
Selenium Nanomaterials Enhance the Nutrients and Functional Components of Fuding Dabai Tea
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Synthesis and Characteristic of Se NMs
2.2. Field Experimental Design and NMs Exposure
2.3. Mineral Elements Analysis after Se NMs in Tea Buds
2.4. Theanine, Catechins, and Caffeine Analysis and Metabolic Response of Tea Buds
2.5. Tea Soil Microbial Community Structure Analysis Based on 16S rRNA Gene Sequencing
2.6. Related Gene Expression of Quantitative Real-Time (qPCR) Analysis
2.7. Statistical Analysis
3. Results and Discussion
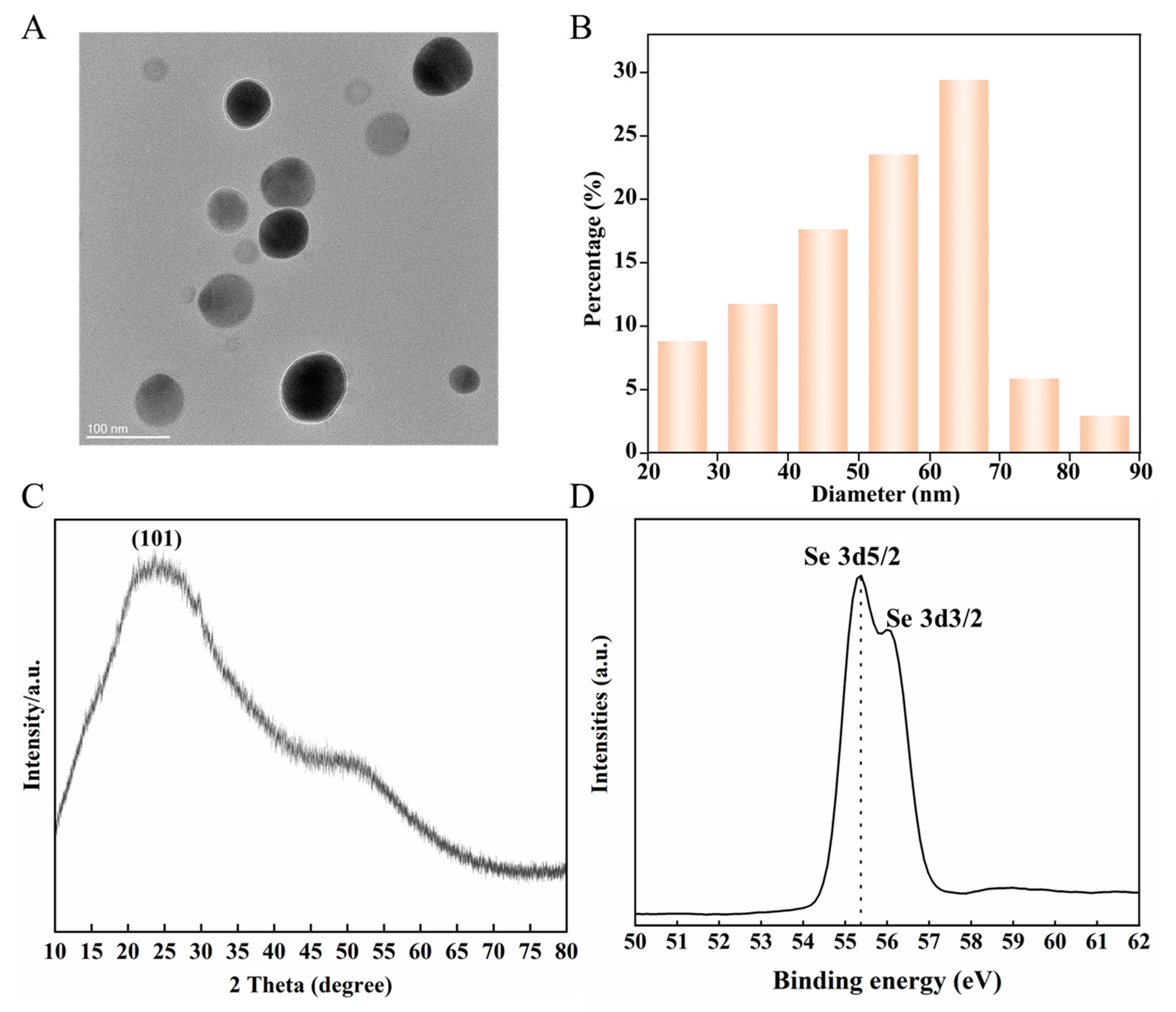
3.1. Characterization of Se NMs
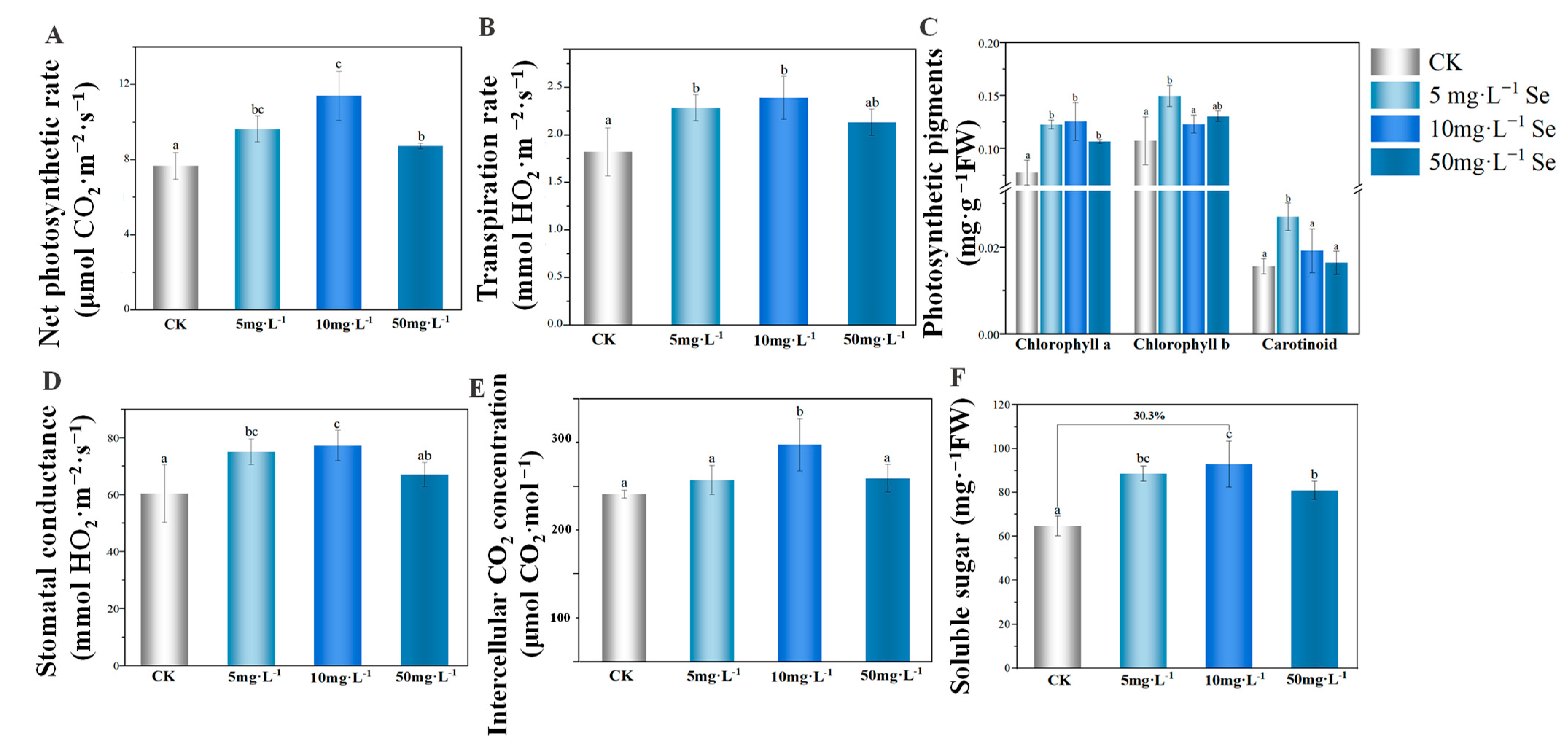
3.2. The Growth of Tea Plant after the Foliar Application of Se NMs
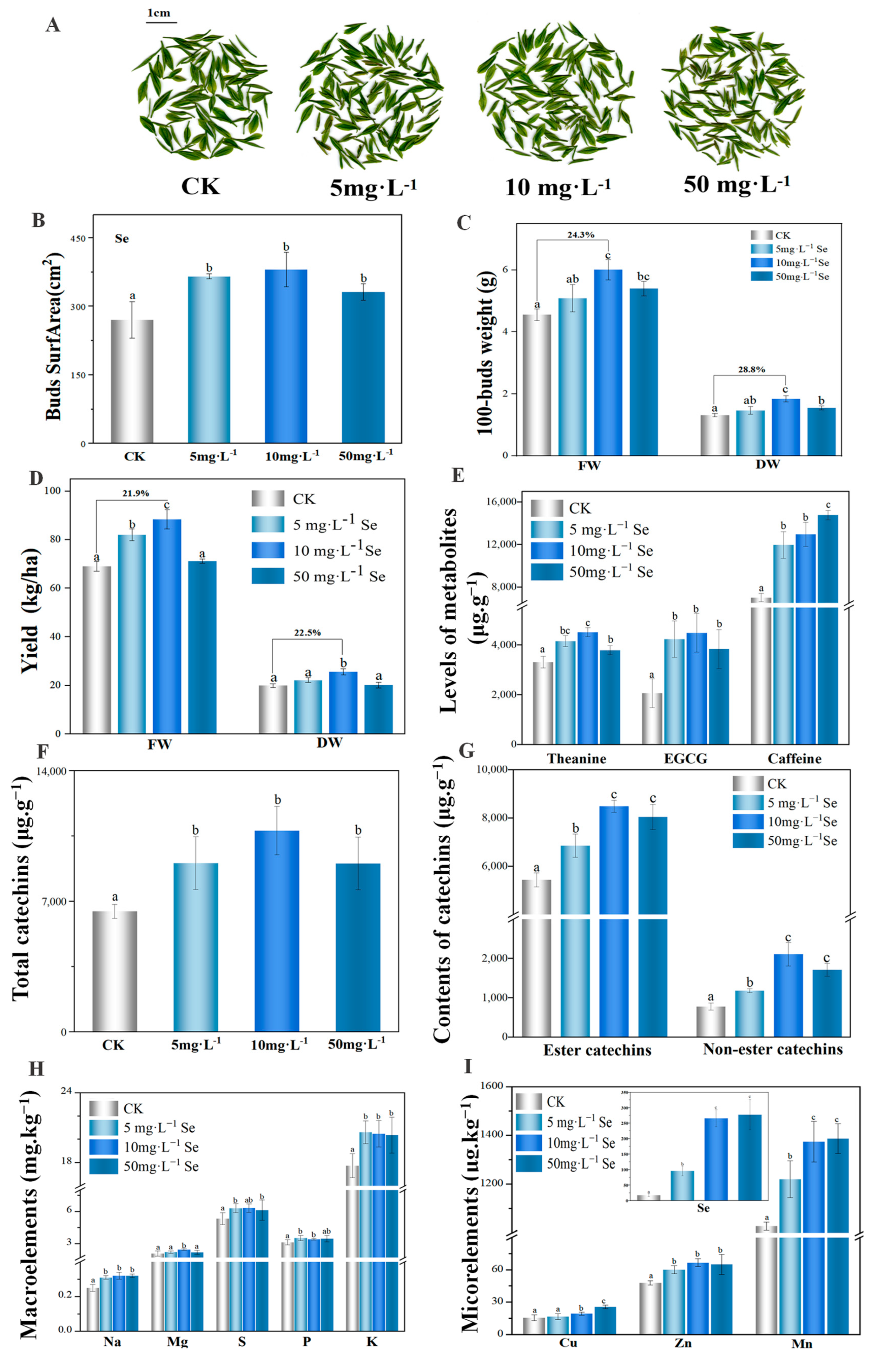
3.3. The Yield and Quality of Tea Plant after the Foliar Application of Se NMs
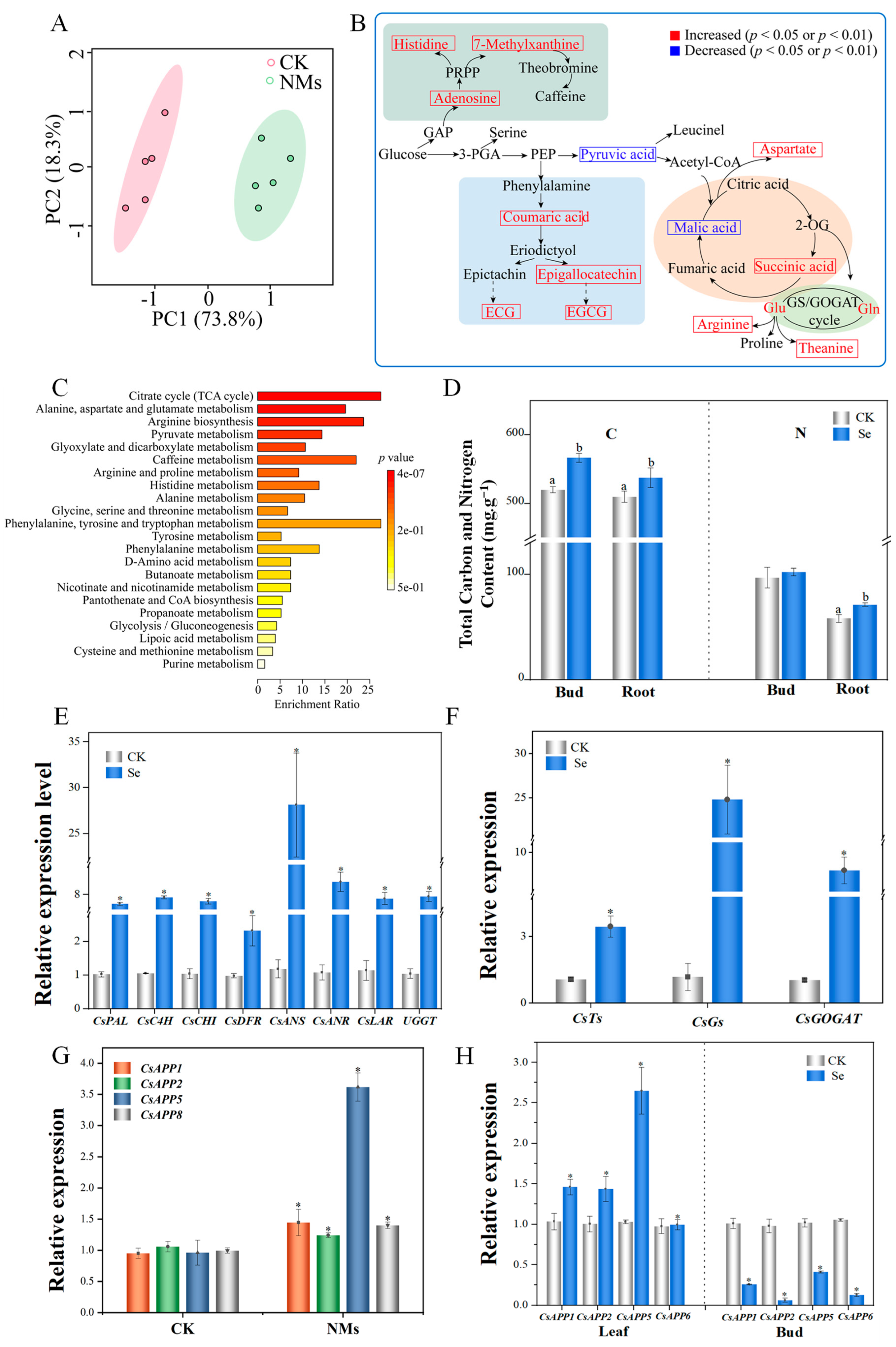
3.4. Foliar Applied Se NMs Improve the Synthesis and Transport of Catechins and Theanine in Tea Buds
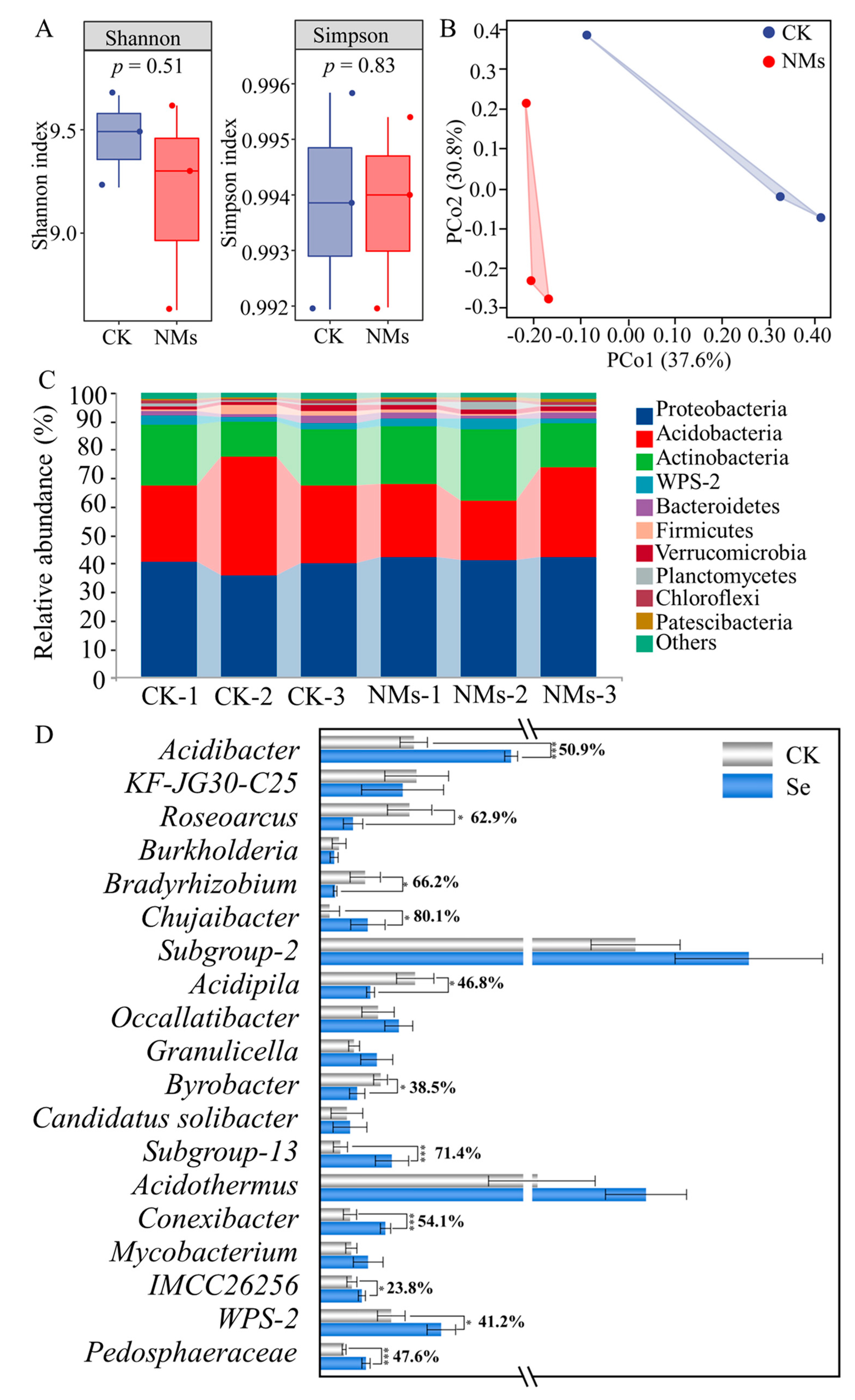
3.5. Application of NMs Changes the Diversity and Structure of Soil Microbial Communities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vuong, Q.V.; Bowyer, M.C.; Roach, P.D. l-Theanine: Properties, synthesis and isolation from tea. J. Sci. Food Agric. 2011, 91, 1931–1939. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Liu, Z.; Huang, J.; Luo, G.; Liang, Q.; Wang, D.; Ye, X.; Wu, C.; Wang, L.; Hu, J. Anti-obesity and hypolipidemic effects of Fuzhuan brick tea water extract in high-fat diet-induced obese rats. J. Sci. Food Agric. 2013, 93, 1310–1316. [Google Scholar] [CrossRef] [PubMed]
- Arnoriaga-Rodriguez, M.; Mayneris-Perxachs, J.; Burokas, A.; Contreras-Rodriguez, O.; Blasco, G.; Coll, C.; Biarnes, C.; Miranda-Olivos, R.; Latorre, J.; Moreno-Navarrete, J.M.; et al. Obesity impairs short-term and working memory through gut microbial metabolism of aromatic amino acids. Cell Metab. 2020, 32, 548–560. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Mondal, A.; Majumder, A.; Banik, A. Tea and its phytochemicals: Hidden health benefits & modulation of signaling cascade by phytochemicals. Food Chem. 2022, 371, 131098. [Google Scholar] [PubMed]
- Anderson, M.R.; Shashaty, M.G.S. Impact of obesity in critical illness. Chest 2021, 160, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.K.; Lin-Shiau, S.Y. Mechanisms of hypolipidemic and anti-obesity effects of tea and tea polyphenols. Mol. Nutr. Food Res. 2006, 50, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zhong, Y.; Duan, Y.; Chen, Q.; Li, F. Antioxidant mechanism of tea polyphenols and its impact on health benefits. Anim. Nutr. 2020, 6, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Feng, H.; Yang, F.; Zhao, Z.; Hu, X.; Wei, C.; Liang, T.; Li, H.; Geng, Y. Does dual reduction in chemical fertilizer and pesticides improve nutrient loss and tea yield and quality? A pilot study in a green tea garden in Shaoxing, Zhejiang Province, China. Environ. Sci. Pollut. Res. Int. 2019, 26, 2464–2476. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.-d.; Ni, K.; Shi, Y.-z.; Yi, X.-y.; Zhang, Q.-f.; Fang, L.; Ma, L.-f.; Ruan, J. Effects of long-term nitrogen application on soil acidification and solution chemistry of a tea plantation in China. Agric. Ecosyst. Environ. 2018, 252, 74–82. [Google Scholar] [CrossRef]
- Ruan, J.; Ma, L.; Shi, Y. Potassium management in tea plantations: Its uptake by field plants, status in soils, and efficacy on yields and quality of teas in China. J. Plant Nutr. Soil Sci. 2013, 176, 450–459. [Google Scholar] [CrossRef]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in sustainable agriculture: Recent developments, challenges, and perspectives. Front. Microbiol. 2017, 8, 1014. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Cai, L.; Jia, H.; Liu, C.; Wang, D.; Sun, X. Foliar exposure of Fe3O4 nanoparticles on Nicotiana benthamiana: Evidence for nanoparticles uptake, plant growth promoter and defense response elicitor against plant virus. J. Hazard. Mater. 2020, 393, 122415. [Google Scholar] [CrossRef]
- Li, J.; Ma, Y.; Xie, Y. Stimulatory effect of Fe3O4 Nanoparticles on the growth and yield of Pseudostellaria heterophylla via improved photosynthetic performance. HortScience 2021, 56, 753–761. [Google Scholar] [CrossRef]
- Ghafariyan, M.H.; Malakouti, M.J.; Dadpour, M.R.; Stroeve, P.; Mahmoudi, M. Effects of magnetite nanoparticles on soybean chlorophyll. Environ. Sci. Technol. 2013, 47, 10645–10652. [Google Scholar] [CrossRef]
- Mikula, K.; Izydorczyk, G.; Skrzypczak, D.; Mironiuk, M.; Moustakas, K.; Witek-Krowiak, A.; Chojnacka, K. Controlled release micronutrient fertilizers for precision agriculture—A review. Sci. Total. Environ. 2020, 712, 136365. [Google Scholar] [CrossRef] [PubMed]
- Serov, D.A.; Khabatova, V.V.; Vodeneev, V.; Li, R.; Gudkov, S.V. A review of the antibacterial, fungicidal and antiviral properties of selenium nanoparticles. Materials 2023, 16, 5363. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Gao, M.; Simakin, A.V.; Baryshev, A.S.; Pobedonostsev, R.V.; Baimler, I.V.; Rebezov, M.B.; Sarimov, R.M.; Astashev, M.E.; Dikovskaya, A.O.; et al. Laser ablation-generated crystalline selenium nanoparticles prevent damage of DNA and proteins induced by reactive oxygen species and protect mice against injuries caused by radiation-induced oxidative stress. Materials 2023, 16, 5164. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhou, C.; Zou, N.; Wu, Y.; Zhang, J.; An, Q.; Li, J.Q.; Pan, C. Nanoselenium foliar application enhances biosynthesis of tea leaves in metabolic cycles and associated responsive pathways. Environ. Pollut. 2021, 273, 116503. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Liu, K.; Li, S.; Han, J.; Zeng, X.; Ling, M.; Mao, J.; Li, Y.; Jiang, J. Effect of selenium on tea (Camellia sinensis) under low temperature: Changes in physiological and biochemical responses and quality. Environ. Exp. Bot. 2021, 188, 104475. [Google Scholar] [CrossRef]
- Liu, S.H.; Li, J.; Huang, J.A.; Liu, Z.H.; Xiong, L.G. New advances in genetic engineering for l-theanine biosynthesis. Trends Food Sci. Technol. 2021, 114, 540–551. [Google Scholar] [CrossRef]
- Galili, G.; Amir, R.; Fernie, A.R. The Regulation of Essential Amino Acid Synthesis and Accumulation in Plants. Annu. Rev. Plant Biol. 2016, 67, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Sharma, A.R.; Bhavesh, R.; Park, J.; Ganbold, B.; Nam, J.S.; Lee, S.S. Biomolecule-mediated synthesis of selenium nanoparticles using dried Vitis vinifera (raisin) extract. Molecules 2014, 19, 2761–2770. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Xu, X.; Xu, C.; Wang, H.; Zhang, X.; Liu, G. Preparation, characterization, and phosphate removal and recovery of magnetic MnFe2O4 nano-particles as adsorbents. Environ. Technol. 2016, 37, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zuo, T.; Xu, W.; Zhang, Y.; Ni, W. Improving albino tea quality by foliar application of glycinebetaine as a green regulator under lower temperature conditions. J. Agric. Food Chem. 2021, 69, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Arcos, C.; Kai, M.; Svatos, A.; Gershenzon, J.; Kunert, G. Untargeted metabolomics approach reveals differences in host plant chemistry before and after infestation with different pea aphid host races. Front. Plant Sci. 2019, 10, 00188. [Google Scholar] [CrossRef] [PubMed]
- Wa’el, A.A.; Nelson, A. Characterization of MoSe2(0001) and ion-sputtered MoSe2 by XPS. J. Mater. 2005, 40, 2679–2681. [Google Scholar]
- Avellan, A.; Yun, J.; Zhang, Y.; Spielman-Sun, E.; Unrine, J.M.; Thieme, J.; Li, J.; Lombi, E.; Bland, G.; Lowry, G.V. Nanoparticle size and coating chemistry control foliar uptake pathways, translocation, and leaf-to-rhizosphere transport in wheat. ACS Nano 2019, 13, 5291–5305. [Google Scholar] [CrossRef] [PubMed]
- Avellan, A.; Yun, J.; Morais, B.P.; Clement, E.T.; Rodrigues, S.M.; Lowry, G.V. Critical Review: Role of inorganic nanoparticle properties on their foliar uptake and in planta translocation. Environ. Sci. Technol. 2021, 55, 13417–13431. [Google Scholar] [CrossRef]
- Nair, R.; Varghese, S.H.; Nair, B.G.; Maekawa, T.; Yoshida, Y.; Kumar, D.S. Nanoparticulate material delivery to plants. Plant Sci. 2010, 179, 154–163. [Google Scholar] [CrossRef]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Perez-Alvarez, M.; Hernandez-Fuentes, A.D.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Valdes-Reyna, J.; Juarez-Maldonado, A. Se nanoparticles induce changes in the growth, antioxidant responses, and fruit quality of tomato developed under NaCl stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Zhao, C.; Zhang, T. Selenium transformation and selenium-rich foods. Food Biosci. 2021, 40, 100875. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Tsuji, P.A.; Carlson, B.A.; Gladyshev, V.N. Selenium and selenocysteine: Roles in cancer, health, and development. Trends Biochem. Sci. 2013, 39, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.-Y.; Tang, D.; Shi, Y.; Ma, L.; Li, Y.; Zhang, Q.; Ruan, J. Short-term inhibition of glutamine synthetase leads to reprogramming of amino acid and lipid metabolism in roots and leaves of tea plant (Camellia sinensis L.). BMC Plant Biol. 2019, 19, 425. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Liu, Y.; Zhao, L.; Jiang, X.; Li, M.; Wang, Y.; Xu, Y.; Gao, L.; Xia, T. Effect of low-intensity white light mediated de-etiolation on the biosynthesis of polyphenols in tea seedlings. Plant Physiol. Biochem. 2014, 80, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Li, F.; Yang, T.; Feng, L.; Zhang, S.; Li, F.; Li, W.; Xu, G.; Bao, S.; Wan, X.; et al. Theanine transporters identified in tea plants (Camellia sinensis L.). Plant J. 2020, 101, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Zhang, J.; Yu, Y.; Tian, Y.; Li, H.; Chen, X.; Li, W.; Liu, Y.; Lu, T.; He, B. Root microbiota of tea plants regulate nitrogen homeostasis and theanine synthesis to influence tea quality. Curr. Biol. 2024, 34, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yue, L.; Dhankher, O.P.; Xing, B. Nano-enabled improvements of growth and nutritional quality in food plants driven by rhizosphere processes. Environ. Int. 2020, 142, 105831. [Google Scholar] [CrossRef]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loque, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef]
- Yao, L.; Ding, C.; Hao, X.; Zeng, J.; Yang, Y.; Wang, X.; Wang, L. CsSWEET1a and CsSWEET17 mediate growth and freezing tolerance by promoting sugar transport across the plasma membrane. Plant Cell Physiol. 2020, 61, 1669–1682. [Google Scholar] [CrossRef] [PubMed]
- Smeekens, S.; Ma, J.; Hanson, J.; Rolland, F. Sugar signals and molecular networks controlling plant growth. Curr. Opin. Plant Biol. 2010, 13, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cao, X.; Wang, C.; Chen, F.; Feng, Y.; Yue, L.; Wang, Z.; Xing, B. Fe-based nanomaterial-induced root nodulation is modulated by flavonoids to improve soybean (Glycine max) growth and quality. ACS Nano 2022, 16, 21047–21062. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Jiao, L.; Tao, M.; Xu, L.; Cao, X.; Chen, F.; Wang, C.; Cheng, B.; Wang, Z. Dynamics of organic acid exudation and rhizobacteria in maize rhizosphere respond to N-CDs. Sci. Total Environ. 2023, 901, 166500. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2011, 32, 227–243. [Google Scholar] [CrossRef]
- Gulati, A.; Sood, S.; Rahi, P.; Thakur, R.; Chauhan, S.; Chadha, I.C.N. Diversity analysis of diazotrophic bacteria associated with the roots of Tea (Camellia sinensis (L.) O. Kuntze). J. Microbiol. Biotechnol. 2011, 21, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Li, H.; Yin, P.; Mei, H.; Li, J.; Zhou, P.; Wang, Y.; Ma, Q.; Jeyaraj, A.; Thangaraj, K.; et al. Integrated application of rapeseed cake and green manure enhances soil nutrients and microbial communities in tea garden soil. Sustainability 2021, 13, 2967. [Google Scholar] [CrossRef]
- Chen, W.-C.; Ko, C.-H.; Su, Y.-S.; Lai, W.-A.; Shen, F.-T. Metabolic potential and community structure of bacteria in an organic tea plantation. Appl. Soil Ecol. 2021, 157, 103762. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Lee, S.D. Conexibacter stalactiti sp. nov., isolated from stalactites in a lava cave and emended description of the genus Conexibacter. Int. J. Syst. Evol. Microbiol. 2017, 67, 3214–3218. [Google Scholar] [CrossRef]
- Yu, F.; Liang, J.F.; Song, J.; Wang, S.K.; Lu, J.K. Bacterial community selection of Russula griseocarnosa mycosphere soil. Front. Microbiol. 2020, 11, 00347. [Google Scholar] [CrossRef] [PubMed]
- Ruan, L.; Wei, K.; Wang, L.; Cheng, H.; Zhang, F.; Wu, L.; Bai, P.; Zhang, C. Characteristics of NH4+ and NO3− fluxes in tea (Camellia sinensis) roots measured by scanning ion-selective electrode technique. Sci. Rep. 2016, 6, 38370. [Google Scholar] [CrossRef] [PubMed]
- Anthony, D.; Dev, T.; Brent, N.; James, R.; Herbert, J.; Anshuman, K.; Mamoru, O.; Suman, R.; Siddiqi, M.; Shiela, E.; et al. The regulation of nitrate and ammonium transport systems in plants. J. Exp. Bot. 2002, 53, 855–864. [Google Scholar]
- Fan, K.; Zhang, Q.; Tang, D.; Shi, Y.; Ma, L.; Liu, M.; Ruan, J. Dynamics of nitrogen translocation from mature leaves to new shoots and related gene expression during spring shoots development in tea plants (Camellia sinensis L.). J. Plant Nutr. Soil Sci. 2020, 183, 180–191. [Google Scholar] [CrossRef]
- Dechorgnat, J.; Nguyen, C.T.; Armengaud, P.; Jossier, M.; Diatloff, E.; Filleur, S.; Daniel-Vedele, F. From the soil to the seeds: The long journey of nitrate in plants. J. Exp. Bot. 2011, 62, 1349–1359. [Google Scholar] [CrossRef]
- Tang, S.; Ma, Q.; Luo, J.; Xie, Y.; Hashmi, M.; Pan, W.; Zheng, N.; Liu, M.; Wu, L. The inhibition effect of tea polyphenols on soil nitrification is greater than denitrification in tea garden soil. Sci. Total Environ. 2021, 778, 146328. [Google Scholar] [CrossRef]
- Zhan, X.; Yuan, J.; Yue, L.; Xu, G.; Hu, B.; Xu, R. Response of uptake and translocation of phenanthrene to nitrogen form in lettuce and wheat seedlings. Environ. Sci. Pollut. Res. Int. 2015, 22, 6280–62873. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Li, X.; Chen, F.; Cao, X.; Wang, C.; Jiao, L.; Yue, L.; Wang, Z. Selenium Nanomaterials Enhance the Nutrients and Functional Components of Fuding Dabai Tea. Nanomaterials 2024, 14, 681. https://doi.org/10.3390/nano14080681
Zhang X, Li X, Chen F, Cao X, Wang C, Jiao L, Yue L, Wang Z. Selenium Nanomaterials Enhance the Nutrients and Functional Components of Fuding Dabai Tea. Nanomaterials. 2024; 14(8):681. https://doi.org/10.3390/nano14080681
Chicago/Turabian StyleZhang, Xiaoli, Xiaona Li, Feiran Chen, Xuesong Cao, Chuanxi Wang, Liya Jiao, Le Yue, and Zhenyu Wang. 2024. "Selenium Nanomaterials Enhance the Nutrients and Functional Components of Fuding Dabai Tea" Nanomaterials 14, no. 8: 681. https://doi.org/10.3390/nano14080681