
Plankton Resting Stages Distribution in Bottom Sediments along the Confinement Gradient of the Taranto Sea System (Ionian Sea, Southern Italy)




Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Sampling
2.4. Laboratory Procedures and Analyses
2.5. Statistical Analyses
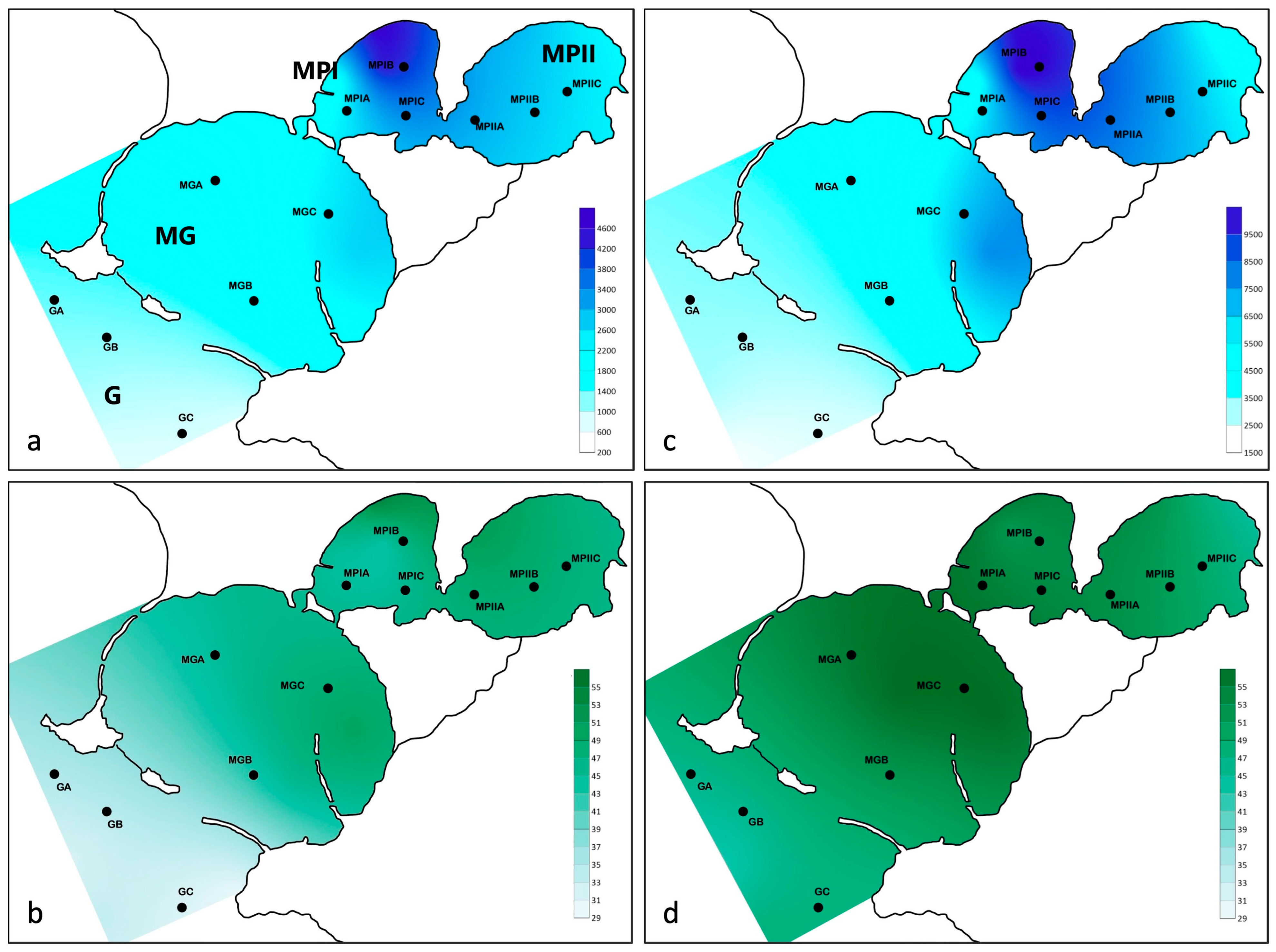
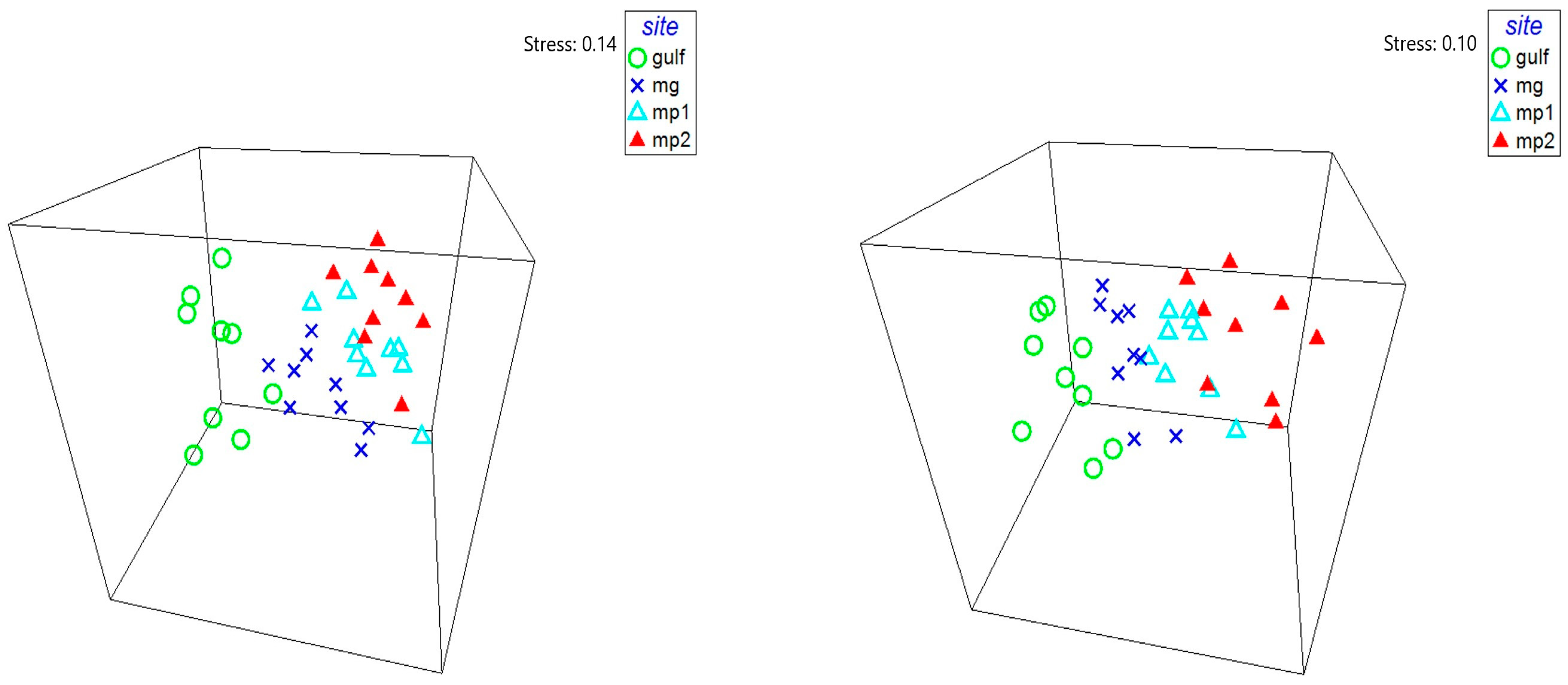
3. Results
3.1. Abiotic Features of the Study Area
3.2. Cyst Bank Composition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belmonte, G.; Castello, P.; Piccinni, M.R.; Quarta, S.; Rubino, F.; Geraci, S.; Boero, F. Resting stages in marine sediments off the Italian coast. In Biology and Ecology of Shallow Coastal Waters; Elefteriou, A., Ansel, A.D., Smith, C.J., Eds.; Olsen and Olsen: Fredensborg, Denmark, 1995; pp. 53–58. [Google Scholar]
- Belmonte, G.; Rubino, F. Resting cysts from coastal marine plankton. Oceanogr. Mar. Biol. Annu. Rev. 2019, 57, 1–88. [Google Scholar]
- Guelorget, O.; Perthuisot, J.P. Paralic ecosystems—Biological organization and functioning. Vie Milieu 1992, 42, 215–251. [Google Scholar]
- Rice, E.; Dam, H.G.; Stewart, J. Impact of climate change on estuarine zooplankton: Surface water warming in Long Island Sound is associated with changes in copepod size and community structure. Estuaries Coasts 2015, 38, 13–23. [Google Scholar] [CrossRef]
- Ikeda, T. Relationship between respiration rate and body size in marine plankton animals as a function of the temperature of habitat. Bull. Fac. Fish. Hokkaido Univ. 1970, 21, 91–112. [Google Scholar]
- Uye, S.-I. Replacement of large copepods by small ones with eutrophication of embayments: Cause and consequence. Hydrobiologia 1994, 292, 513–519. [Google Scholar] [CrossRef]
- Hart, R.C.; Bychek, E.A. Body size in freshwater planktonic crustaceans: An overview of extrinsic determinants and modifying influences of biotic interactions. Hydrobiologia 2011, 668, 61–108. [Google Scholar] [CrossRef]
- Belmonte, G.; Cavallo, A. Body size and its variability in the copepod Acartia margalefi (Calanoida) from lake Acquatina (SE Italy). Ital. J. Zool. 1997, 64, 377–382. [Google Scholar] [CrossRef]
- Gyllström, M.; Hansson, L.-A. Dormancy in freshwater zooplankton: Induction, termination and the importance of benthic-pelagic coupling. Aquat. Sci. 2004, 66, 274–295. [Google Scholar] [CrossRef]
- Marcus, N.H.; Boero, F. Production and plankton community dynamics in coastal aquatic systems: The importance of benthic pelagic coupling in the forgotten role of life cycles. Limnol. Oceanogr. 1998, 43, 763–768. [Google Scholar] [CrossRef]
- Boero, F.; Belmonte, G.; Fanelli, G.; Piraino, S.; Rubino, F. The continuity of living matter and the discontinuities of its constituents: Do plankton and benthos really exist? Trends Ecol. Evol. 1996, 11, 177–180. [Google Scholar] [CrossRef]
- Leck, M.A.; Parker, V.Y.; Simpson, R.L. Ecology of Soil Seed Banks; Academic Press Inc.: San Diego, CA, USA, 1989; 445p. [Google Scholar]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, G.; Vaglio, I.; Rubino, F.; Alabiso, G. Zooplankton composition along the confinement gradient of the Taranto Sea System (Ionian Sea, south-eastern Italy). J. Mar. Syst. 2013, 128, 222–238. [Google Scholar] [CrossRef]
- Rubino, F.; Belmonte, G. Habitat Shift for Plankton: The Living Side of Benthic-Pelagic Coupling in the Mar Piccolo of Taranto (Southern Italy, Ionian Sea). Water 2021, 13, 3619. [Google Scholar] [CrossRef]
- García-Moreiras, I.; Oliveira, A.; Santos, A.I.; Oliveira, P.B.; Amorim, A. Environmental Factors Affecting Spatial Dinoflagellate Cyst Distribution in Surface Sediments off Aveiro-Figueira da Foz (Atlantic Iberian Margin). Front. Mar. Sci. 2021, 8, 699483. [Google Scholar] [CrossRef]
- Kerfoot, W.C.; Weider, L.J. Experimental paleoecology (resurrection ecology): Chasing Van Valen’s Red Queen hypothesis. Limnol. Oceanogr. 2004, 49, 1300–1310. [Google Scholar] [CrossRef]
- Rodrigues, R.V.; Patil, J.S. Salinity changes may influence dinoflagellate cyst morphometry: Data from monsoon-influenced tropical coastal ecosystems. J. Plankton Res. 2021, 43, 853–864. [Google Scholar] [CrossRef]
- Ferraro, L.; Rubino, F.; Belmonte, M.; Prato, S.D.; Greco, M.; Frontalini, F. A multidisciplinary approach to study confined marine basins: The holobenthic and merobenthic assemblages in the Mar Piccolo of Taranto (Ionian Sea, Mediterranean). Mar. Biodiv. 2017, 47, 887–911. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Lei, M.; Ji, S.; Chen, J.; Zheng, H.; Tang, Y.; Hu, R. Distribution of Dinoflagellate cysts in surface sediments from the Qingdao coast, the Yellow Sea, China: The potential risk of harmful algal blooms. Front. Mar. Sci. 2022, 9, 910327. [Google Scholar] [CrossRef]
- Di Leo, A.; Annicchiarico, C.; Cardellicchio, N.; Giandomenico, S.; Conversano, M.; Castellano, G.; Basile, F.; Martinelli, W.; Scortichini, G.; Spada, L. Monitoring of PCDD/Fs and dioxin-like PCBs and seasonal variations in mussels from the Mar Grande and the Mar Piccolo of Taranto (Ionian Sea, Southern Italy). Environm. Sci. Poll. Res. 2014, 21, 13196–13207. [Google Scholar] [CrossRef]
- Vatova, A.R. La produttività delle acque del Mar Grande e del Mar Piccolo di Taranto (1962–1969). Boll. Pes. Pis. Idrobiol. 1972, 27, 81–103. [Google Scholar]
- De Pascalis, F.; Petrizzo, A.; Ghezzo, M.; Lorenzetti, G.; Manfè, G.; Alabiso, G.; Zaggia, L. Estuarine circulation in the Taranto Seas. Environm. Sci. Poll. Res. 2016, 23, 12515–12534. [Google Scholar] [CrossRef] [PubMed]
- Montresor, M.; Bastianini, M.; Cucchiari, E.; Giacobbe, M.; Penna, A.; Rubino, F.; Satta, C.T. Stadi di resistenza del plancton. In Metodologie di Studio del Plancton Marino, ISPRA—Istituto Superiore per la Protezione e la Ricerca Ambientale; Socal, G., Buttino, I., Cabrini, M., Mangoni, O., Penna, A., Totti, C., Eds.; Institute for Environmental Protection and Research (ISPRA): Rome, Italy, 2010; pp. 271–285. [Google Scholar]
- Belmonte, G.; Moscatello, S.; Rubino, F. Forme di resistenza dello zooplancton. In Metodologie di Studio del Plancton Marino, ISPRA—Istituto Superiore per la Protezione e la Ricerca Ambientale; Socal, G., Buttino, I., Cabrini, M., Mangoni, O., Penna, A., Totti, C., Eds.; Institute for Environmental Protection and Research (ISPRA): Rome, Italy, 2010; pp. 507–518. [Google Scholar]
- Mudie, P.J.; Marret, F.; Mertens, K.N.; Shumilovskikh, L.; Leroy, S.A.G. Atlas of modern dinoflagellate cyst distributions in the Black Sea Corridor: From Aegean to Aral Seas, including Marmara, Black, Azov and Caspian Seas. Mar. Micropaleontol. 2017, 134, 1–152. [Google Scholar] [CrossRef]
- Zonneveld, K.A.F.; Pospelova, V. A determination key for modern dinoflagellate cysts. Palynology 2015, 39, 387–409. [Google Scholar] [CrossRef]
- Pielou, E.C. Shannon’s formula as a measure of specific diversity: Its use and misuse. Am. Nat. 1966, 100, 463–465. [Google Scholar] [CrossRef]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication. University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Ellegaard, M.; Godhe, A.; Ribeiro, S. Time capsules in natural sediment archives—Tracking phytoplankton population genetic diversity and adaptation over multidecadal timescales in the face of environmental change. Evol. Appl. 2018, 11, 11–16. [Google Scholar] [CrossRef]
- Pospelova, V.; Chmura, G.L.; Walker, H.A. Environmental factors influencing the spatial distribution of dinoflagellate cyst assemblages in shallow lagoons of southern New England (USA). Rev. Palaeobot. Palynol. 2004, 128, 7–34. [Google Scholar] [CrossRef]
- García-Moreiras, I.; Costas, S.V.; García-Gil, S.; Sobrino, C.M. Organic-walled dinoflagellate cyst assemblages in surface sediments of the Ría de Vigo (Atlantic margin of NW Iberia) in relation to environmental gradients. Mar. Micropaleontol. 2023, 180, 102217. [Google Scholar] [CrossRef]
- Likumahua, S.; Sangiorgi, F.; Boer, M.K.; de Tatipatta, W.M.; Pelasula, D.D.; Polnaya, D.; Hehuwat, J.; Siahaya, D.M.; Buma, A.G.J. Dinoflagellate cyst distribution in surface sediments of Ambon Bay (eastern Indonesia): Environmental conditions and harmful blooms. Mar. Pollut. Bull. 2021, 166, 112269. [Google Scholar] [CrossRef]
- Price, A.M.; Pospelova, V.; Coffin, M.R.S.; Latimer, J.S.; Chmura, G.L. Biogeography of dinoflagellate cysts in northwest Atlantic estuaries. Ecol. Evol. 2016, 6, 5648–5662. [Google Scholar] [CrossRef]
- Rubino, F.; Belmonte, M.; Caroppo, C.; Giacobbe, M. Dinoflagellate cysts from surface sediments of Syracuse Bay (Western Ionian Sea, Mediterranean). Deep Sea Res. Part II Top. Stud. Oceanogr. 2010, 57, 243–247. [Google Scholar] [CrossRef]
- Rubino, F.; Belmonte, M.; Galil, B.S. Plankton resting stages in recent sediments of Haifa port, Israel (Eastern Mediterranean)-Distribution, viability and potential environmental consequences. Mar. Poll. Bull. 2017, 116, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Kremp, A.; Anderson, D.M. Factors regulating germination of resting cysts of the spring bloom dinoflagellate Scrippsiella hangoei from the northern Baltic Sea. J. Plankton Res. 2000, 22, 1311–1327. [Google Scholar] [CrossRef]
- Ribeiro, S.; Berge, T.; Lundholm, N.; Ellegaard, M. Hundred years of environmental change and phytoplankton ecophysiological variability archived in coastal sediments. PLoS ONE 2013, 8, e61184-8. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.Y.; Belmonte, G.; Hyun, B.; Jang, P.-G.; Lee, W.J.; Soh, H.Y.; Seo, M.H.; Seo, J.Y.; Shin, K.; Jang, M.-C. Effects of hypoxia on copepod egg hatching success: An in situ study. J. Mar. Syst. 2023, 245, 103979. [Google Scholar] [CrossRef]
- Rubino, F.; Cibic, T.; Belmonte, M.; Rogelja, M. Microbenthic community structure and trophic status of sediments in the Mar Piccolo of Taranto (Mediterranean, Ionian Sea). Environ. Sci. Poll. Res. 2016, 23, 12624–12644. [Google Scholar] [CrossRef]
- Rodríguez-Villegas, C.; Lee, M.R.; Salgado, P.; Figueroa, R.I.; Baldrich, A.; Pérez-Santos, I.; Tomasetti, S.J.; Niklitschek, E.; Diaz, M.; Alvarez, G.; et al. Drivers of dinoflagellate benthic cyst assemblages in the NW Patagonian fjords system and its adjacent oceanic shelf, with a focus on harmful species. Sci. Total Environ. 2021, 785, 147378. [Google Scholar] [CrossRef]
- Hedberg, P.; Olsson, M.; Höglander, H.; Brückert, V.; Winder, M. Climate change effects on plankton recruitment from coastal sediments. J. Plankton Res. 2024, 46, 117–125. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Station | Lat N | Long E | Depth (m) | Temperature (°C) | Salinity (psu) | Dissolved O2 (%) | pH | PAR (μmol phot m−2 s−1) | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Surface | Bottom | Surface | Bottom | Surface | Bottom | Surface | Bottom | Surface | Bottom | ||||
| G A | 40°26.500′ | 17°08.531′ | 58.4 | 22.1 | 17.3 | 38.2 | 38.9 | 95.2 | 90.4 | 8.36 | 8.31 | 1845.3 | 6.4 |
| G B | 40°25.483′ | 17°08.592′ | 56.2 | 22.3 | 17.6 | 38.3 | 38.9 | 97.5 | 92.3 | 8.36 | 8.32 | 1768.8 | 8.8 |
| G C | 40°25.006′ | 17°10.789′ | 74.5 | 21.9 | 17.2 | 38.2 | 38.8 | 96.5 | 92.5 | 8.35 | 8.32 | 1912.1 | 10.3 |
| MG A | 40°26.191′ | 17°12.212′ | 26.7 | 22.5 | 21.9 | 37.9 | 39.1 | 102.3 | 101.3 | 8.36 | 8.38 | 1913.6 | 7.7 |
| MG B | 40°27.471′ | 17°12.275′ | 18.0 | 22.6 | 22.4 | 38.1 | 39.1 | 89.4 | 80.9 | 8.31 | 8.32 | 1708.6 | 10.2 |
| MG C | 40°26.499′ | 17°13.463′ | 26.8 | 22.7 | 22.2 | 38.2 | 38.8 | 118.0 | 115.3 | 8.40 | 8.36 | 1189.2 | 3.5 |
| MPI A | 40°28.959′ | 17°14.417′ | 10.8 | 22.2 | 22.8 | 36.7 | 38.4 | 97.5 | 91.9 | 8.20 | 8.21 | 2218.6 | 141.2 |
| MPI B | 40°29.481′ | 17°15.303′ | 11.5 | 22.4 | 22.8 | 36.8 | 38.6 | 102.2 | 92.9 | 8.22 | 8.30 | 2551.5 | 134.4 |
| MPI C | 40°28.840′ | 17°15.102′ | 12.1 | 22.4 | 22.8 | 37.3 | 38.6 | 108.6 | 100.7 | 8.24 | 8.30 | 1095.8 | 40.7 |
| MPII A | 40°28.559′ | 17°16.459′ | 12.0 | 22.2 | 22.9 | 36.9 | 38.5 | 98.5 | 75.7 | 8.18 | 8.21 | 2050.3 | 108.2 |
| MPII B | 40°28.845′ | 17°17.681′ | 7.6 | 22.2 | 23.0 | 37.0 | 38.3 | 92.8 | 78.7 | 8.18 | 8.21 | 918.1 | 74.0 |
| MPII C | 40°29.019′ | 17°18.738′ | 5.7 | 22.2 | 23.2 | 36.9 | 38.0 | 107.4 | 86.0 | 8.21 | 8.20 | 1521.8 | 375.9 |
| G | 63.0 ± 10.0 | 22.1 ± 0.2 | 17.4 ± 0.2 | 38.2 ± 0.1 | 38.9 ± 0.1 | 96.4 ± 1.2 | 91.7 ± 1.2 | 8.4 ± 0.0 | 8.3 ± 0.0 | 1842.1 ± 71.7 | 8.5 ± 2.0 | ||
| MG | 23.8 ± 5.1 | 22.6 ± 0.1 | 22.2 ± 0.3 | 38.1 ± 0.2 | 39.0 ± 0.2 | 103.2 ± 14.3 | 99.2 ± 17.3 | 8.4 ± 0.0 | 8.4 ± 0.0 | 1603.8 ± 373.4 | 7.1 ± 3.4 | ||
| MPI | 11.8 ± 0.4 | 22.3 ± 0.1 | 22.8 ± 0.0 | 36.9 ± 0.3 | 38.5 ± 0.1 | 102.8 ± 5.6 | 95.2 ± 4.8 | 8.2 ± 0.0 | 8.3 ± 0.1 | 1955.3 ± 762.7 | 105.4 ± 56.2 | ||
| MPII | 8.4 ± 3.2 | 22.2 ± 0.0 | 23.0 ± 0.2 | 36.9 ± 0.1 | 38.3 ± 0.3 | 99.6 ± 7.4 | 80.1 ± 5.3 | 8.2 ± 0.0 | 8.2 ± 0.0 | 1496.7 ± 566.5 | 186.0 ± 165.3 | ||
| Dinophyta | |
| Paleontological Taxon (Cyst Based) | Modern Taxon |
| # Alexandrium minutum Halim | |
| Alexandrium pseudogonyaulax (Biecheler) Horiguchi | |
| # Alexandrium tamarense (Lebour) Balech | |
| Alexandrium sp. 1 | |
| # Alexandrium sp. 2 | |
| Calciodinellum albatrosianum (Kamptner) | Calciodinellum albatrosianum (Kamptner) Janofske & Karwath |
| Calciodinellum operosum (Deflandre) | Calciodinellum operosum (Deflandre) Montresor |
| Calciperidinium asymmetricum Versteegh | Unknown |
| Cochlodinium polykrikoides Margalef | |
| # Diplopsalis lenticula Bergh | |
| Follisdinellum splendidum Versteegh | Unknown |
| Spiniferites undet | Gonyaulax group |
| # Gymnodinium impudicum (Fraga & Bravo) G. Hansen & Moestrup | |
| Gymnodinium cf. litoralis A. Reñé | |
| * Gymnodinium nolleri Ellegaard & Moestrup | |
| Gymnodinium sp. 1 | |
| Levanderina fissa (Levander) Moestrup, Hakanen, Gert Hansen, Daugbjerg & M.Ellegaard | |
| Lingulodinium machaerophorum (Deflandre & Cookson) Wall | # Lingulodinium polyedra (Stein) Dodge |
| Lingulodinium sp. | |
| Melodomuncula berlinensis Versteegh | Unknown |
| Oblea acanthocysta Kawami, Iwataki & Matsuoka | |
| # Oblea rotunda (Lebour) Balech ex Sournia | |
| # Pentapharsodinium dalei Indelicato & Loeblich | |
| Calcicarpinum bivalvum Versteegh | # 1 Pentapharsodinium tyrrhenicum Montresor, Zingone & Marino |
| Calcicarpinum bivalvum Versteegh | Pentapharsodinium cf. tyrrhenicum Montresor, Zingone & Marino |
| Polykrikos kofoidii Chatton | |
| Polykrikos schwartzii Bütschli | |
| Bicarinellum tricarinelloides Versteegh | Posoniella tricarinelloides (Versteegh) Streng et al. |
| Operculodinium centrocarpum (Deflandre & Cookson) Wall | Protoceratium reticulatum (Claparède & Lachmann) Butschli |
| Protoperidinium claudicans (Paulsen) Balech | |
| Selenopemphix quanta (Bradford) Matsuoka | Protoperidinium conicum (Gran) Balech |
| Xandarodinium xanthum Reid | # Protoperidinium divaricatum (Meunier) Parke & Dodge |
| Votadinium calvum Reid | Protoperidinium latidorsale (Balech) Balech |
| Quinquecuspis concretum (Reid) Harland | Protoperidinium leonis (Pavillard) Balech |
| Protoperidinium minutum (Kofoid) Loeblich III | |
| Protoperidinium parthenopes Zingone & Montresor | |
| Trinovantedinium applanatum (Bradford) Bujak & Davies | Protoperidinium shanghaiense Gu, Liu & Mertens |
| # Protoperidinium stellatum (Wall in Wall & Dale 1968) Head | |
| Protoperidinium thorianum (Paulsen) Balech | |
| Protoperidinium sp. 1 | |
| Protoperidinium sp. 4 | |
| Protoperidinium sp. 5 | |
| cf. Pseudocochlodinium profundisulcus Hu, Xu, Gu, Iwataki, Takahashi, Tang & Matsuoka | |
| Polysphaeridium zoharyi (Rosignol) Bujak et al. | Pyrodinium bahamense var. compressum (Böhm) Steidinger, Tester & Taylor |
| Pyrophacus horologium Stein | |
| 2 Scrippsiella acuminata complex (Ehrenb.) Kretschmann, Elbr., Zinssmeister, S. Soehner, Kirsch, Kusber & Gottschling | |
| Scrippsiella cf. erinaceus (Kamptner) Kretschmann, Zinssmeister & Gottschling | |
| # Scrippsiella lachrymosa Lewis | |
| Scrippsiella precaria Montresor & Zingone | |
| # Scrippsiella ramonii Montresor | |
| # Scrippsiella spinifera Honsell & Cabrini | |
| Scrippsiella trifida Lewis | |
| # Scrippsiella sp. 1 | |
| # Scrippsiella sp. 3 | |
| # Scrippsiella sp. 4 | |
| Scrippsiella sp. 5 | |
| Scrippsiella sp. 6 | |
| Scrippsiella sp. 8 | |
| * Warnowia rosea (Pouchet) Kofoid & Schwezy | |
| Dinophyta sp. 1 | |
| Dinophyta sp. 2 | |
| Dinophyta sp. 9 | |
| Dinophyta sp. 14 | |
| Dinophyta sp. 17 | |
| * Dinophyta sp. 24 | |
| * Dinophyta sp. 26 | |
| Dinophyta sp. 29 | |
| Dinophyta sp. 31 | |
| Ciliophora | Crustacea Copepoda |
| Cyrtostrombidium boreale Kim, Suzuki & Taniguchi | Acartia cf. italica Steuer (subitaneous egg) |
| Hexasterias problematica Cleve | Acartia sp. 2 |
| *Laboea strobila Lohman | Acartia sp. 3 |
| Strombidium acutum (Claparéde & Lachmann) Jörgensen | Acartia sp. 4 |
| Strombidium conicum (Lohmann) Wulff | Centropages sp. (resting egg) |
| Strombidium crassulum (Leegaard) Kahl | * Centropages sp. (subitaneous egg) |
| Strombidium sp. | Paracartia latisetosa (Krizcaguin) |
| Ciliophora sp. 1 | Pteriacartia josephinae (Crisafi) |
| Ciliophora sp. 2 | Copepoda sp. 1 |
| Ciliophora sp. 5 | |
| Ciliophora sp. 6 | |
| Ciliophora sp. 7 | |
| Rotifera | Unidentified |
| Synchaeta sp. 1 (smooth) | Cyst type 2 |
| Synchaeta sp. 2 (tubular) | Cyst type 3 |
| Synchaeta sp. 3 (spiny) | * Cyst type 4 |
| Resting egg 1 |
| Full Cysts | Empty Cysts | |||||||
|---|---|---|---|---|---|---|---|---|
| Station | S | N | J′ | H′(loge) | S | N | J′ | H′(loge) |
| G A | 31 | 708 ± 437 | 0.69 | 2.38 | 43 | 2491 ± 790 | 0.60 | 2.27 |
| G B | 33 | 430 ± 163 | 0.71 | 2.47 | 48 | 1688 ± 321 | 0.59 | 2.30 |
| G C | 29 | 467 ± 283 | 0.68 | 2.29 | 48 | 2113 ± 579 | 0.57 | 2.20 |
| MG A | 46 | 1774 ± 1189 | 0.68 | 2.60 | 54 | 4292 ± 618 | 0.65 | 2.59 |
| MG B | 47 | 1687 ± 722 | 0.73 | 2.82 | 59 | 4067 ± 808 | 0.66 | 2.71 |
| MG C | 51 | 2382 ± 1533 | 0.72 | 2.84 | 59 | 7115 ± 2187 | 0.62 | 2.52 |
| MPI A | 45 | 1726 ± 414 | 0.76 | 2.90 | 57 | 4135 ± 849 | 0.70 | 2.84 |
| MPI B | 56 | 4451 ± 3320 | 0.74 | 2.97 | 59 | 9443 ± 2066 | 0.71 | 2.88 |
| MPI C | 44 | 4147 ± 1554 | 0.75 | 2.83 | 53 | 9785 ± 836 | 0.73 | 2.89 |
| MPII A | 53 | 2860 ± 2238 | 0.74 | 2.93 | 55 | 7941 ± 2881 | 0.70 | 2.81 |
| MPII B | 50 | 2711 ± 687 | 0.72 | 2.80 | 56 | 6360 ± 788 | 0.65 | 2.63 |
| MPII C | 46 | 1786 ± 681 | 0.74 | 2.83 | 37 | 1833 ± 1350 | 0.56 | 2.03 |
| basin | ||||||||
| G | 44 | 535 ± 303 | 0.69 | 2.38 | 59 | 2097 ± 622 | 0.59 | 2.26 |
| MG | 66 | 1948 ± 1086 | 0.71 | 2.75 | 71 | 5158 ± 1902 | 0.64 | 2.61 |
| MPI | 68 | 3441 ± 2253 | 0.75 | 2.90 | 71 | 7787 ± 2992 | 0.71 | 2.87 |
| MPII | 71 | 2452 ± 1319 | 0.73 | 2.85 | 65 | 5378 ± 3182 | 0.64 | 2.49 |
| Groups | R | p Value |
|---|---|---|
| G, MG | 0.448 | 0.001 |
| G, MPI | 0.754 | 0.001 |
| G, MPII | 0.869 | 0.001 |
| MG, MPI | 0.245 | 0.014 |
| MG, MPII | 0.580 | 0.001 |
| MPI, MPII | 0.106 | 0.094 |
| Group | Aver. Simil. | Groups | Aver.Dissimil. |
|---|---|---|---|
| G | 58.80 | G, MG | 43.67 |
| MG | 65.99 | G, MPI | 47.65 |
| MPI | 65.87 | G, MPII | 49.84 |
| MPII | 66.41 | MG, MPI | 36.69 |
| MG, MPII | 39.98 | ||
| MPI, MPII | 34.68 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubino, F.; Denti, G.; Belmonte, M.; Belmonte, G. Plankton Resting Stages Distribution in Bottom Sediments along the Confinement Gradient of the Taranto Sea System (Ionian Sea, Southern Italy). J. Mar. Sci. Eng. 2024, 12, 1653. https://doi.org/10.3390/jmse12091653
Rubino F, Denti G, Belmonte M, Belmonte G. Plankton Resting Stages Distribution in Bottom Sediments along the Confinement Gradient of the Taranto Sea System (Ionian Sea, Southern Italy). Journal of Marine Science and Engineering. 2024; 12(9):1653. https://doi.org/10.3390/jmse12091653
Chicago/Turabian StyleRubino, Fernando, Giuseppe Denti, Manuela Belmonte, and Genuario Belmonte. 2024. "Plankton Resting Stages Distribution in Bottom Sediments along the Confinement Gradient of the Taranto Sea System (Ionian Sea, Southern Italy)" Journal of Marine Science and Engineering 12, no. 9: 1653. https://doi.org/10.3390/jmse12091653
APA StyleRubino, F., Denti, G., Belmonte, M., & Belmonte, G. (2024). Plankton Resting Stages Distribution in Bottom Sediments along the Confinement Gradient of the Taranto Sea System (Ionian Sea, Southern Italy). Journal of Marine Science and Engineering, 12(9), 1653. https://doi.org/10.3390/jmse12091653