC concentration was demonstrated between the harbour area under high anthropogenic pressure and the open area under low anthropogenic pressure, suggesting a temporal variability in human influence on DOM quantitative properties.

Figure 3.
DOC concentration in the sampled waters for each campaign. Error bars represent standard error calculated from replicated measurements. Letters above error bars represent the result of Tukey multiple comparisons, the absence of common letters indicating a significant difference (p-value < 0.05).
Significantly higher DOC concentrations were observed in July relative to March for both sites (p-value < 0.05), while DOC concentrations in July and November were not significantly different (p-value > 0.05). With average concentrations of 71.8, 101.7 and 104.7 µM in March, July and November, respectively, our data are in good agreement with the annual cycle of the DOM in surface waters of the Mediterranean Sea [24,25,26].
With a maximal increase of 34.9 µM for H and 30.9 µM for O, the temporal variability in DOC concentration appeared higher than the spatial one at a given date (maximal significant spatial difference of 11.9 µM observed in March). This suggests a globally higher influence of temporal processes than spatial heterogeneity of sources and water residence time on DOC concentration. Such trend suggests an important control of phytoplanktonic growth and continental inputs on quantitative properties of DOM at rather large scale in the coastal area, in agreement with previous observations on phytoplanktonic dynamics in the studied area [27].
The dilution experiments showed a significant net growth after only 48 h of incubation (Figure 4). HP growth was always significantly higher with dissolved substances of the harbour area, regardless of HP community origin and sampling date. This systematic difference in HP growth suggests that the harbour area hosted more dissolved substances that have a high growth-stimulating potential than open coastal areas under low anthropogenic influences. Based on our observations, heavily anthropized coastal environments such as harbour could be considered as hotspots for heterotrophic secondary production. In March and July, the higher DOC concentration in harbour area could be responsible for the differences in growth stimulation observed. However, significantly higher HP growth can be observed without a significantly higher DOC concentration such as in November where no significant difference in DOC concentration was observed between sites. This observation was comforted by the absence of significant correlation between DOC concentration and net HP growth (p-value > 0.05). Considering similar competitive and top-down pressures in all experimental conditions, this observation suggests that our experimental approach could be able to differentiate DOM pools based on their potential to support heterotrophic growth at the base of the marine planktonic food web, while conventional chemical measurement hardly do so. While previous work monitored HP growth in experimental approaches to decipher its mains controls, it highlighted the predominant influence of grazing by comparing filtered vs. non-filtered conditions [28]. By a systematic dilution of filtered HP community, our approach allowed us to demonstrate the heterogeneity of bottom up constrains in the coastal environment.
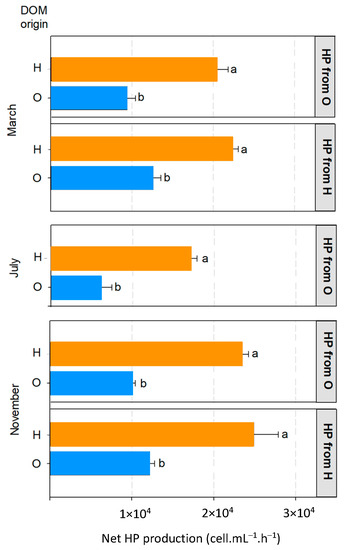
Figure 4.
Net HP growth as a function of DOM origin for each campaign. HP origin is specified on the right-hand side, one panel corresponding to one community. H: harbour. O: open coastal area. Orange bars: DOM from H. Blue bars: DOM from O. Error bars represent the standard deviation between experimental triplicates. a and b represent the result of Tukey post-hocpost hoc multiple comparisons within the same block, the absence of common letters indicating significant difference (p < 0.05).
To conclude, the method presented here provides several advantages: it requires few materials, it is easy to set up, results are obtained quickly with only 2 days of incubation using equipment found in most of oceanography laboratories and, thus, numerous comparisons could be performed at the same time. It allowed us to evidence important spatial differences in the ability of DOM to sustain heterotrophic growth at the basis of marine planktonic food webs. Further systematic deployment along with classical measurements of DOM properties could open interesting perspectives for a causal understanding of the links between DOM fate and microbial loop functioning in marine waters, as well as for a better biogeochemical understanding of human influence on the coastal ocean.
Author Contributions
Conceptualization, C.D., V.L., O.P. and B.M.; methodology, C.D., V.L. and B.M.; formal analysis, C.D.; writing—original draft preparation, C.D.; writing—review and editing, V.L., O.P. and B.M.; funding acquisition, V.L. and B.M. All authors have read and agreed to the published version of the manuscript.
Funding
This project was financially supported by the European Interreg Italy-France Maritime 2014–2020 Project «GEREMIA» (Gestione dei reflui per il miglioramento delle acque portuali), and by Toulon University, the CARTT of the University Institute of Technology of Toulon and Toulon Provence Mediterrannée through project “C-OMICS” (Coupling OMICS approaches for in-depth study of human-induced responses of microbial communities). Clara Dignan received a Ph.D. fellowship from the French Ministère de l’Enseignement Supérieur et de la Recherche and this paper is a part of her Ph.D.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request.
Acknowledgments
The authors are grateful to Bruno Zakardjian, the crew and captain of the R/V Antedon II, the French Navy and the LASEM from Toulon for their contribution to sampling. The authors also thank Benjamin Oursel for his contribution to DOC quantification.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Farrington, J. Overview and key recommendations. Marine organic geochemistry workshop, January 1990. Mar. Chem. 1992, 39, 5–9. [Google Scholar]
- Azam, F. Microbial control of oceanic carbon flux: The plot thickens. Science 1998, 280, 694–696. [Google Scholar] [CrossRef]
- Duclow, H.W.; Kirchman, D.L.; Anderson, T.R. Themagnitude of spring bacterial production in the NorthAtlantic Ocean. Limnol. Oceanogr. 2002, 47, 1684–1693. [Google Scholar] [CrossRef]
- Amon, R.M.W.; Benner, R. Linkages among the bioreactivity, chemical composition, and diagenetic state of marinedissolved organic matter. Limnol. Oceanogr. 2001, 46, 287–297. [Google Scholar] [CrossRef]
- Borum, J.; Sand-Jensen, K. Is total primary production in shallow coastal marine waters stimulated by nitrogen loading? Oikos 1996, 76, 406–410. [Google Scholar] [CrossRef]
- Cloern, J.E.; Foster, S.Q.; Kleckner, A.E. Phytoplankton primary production in the world’s estuarine-coastal ecosystems. Biogeosciences 2014, 11, 2477–2501. [Google Scholar] [CrossRef]
- Coclet, C.; Garnier, C.; Durrieu, G.; Omanovic, D.; D’Onofrio, S.; Le Poupon, C.; Mullot, J.U.; Briand, J.F.; Misson, B. Changes in Bacterioplankton Communities Resulting from Direct and Indirect Interactions with Trace Metal Gradients in an Urbanized Marine Coastal Area. Front. Microbiol. 2019, 10, 257. [Google Scholar] [CrossRef]
- Coclet, C.; Garnier, C.; Durrieu, G.; D’Onofrio, S.; Layglon, N.; Briand, J.F.; Misson, B. Impacts of copper and lead exposure on prokaryotic communities from contaminated contrasted coastal seawaters: The influence of previous metal exposure. FEMS Microbiol. Ecol. 2020, 96, fiaa048. [Google Scholar] [CrossRef]
- United Nations Environment Programme/Mediterranean Action Plan and Plan Bleu. State of the Environment and Development in the Mediterranean: Key Messages; United Nations Environment Programme: Nairobi, Kenya, 2020. [Google Scholar]
- Robarts, R.D. Incorporation of Radioactive Precursors into Macromolecules as Measures of Bacterial Growth: Problems and Pitfalls. In Molecular Approaches to the Study of the Ocean; Cooksey, K.E., Ed.; Springer: Dordrecht, The Nertherland, 1998. [Google Scholar] [CrossRef]
- Volk, C.; Renner, C.; Robert, C.; Joret, J.C. Comparison of two techniques for measuring biodegradable dissolved organic carbon in water. Environ. Technol. 1994, 15, 545–556. [Google Scholar] [CrossRef]
- Ammerman, J.W.; Fuhrman, J.A.; Hagström, A.; Azam, F. Bacterioplankton growth in seawater: I. Growth kinetics and cellular characteristics in seawater cultures. Mar. Ecol. Prog. Ser. 1984, 18, 31–39. [Google Scholar] [CrossRef]
- Layglon, N.; Misson, B.; Durieu, G.; Coclet, C.; D’Onofrio, S.; Dang, D.H.; François, D.; Mullot, J.U.; Mounier, S.; Lenoble, V.; et al. Long-term monitoring emphasizes impacts of the dredging on dissolved Cu and Pb contamination along with ultrplankton distribution and structure in Toulon Bay (NW Mediterranean Sea, France). Mar. Pollut. Bull. 2020, 156, 111196. [Google Scholar] [CrossRef] [PubMed]
- Paix, B.; Layglon, N.; Le Poupon, C.; D’Onofrio, S.; Misson, B.; Garnier, C.; Culioli, G.; Briand, J.F. Integration of spatio-temporal variations of surface metabolomes and epibacterial communities highlights the importance of copper stress as a major factor shaping host-microbiota interactions within a Mediterranean seaweed holobiont. Microbiome 2021, 9, 201. [Google Scholar] [CrossRef] [PubMed]
- Louis, Y.; Garnier, C.; Lenoble, V.; Mounier, S.; Cukrov, N.; Omanović, D.; Pižeta, I. Kinetic and equilibrium studies of copper-dissolved organic matter complexation in water column of the stratified Krka River estuary (Croatia). Mar. Chem. 2009, 114, 110–119. [Google Scholar] [CrossRef]
- Hansell, D.A. Dissolved Organic Carbon Reference Material Program. Eos Trans. Am. Geophys. Union 2005, 86, 318. [Google Scholar] [CrossRef]
- Grégori, G.; Citterio, S.; Ghiani, A.; Labra, M.; Sgorbati, S.; Brown, S.; Denis, M. Resolution of viable and membrane-compromised bacteria in freshwater and marine waters based on analytical flow cytometry and nucleic acid double staining. Appl. Environ. Microbiol. 2001, 67, 4662–4670. [Google Scholar] [CrossRef]
- Sachse, A.; Henrion, R.; Gelbrecht, J.; Steinberg, C.E.W. Classification of dissolved organic carbon (DOC) in river systems: Influence of catchment characteristics and autochthonous processes. Org. Geochem. 2005, 36, 923–935. [Google Scholar] [CrossRef]
- Wen, Z.; Song, K.; Shang, Y.; Lyu, L.; Tao, H.; Liu, G. Natural and anthropogenic impacts on the DOC characteristics in the Yellow River continuum. Environ. Pollut. 2021, 287, 117231. [Google Scholar] [CrossRef]
- Misson, B.; Garnier, C.; Lauga, B.; Dang, D.H.; Ghiglione, J.F.; Mullot, J.U.; Duran, R.; Pringault, O. Chemical multi-contamination drives benthic prokaryotic diversity in the anthropized Toulon Bay. Sci. Total Environ. 2016, 556, 319–329. [Google Scholar] [CrossRef]
- Wafo, E.; Abou, L.; Nicolay, A.; Boissery, P.; Garnier, C.; Portugal, H. Historical trends of polycyclic aromatic hydrocarbons (PAHs) in the sediments of Toulon Bay (South of France). Int. J. Environ. Monit. Anal. 2017, 5, 150–158. [Google Scholar] [CrossRef]
- Guigue, C.; Tedetti, M.; Dang, D.H.; Mullot, J.U.; Garnier, C.; Goutx, M. Remobilization of polycyclic aromatic hydrocarbons and organic matter in seawater during sediment resuspension experiments from a polluted coastal environment: Insights from Toulon Bay (France). Environ. Pollut. 2017, 229, 627–638. [Google Scholar] [CrossRef]
- Layglon, N.; Lenoble, V.; Longo, L.; D’Onofrio, S.; Mounier, S.; Mullot, J.U.; Sartori, D.; Omanovic, D.; Garnier, C.; Misson, B. Cd transfers during marine sediment resuspension over short and long-term period: Associated risk for coastal water quality. Mar. Pollut. Bull. 2022, 180, 113771. [Google Scholar] [CrossRef]
- Copin-Montégut, G.; Avril, B. Vertical distribution and temporal variation of dissolved organic carbon in the North-Western Mediterranean Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 1993, 40, 1963–1972. [Google Scholar] [CrossRef]
- Avril, B. DOC dynamics in the northwestern Mediterranean Sea (DYFAMED site). Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 2163–2182. [Google Scholar] [CrossRef]
- Pujo-Pay, M.; Conan, P. Seasonal variability and export of dissolved organic nitrogen in the northwestern Mediterranean Sea. J. Geophys. Res. Ocean. 2003, 108, 3188. [Google Scholar] [CrossRef]
- Delpy, F.; Serranito, B.; Jamet, J.L.; Grégori, G.; Le Poupon, C.; Jamet, D. Pico-and nanophytoplankton dynamics in two coupled but contrasting coastal bays in the NW Mediterranean Sea (France). Estuaries Coasts 2018, 41, 2039–2055. [Google Scholar] [CrossRef]
- Silva, L.; Calleja, M.L.; Huete-Stauffer, T.M.; Ivetic, S.; Ansari, M.I.; Viegas, M.; Morán, X.A.G. Low abundances but high growth rates of coastal heterotrophic bacteria in the Red Sea. Front. Microbiol. 2019, 9, 3244. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).